
Di- and Mono-Rhamnolipids Produced by the Pseudomonas putida PP021 Isolate Significantly Enhance the Degree of Recovery of Heavy Oil from the Romashkino Oil Field (Tatarstan, Russia)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
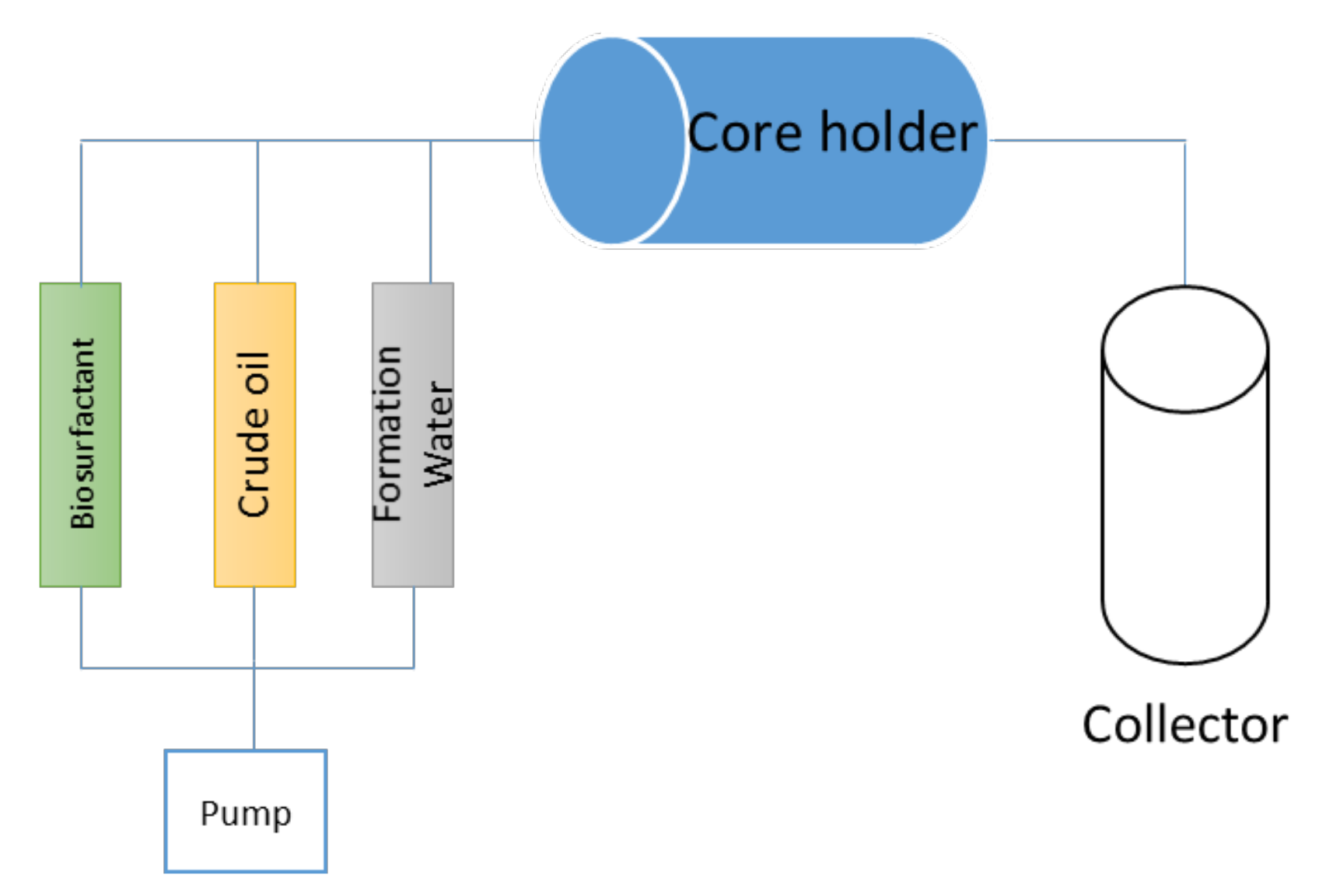
2. Materials and Methods
3. Results
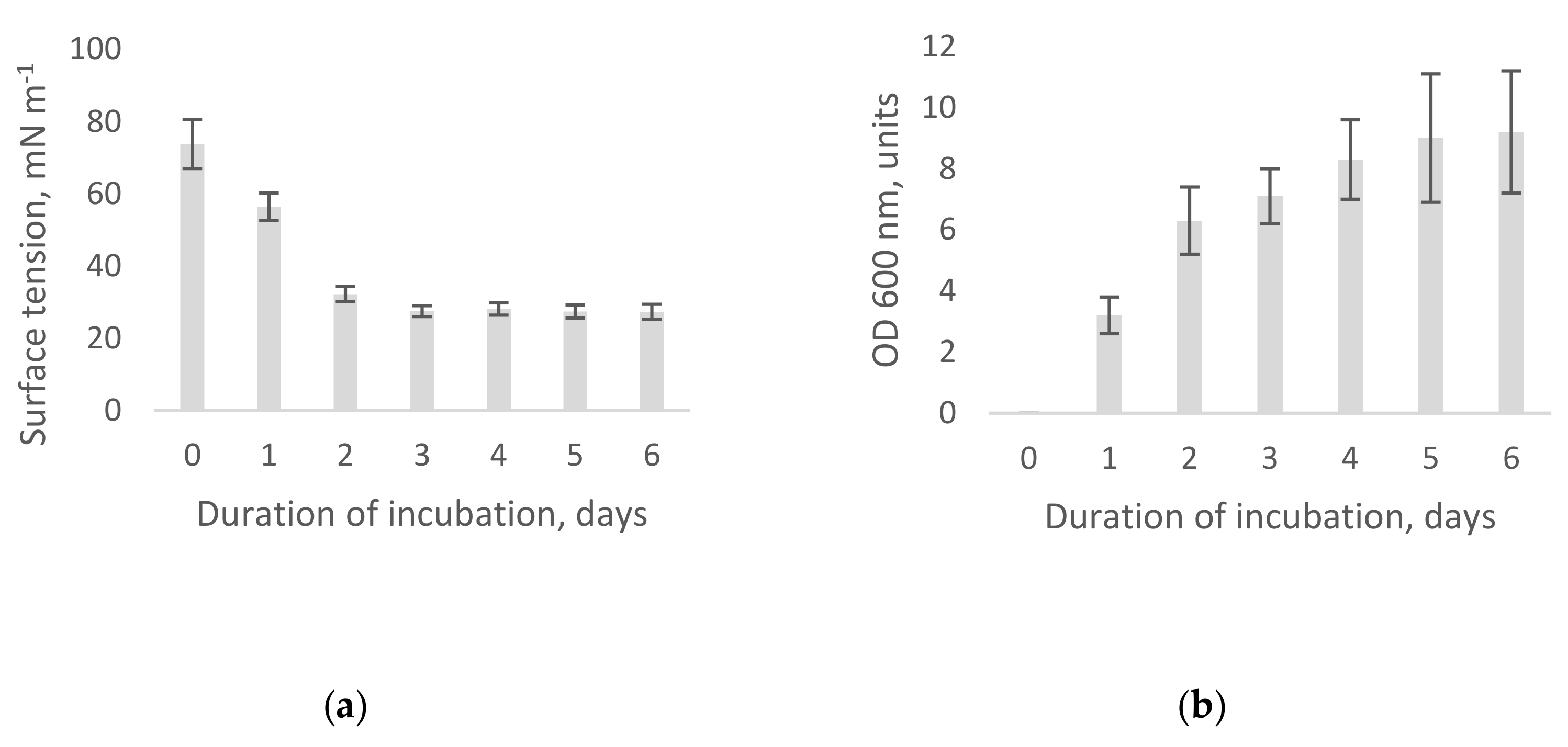
3.1. Air–Water Interface Tension
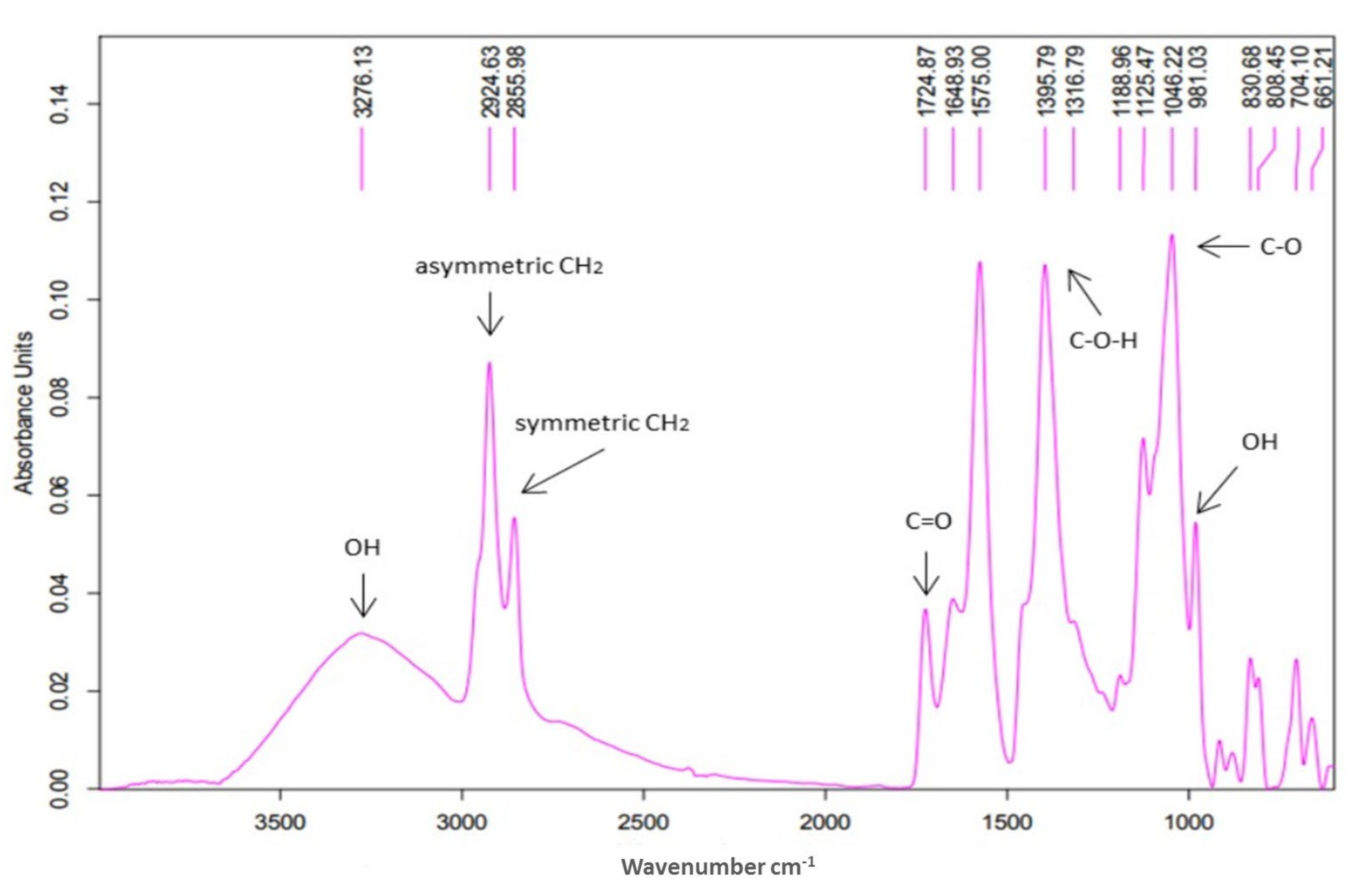
3.2. TLC and FTIR
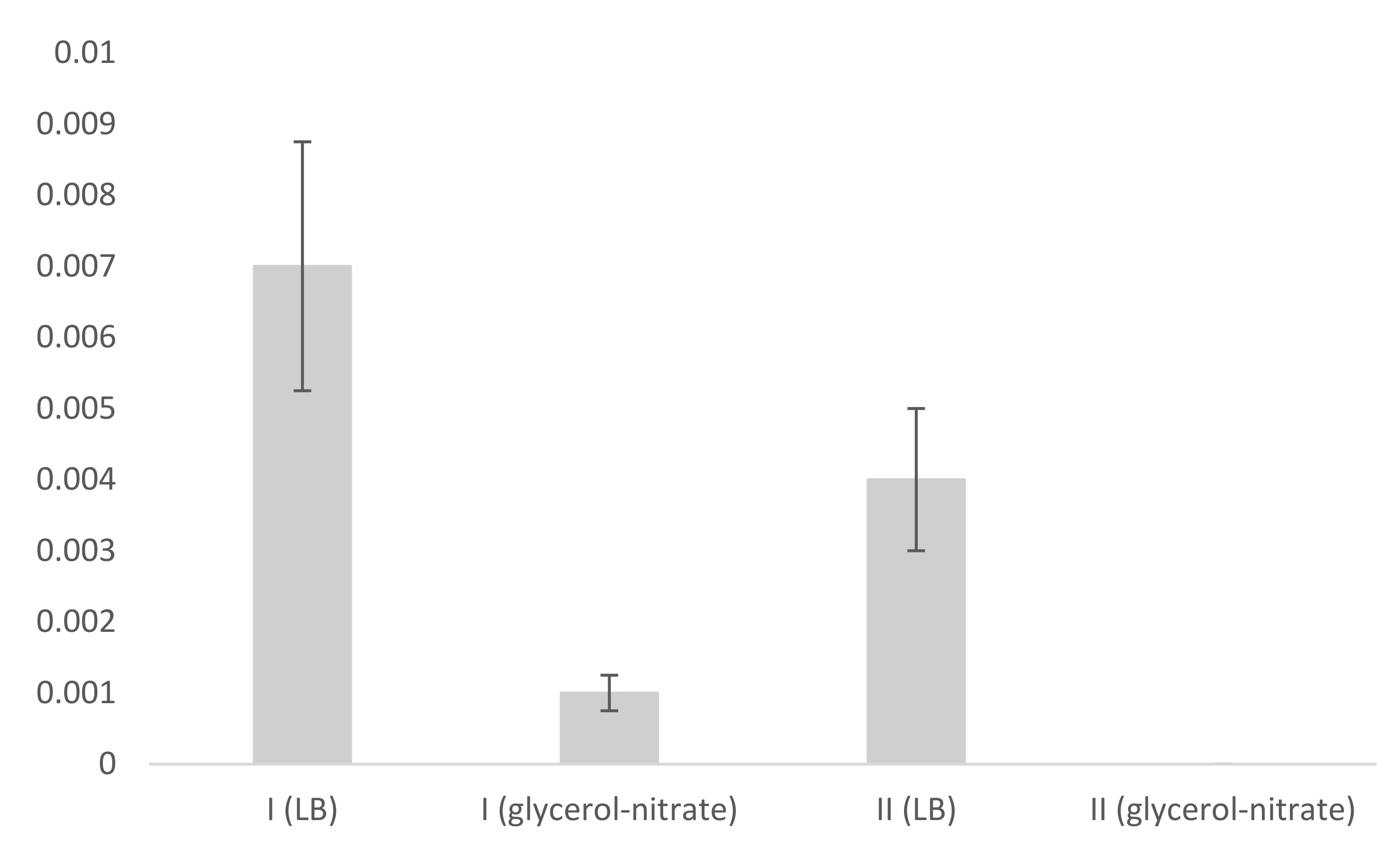
3.3. Presence and Relative Expression of Genes Encoding Rhamnotransferase Synthesis
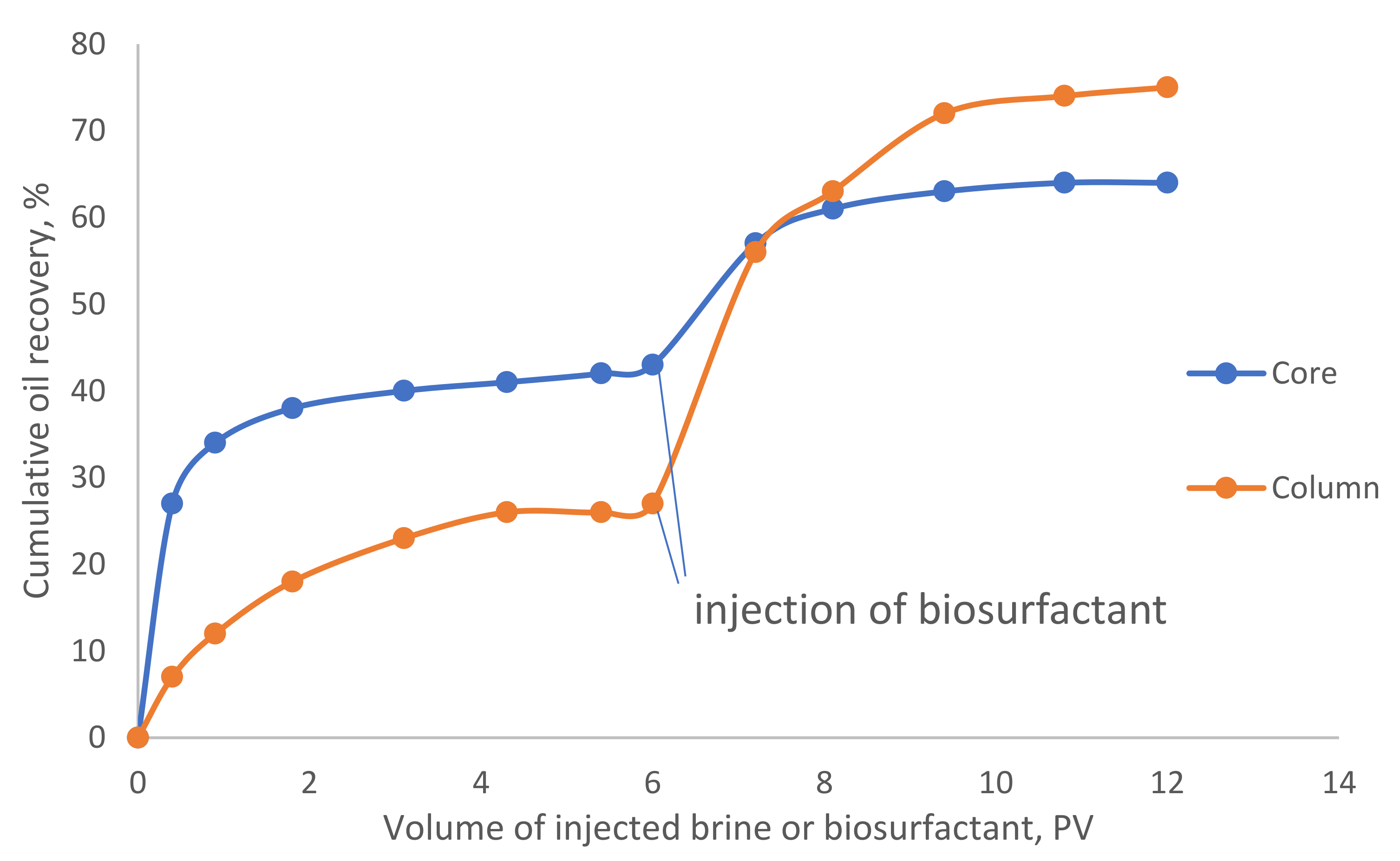
3.4. Enhanced Oil Recovery
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shibulal, B.; Al-Bahry, S.N.; Al-Wahaibi, Y.M.; Elshafie, A.E.; Al-Bemani, A.S.; Joshi, S.J. Microbial enhanced heavy oil recovery by the aid of inhabitant spore-forming bacteria: An insight review. Sci. World J. 2014, 2014, 309159. [Google Scholar] [CrossRef] [PubMed]
- Alikhlalov, K.; Dindoruk, B. Conversion of cyclic steam injection to continous steam injection. In Proceedings of the Proceedings-SPE Annual Technical Conference and Exhibition, Denver, CO, USA, 30 October–2 November 2011. [Google Scholar]
- Lu, L.; Liu, Z.; Liu, H.; Yan, Y. Study on Technical Measures of Romashkino Oil Field after Entering Ultra-High Water Cut Stage. Engineering 2013, 5, 622–628. [Google Scholar] [CrossRef] [Green Version]
- Galitskaya, P.; Gumerova, R.; Ratering, S.; Schnell, S.; Blagodatskaya, E.; Selivanovskaya, S. Oily waste containing natural radionuclides: Does it cause stimulation or inhibition of soil bacterial community? J. Plant Nutr. Soil Sci. 2015, 178, 825–833. [Google Scholar] [CrossRef]
- Liu, Q.; Niu, J.; Liu, Y.; Li, L.; Lv, J. Optimization of lipopeptide biosurfactant production by Bacillus licheniformis L20 and performance evaluation of biosurfactant mixed system for enhanced oil recovery. J. Pet. Sci. Eng. 2022, 208, 109678. [Google Scholar] [CrossRef]
- Gumerova, R.K.; Galitskaya, P.Y.; Badrutdinov, O.R.; Selivanovskaya, S.Y. Changes of hydrocarbon and oil fractions contents in oily waste treated by different methods of bioremediation. Neft. Khozyaystvo-Oil Ind. 2013, 118–120. [Google Scholar]
- Galitskaya, P.Y.; Gumerova, R.K.; Selivanovskaya, S.Y. Bioremediation of oil waste under field experiment. World Appl. Sci. J. 2014, 30, 1694–1698. [Google Scholar] [CrossRef]
- Alsayegh, S.Y.; Al Disi, Z.; Al-Ghouti, M.A.; Zouari, N. Evaluation by MALDI-TOF MS and PCA of the diversity of biosurfactants and their producing bacteria, as adaption to weathered oil components. Biotechnol. Rep. 2021, 31, e00660. [Google Scholar] [CrossRef]
- Zhao, F.; Yuan, M.; Lei, L.; Li, C.; Xu, X. Enhanced production of mono-rhamnolipid in Pseudomonas aeruginosa and application potential in agriculture and petroleum industry. Bioresour. Technol. 2021, 323, 124605. [Google Scholar] [CrossRef]
- Ambust, S.; Das, A.J.; Kumar, R. Bioremediation of petroleum contaminated soil through biosurfactant and Pseudomonas sp. SA3 amended design treatments. Curr. Res. Microb. Sci. 2021, 2, 100031. [Google Scholar] [CrossRef]
- Johnson, P.; Trybala, A.; Starov, V.; Pinfield, V.J. Effect of synthetic surfactants on the environment and the potential for substitution by biosurfactants. Adv. Colloid Interface Sci. 2021, 288, 102340. [Google Scholar] [CrossRef]
- Markande, A.R.; Patel, D.; Varjani, S. A review on biosurfactants: Properties, applications and current developments. Bioresour. Technol. 2021, 330, 124963. [Google Scholar] [CrossRef]
- Nazina, T.N.; Pavlova, N.K.; Tatarkin, Y.V.; Shestakova, N.M.; Babich, T.L.; Sokolova, D.S. Development of the biotechnology for the enhancement of oil recovery from carbonate petroleum reservoirs of the tatarstan republic region. Actual Probl. Oil Gas 2012, 6, 16. [Google Scholar]
- Manga, E.B.; Celik, P.A.; Cabuk, A.; Banat, I.M. Biosurfactants: Opportunities for the development of a sustainable future. Curr. Opin. Colloid Interface Sci. 2021, 56, 101514. [Google Scholar] [CrossRef]
- Rudakova, M.A.; Galitskaya, P.Y.; Selivanovskaya, S.Y. Biosurfactants: Current application trends. Uchenye Zap. Kazan. Univ. Seriya Estestv. Nauk. 2021, 163, 177–208. [Google Scholar] [CrossRef]
- Rani, M.; Weadge, J.T.; Jabaji, S. Isolation and Characterization of Biosurfactant-Producing Bacteria From Oil Well Batteries with Antimicrobial Activities Against Food-Borne and Plant Pathogens. Front. Microbiol. 2020, 11, 64. [Google Scholar] [CrossRef]
- Nalini, S.; Parthasarathi, R. Optimization of rhamnolipid biosurfactant production from Serratia rubidaea SNAU02 under solid-state fermentation and its biocontrol efficacy against Fusarium wilt of eggplant. Ann. Agrar. Sci. 2018, 18, 108–115. [Google Scholar] [CrossRef]
- Marchut-Mikolajczyk, O.; Drożdżyński, P.; Pietrzyk, D.; Antczak, T. Biosurfactant production and hydrocarbon degradation activity of endophytic bacteria isolated from Chelidonium majus L. Microb. Cell Fact. 2018, 17, 171. [Google Scholar] [CrossRef] [Green Version]
- Sachdev, D.P.; Cameotra, S.S. Biosurfactants in agriculture. Appl. Microbiol. Biotechnol. 2013, 97, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Ghosh, U. Production, purification and characterization of biosurfactant isolated from Bacillus oceanisediminis H2. Mater. Today Proc. 2021. [Google Scholar] [CrossRef]
- Wu, J.J.; Chou, H.P.; Huang, J.W.; Deng, W.L. Genomic and biochemical characterization of antifungal compounds produced by Bacillus subtilis PMB102 against Alternaria brassicicola. Microbiol. Res. 2021, 251, 126815. [Google Scholar] [CrossRef]
- Pacwa-Płociniczak, M.; Płaza, G.A.; Piotrowska-Seget, Z.; Cameotra, S.S. Environmental applications of biosurfactants: Recent advances. Int. J. Mol. Sci. 2011, 12, 633–654. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Chen, Y.; Liu, X.; Li, D. Rhamnolipid production from waste cooking oil using newly isolated halotolerant Pseudomonas aeruginosa M4. J. Clean. Prod. 2021, 278, 123879. [Google Scholar] [CrossRef]
- Müller, M.M.; Kügler, J.H.; Henkel, M.; Gerlitzki, M.; Hörmann, B.; Pöhnlein, M.; Syldatk, C.; Hausmann, R. Rhamnolipids-Next generation surfactants? J. Biotechnol. 2012, 162, 366–380. [Google Scholar] [CrossRef] [PubMed]
- Henkel, M.; Hausmann, R. Diversity and Classification of Microbial Surfactants. In Biobased Surfactants; AOCS Press: Urbana, IL, USA, 2019. [Google Scholar]
- Robineau, M.; Le Guenic, S.; Sanchez, L.; Chaveriat, L.; Lequart, V.; Joly, N.; Calonne, M.; Jacquard, C.; Declerck, S.; Martin, P.; et al. Synthetic Mono-Rhamnolipids Display Direct Antifungal Effects and Trigger an Innate Immune Response in Tomato against Botrytis Cinerea. Molecules 2020, 25, 3108. [Google Scholar] [CrossRef]
- Rocha, V.A.L.; de Castilho, L.V.A.; de Castro, R.P.V.; Teixeira, D.B.; Magalhães, A.V.; Gomez, J.G.C.; Freire, D.M.G. Comparison of mono-rhamnolipids and di-rhamnolipids on microbial enhanced oil recovery (MEOR) applications. Biotechnol. Prog. 2020, 64, e2981. [Google Scholar] [CrossRef]
- Kronemberger, F.d.A.; Borges, C.P.; Freire, D.M.G. Fed-Batch Biosurfactant Production in a Bioreactor. Int. Rev. Chem. Eng. 2010, 2, 513–518. [Google Scholar]
- Raza, Z.A.; Khan, M.S.; Khalid, Z.M. Evaluation of distant carbon sources in biosurfactant production by a gamma ray-induced Pseudomonas putida mutant. Process Biochem. 2007, 42, 686–692. [Google Scholar] [CrossRef]
- Gaur, V.K.; Sharma, P.; Sirohi, R.; Varjani, S.; Taherzadeh, M.J.; Chang, J.S.; Yong Ng, H.; Wong, J.W.C.; Kim, S.H. Production of biosurfactants from agro-industrial waste and waste cooking oil in a circular bioeconomy: An overview. Bioresour. Technol. 2022, 343, 126059. [Google Scholar] [CrossRef]
- Tahzibi, A.; Kamal, F.; Assadi, M.M. Improved production of rhamnolipids by a Pseudomonas aeruginosa mutant. Iran. Biomed. J. 2004, 8, 25–31. [Google Scholar]
- Tiso, T.; Zauter, R.; Tulke, H.; Leuchtle, B.; Li, W.J.; Behrens, B.; Wittgens, A.; Rosenau, F.; Hayen, H.; Blank, L.M. Designer rhamnolipids by reduction of congener diversity: Production and characterization. Microb. Cell Fact. 2017, 16, 225. [Google Scholar] [CrossRef] [Green Version]
- Cha, M.; Lee, N.; Kim, M.; Kim, M.; Lee, S.-J. Heterologous production of Pseudomonas aeruginosa EMS1 biosurfactant in Pseudomonas putida. Bioresour. Technol. 2008, 99, 2192–2199. [Google Scholar] [CrossRef]
- Poonguzhali, P.; Rajan, S.; Parthasarathi, R.; Srinivasan, R.; Kannappan, A. Optimization of biosurfactant production by Pseudomonas aeruginosa using rice water and its competence in controlling Fusarium wilt of Abelmoschus esculentus. S. Afr. J. Bot. 2021, in press, corrected proof. [Google Scholar] [CrossRef]
- Ángeles, M.T.; Refugio, R.V. In situ biosurfactant production and hydrocarbon removal by Pseudomonas putida CB-100 in bioaugmented and biostimulated oil-contaminated soil. Braz. J. Microbiol. 2013, 44, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Janek, T.; Lukaszewicz, M.; Krasowska, A. Identification and characterization of biosurfactants produced by the Arctic bacterium Pseudomonas putida BD2. Colloids Surf B Biointerfaces 2013, 110, 379–386. [Google Scholar] [CrossRef]
- Ray, M.; Kumar, V.; Banerjee, C. Kinetic modelling, production optimization, functional characterization and phyto-toxicity evaluation of biosurfactant derived from crude oil biodegrading Pseudomonas sp. IITISM 19. J. Environ. Chem. Eng. 2022, 10, 107190. [Google Scholar] [CrossRef]
- Da Silva, R.C.; Luna, J.M.; Rufino, R.D.; Sarubbo, L.A. Ecotoxicity of the formulated biosurfactant from Pseudomonas cepacia CCT 6659 and application in the bioremediation of terrestrial and aquatic environments impacted by oil spills. Process Saf. Environ. Prot. 2021, 154, 338–347. [Google Scholar] [CrossRef]
- Koutinas, M.; Kyriakou, M.; Kostas, A.; Hadjicharalambous, M.; Kaliviotis, E.; Pasias, D.; Kazamias, G.; Varavvas, C.; Vyrides, I. Enhanced biodegradation and valorization of drilling wastewater via simultaneous production of biosurfactants and polyhydroxyalkanoates by Pseudomonas citronellolis SJTE-3. Bioresour. Technol. 2021, 340, 125679. [Google Scholar] [CrossRef]
- Sun, W.; Cao, W.; Jiang, M.; Saren, G.; Liu, J.; Cao, J.; Ali, I.; Yu, X.; Peng, C.; Naz, I. Isolation and characterization of biosurfactant-producing and diesel oil degrading Pseudomonas sp. CQ2 from Changqing oil field, China. RSC Adv. 2018, 8, 39710–39720. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Liu, Y. Remediation technologies for oil-contaminated sediments. Mar. Pollut. Bull. 2015, 101, 483–490. [Google Scholar] [CrossRef]
- Galitskaya, P.; Biktasheva, L.; Blagodatsky, S.; Selivanovskaya, S. Response of bacterial and fungal communities to high petroleum pollution in different soils. Sci. Rep. 2021, 11, 18. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 45e. [Google Scholar] [CrossRef]
- Zargar, A.N.; Lymperatou, A.; Skiadas, I.; Kumar, M.; Srivastava, P. Structural and functional characterization of a novel biosurfactant from Bacillus sp. IITD106. J. Hazard. Mater. 2022, 423, 127201. [Google Scholar] [CrossRef] [PubMed]
- Aboelkhair, H.; Diaz, P.; Attia, A. Biosurfactant production using Egyptian oil fields indigenous bacteria for microbial enhanced oil recovery. J. Pet. Sci. Eng. 2022, 208, 109601. [Google Scholar] [CrossRef]
- Liu, Q.; Niu, J.; Yu, Y.; Wang, C.; Lu, S.; Zhang, S.; Lv, J.; Peng, B. Production, characterization and application of biosurfactant produced by Bacillus licheniformis L20 for microbial enhanced oil recovery. J. Clean. Prod. 2021, 307, 127193. [Google Scholar] [CrossRef]
- Tuleva, B.K.; Ivanov, G.R.; Christova, N.E. Biosurfactant Production by a New Pseudomonas Putida Strain. Z. Nat. C 2002, 57, 356–360. [Google Scholar] [CrossRef]
- Deepika, K.V.; Raghuram, M.; Bramhachari, P.V. Rhamnolipid biosurfactant production by Pseudomonas aeruginosa strain KVD-HR42 isolated from oil contaminated mangrove sediments. Afr. J. Microbiol. Res. 2017, 11, 218–231. [Google Scholar] [CrossRef] [Green Version]
- Leitermann, F.; Syldatk, C.; Hausmann, R. Fast quantitative determination of microbial rhamnolipids from cultivation broths by ATR-FTIR Spectroscopy. J. Biol. Eng. 2008, 2, 13. [Google Scholar] [CrossRef] [Green Version]
- Sabturani, N.; Latif, J.; Radiman, S.; Hamzah, A. Spectroscopic analysis of rhamnolipid produced by produced by Pseudomonas aeruginosa UKMP14T. Malays. J. Anal. Sci. 2016, 20, 31–43. [Google Scholar] [CrossRef]
- Al-Anssari, S.; Arain, Z.-U.-A.; Shanshool, H.A.; Ali, M.; Keshavarz, A.; Iglauer, S.; Sarmadivaleh, M. Effect of Nanoparticles on the Interfacial Tension of CO2-Oil System at High Pressure and Temperature: An Experimental Approach. In Proceedings of the SPE Asia Pacific Oil & Gas Conference and Exhibition, Virtual, 17–19 November 2020. [Google Scholar]
- Bai, Y.; Wang, F.; Shang, X.; Lv, K.; Dong, C. Microstructure, dispersion, and flooding characteristics of intercalated polymer for enhanced oil recovery. J. Mol. Liq. 2021, 340, 117235. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biktasheva, L.; Gordeev, A.; Selivanovskaya, S.; Galitskaya, P. Di- and Mono-Rhamnolipids Produced by the Pseudomonas putida PP021 Isolate Significantly Enhance the Degree of Recovery of Heavy Oil from the Romashkino Oil Field (Tatarstan, Russia). Processes 2022, 10, 779. https://doi.org/10.3390/pr10040779
Biktasheva L, Gordeev A, Selivanovskaya S, Galitskaya P. Di- and Mono-Rhamnolipids Produced by the Pseudomonas putida PP021 Isolate Significantly Enhance the Degree of Recovery of Heavy Oil from the Romashkino Oil Field (Tatarstan, Russia). Processes. 2022; 10(4):779. https://doi.org/10.3390/pr10040779
Chicago/Turabian StyleBiktasheva, Liliya, Alexander Gordeev, Svetlana Selivanovskaya, and Polina Galitskaya. 2022. "Di- and Mono-Rhamnolipids Produced by the Pseudomonas putida PP021 Isolate Significantly Enhance the Degree of Recovery of Heavy Oil from the Romashkino Oil Field (Tatarstan, Russia)" Processes 10, no. 4: 779. https://doi.org/10.3390/pr10040779