

The Difference of Lead Accumulation and Transport in Different Ecotypes of Miscanthus floridulus
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
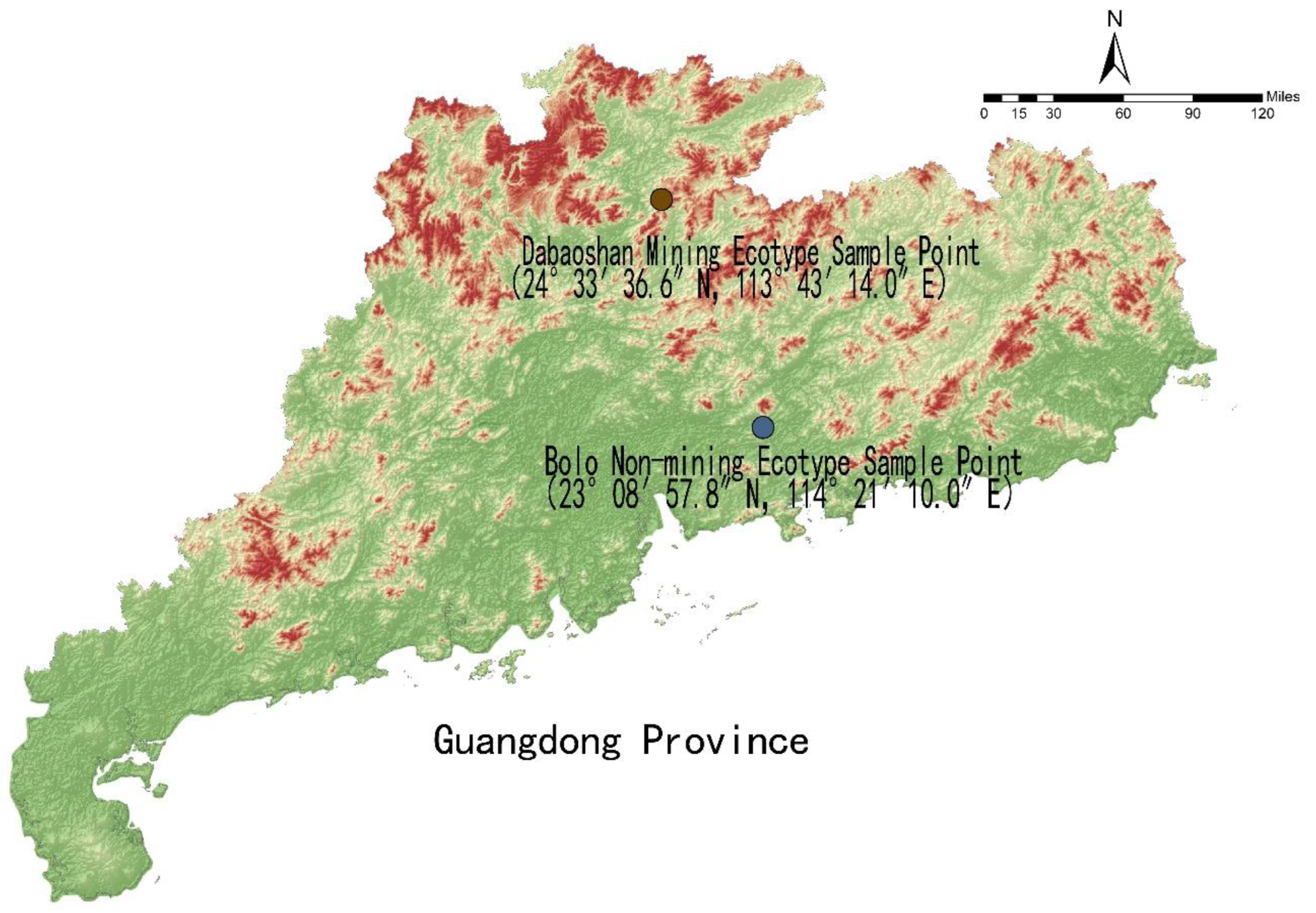
2.1. Test Materials
2.2. Test Soil
2.3. Test Design
2.4. Sample Analysis
2.5. Data Processing and Statistical Analysis
- (1)
- Tolerance index (TI), According to the method laid out by Metwall et al. [32], the tolerance index was calculated to evaluate the tolerance degree of M. floridulus to Pb. TI is defined as:
- (2)
- Translocation factor (TF), This represents the capability for Pb transport from root system to stem and leaf [33]. TF is defined as:
3. Results
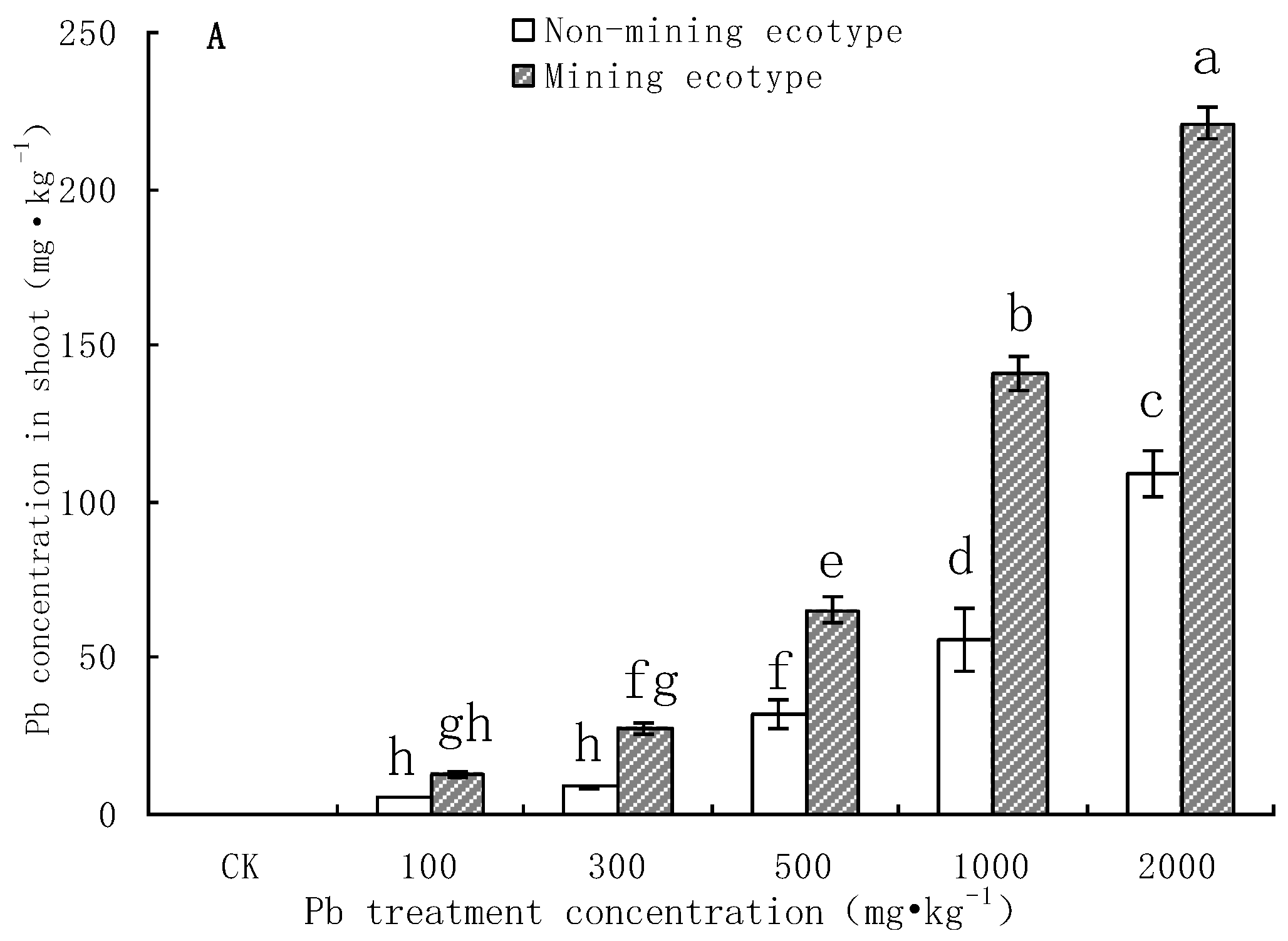
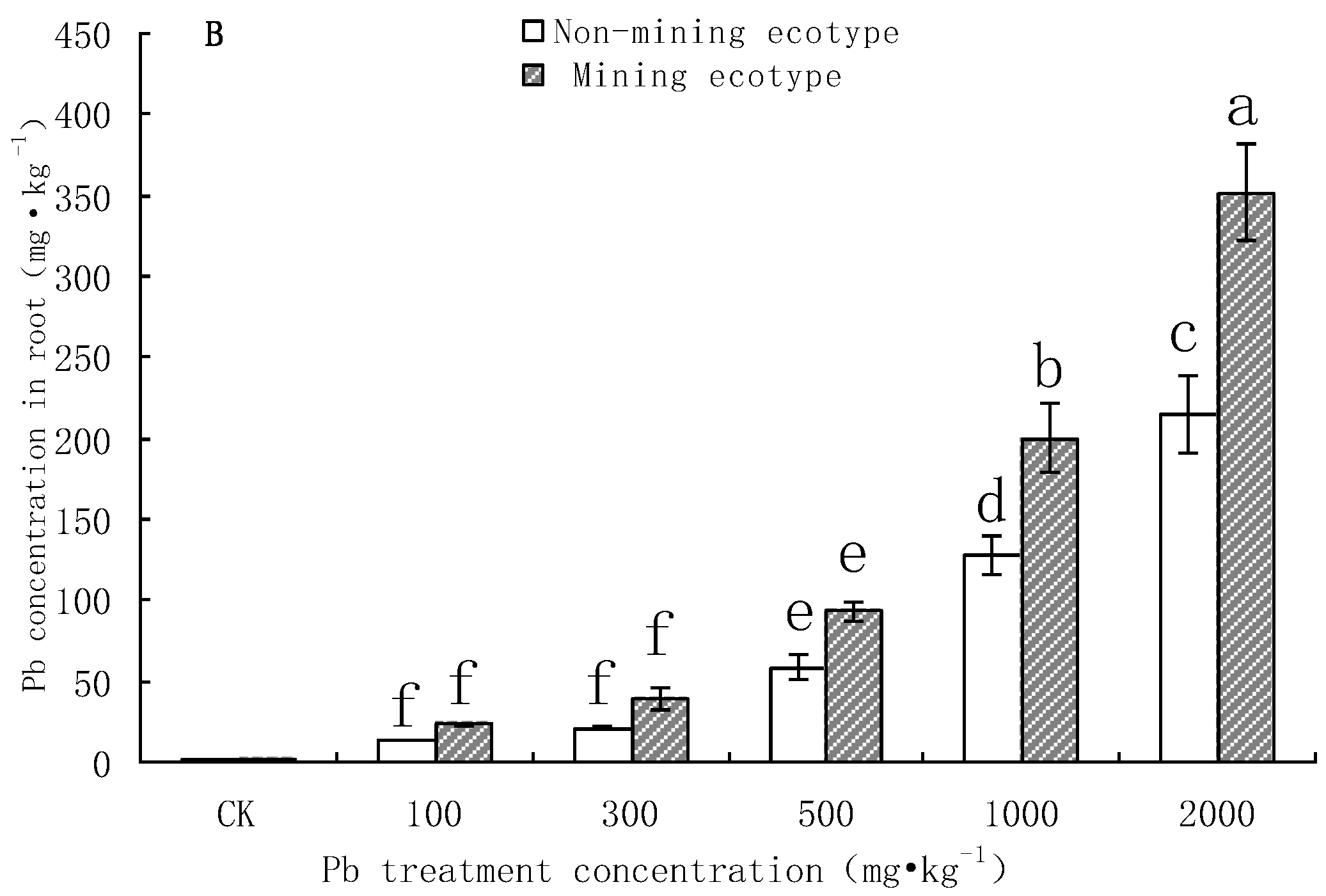
3.1. Pb Content in the Body of M. floridulus under Pb Stress
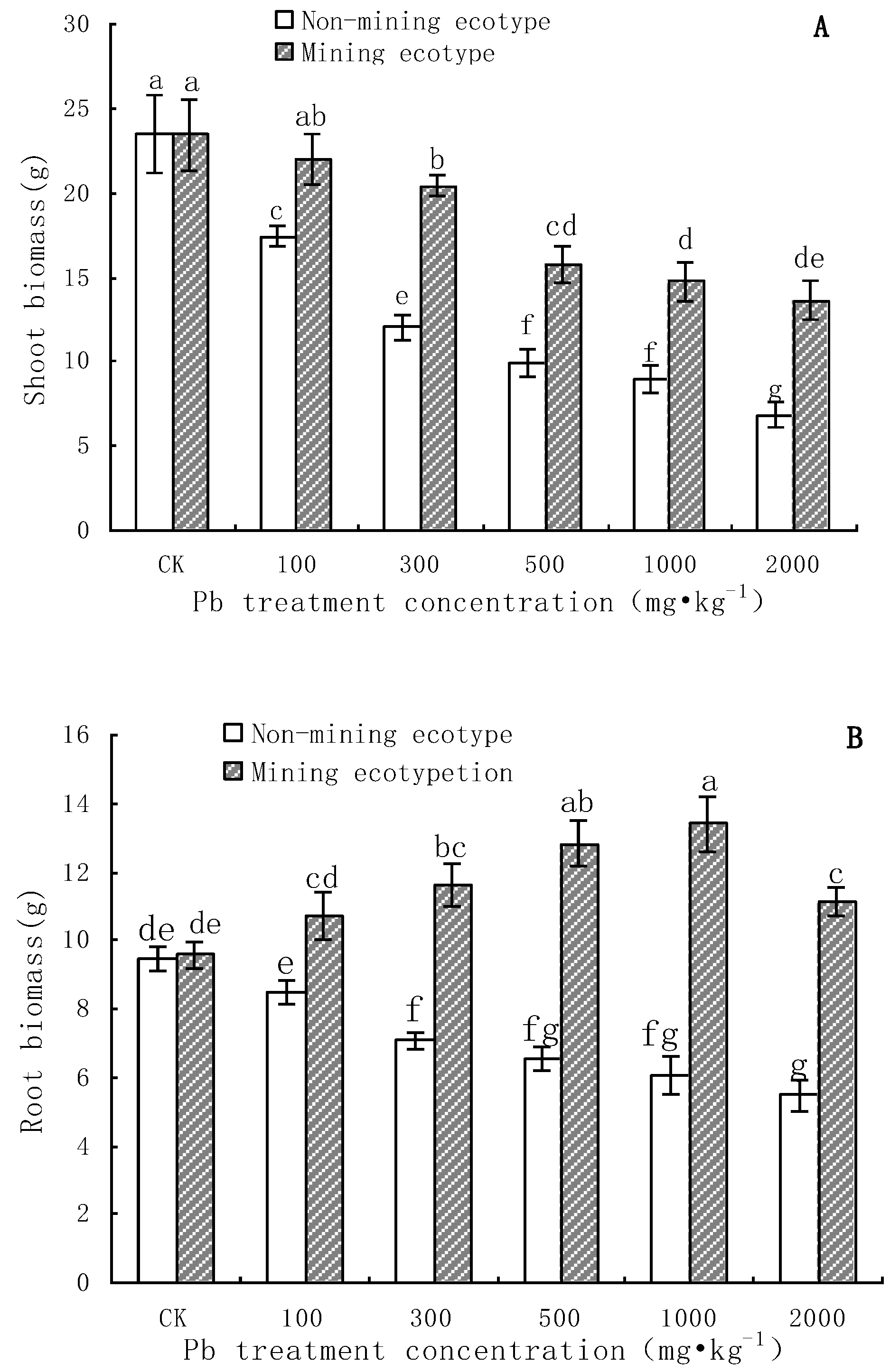
3.2. Aboveground and Root Biomass of M. floridulus under Pb Stress
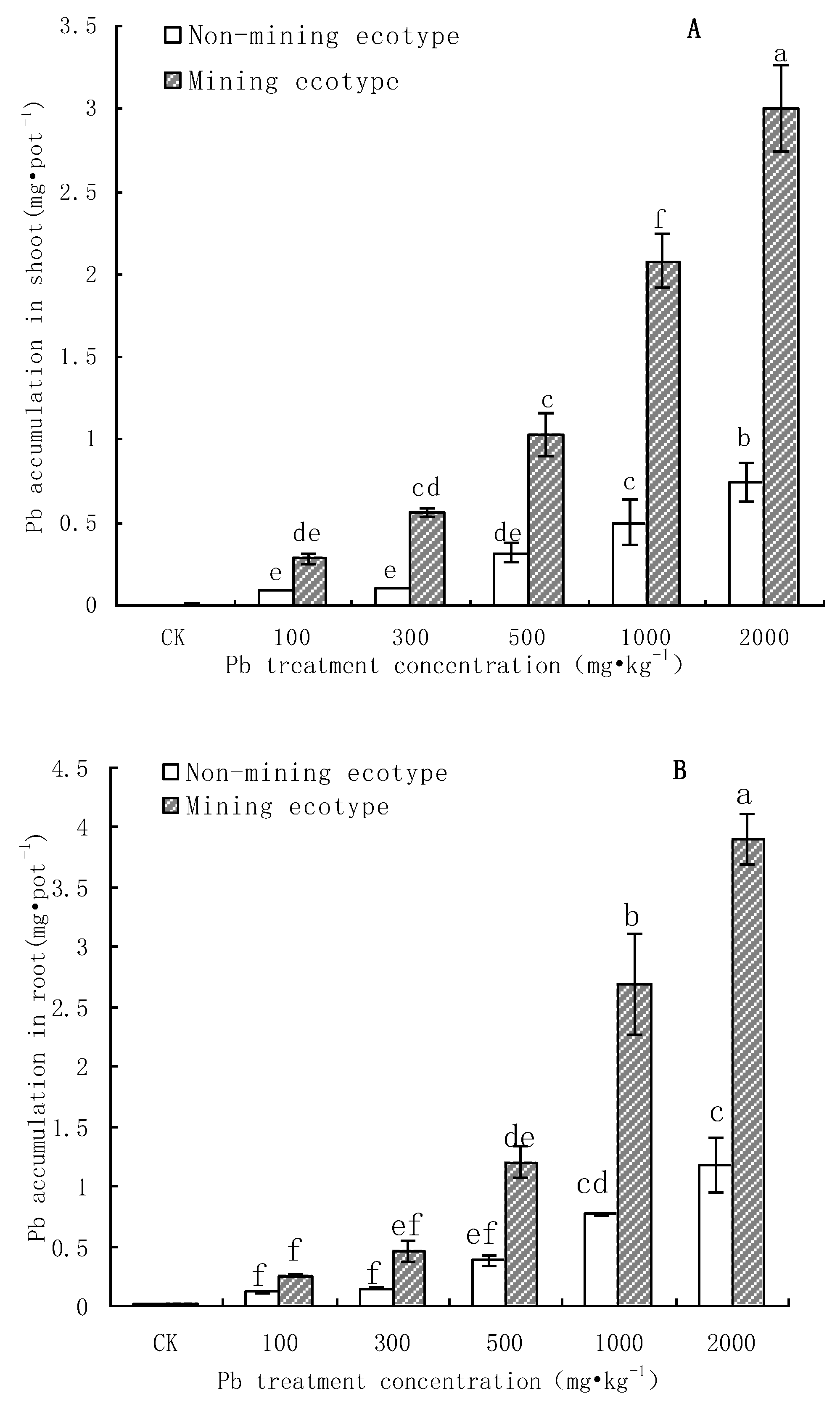
3.3. Pb Accumulation in M. floridulus under Pb Stress
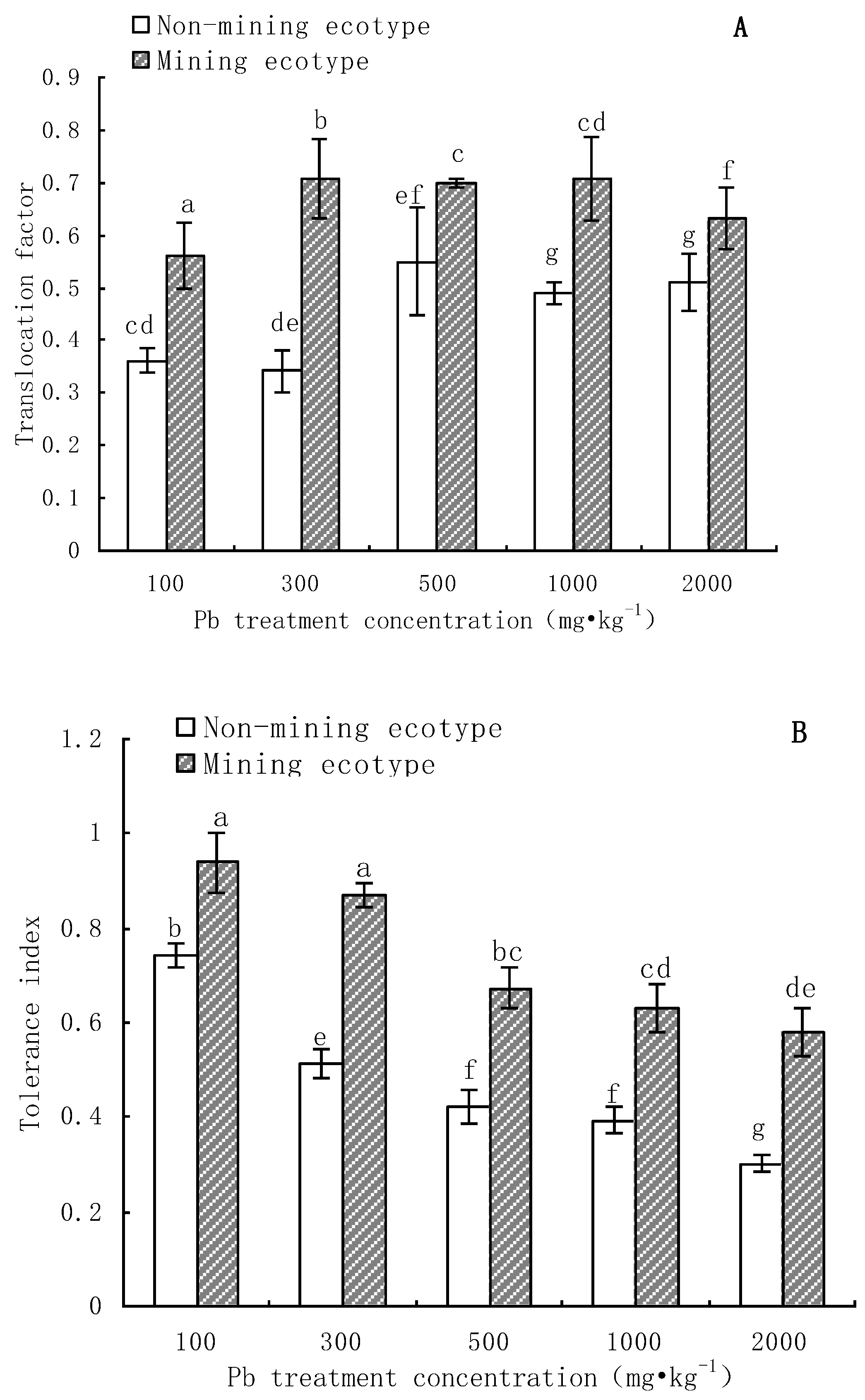
3.4. Transfer Coefficient and Tolerance Index of M. floridulus under Pb Stress
3.5. Variation of Pb Content of NH4OAc Extraction in Soil
3.6. Correlation Analysis of Biomass, Pb Content, TF and TI of M. floridulus
4. Discussion
5. Conclusions
- (1)
- The aboveground and root biomass of the mining ecotype M. floridulus was significantly less affected than that of the non-mining ecotype by Pb concentration in soil. There were also significant differences in Pb uptake, transport, and accumulation between the two ecotypes of M. floridulus. Under the same Pb treatment level, the Pb content and accumulation in the shoots and roots of ecotypes in mining areas were significantly higher than in ecotypes in non-mining areas; the mining ecotype plants were more tolerant to environmental Pb stress than the non-mining ecotype, which is a Pb-stress-tolerant ecotype.
- (2)
- The Pb content in roots was higher than that in shoots, and the translocation factor (TF) was less than 0.7, which indicates that the two ecotypes of M. floridulus not only have a strong ability to absorb Pb from soil, but also can retain a large amount of Pb in the roots. Only a small amount of Pb absorbed by the roots is transferred to the upper part of the ground, thus reducing the toxicity of Pb to the plant.
- (3)
- The amount of NH4OAc-extracted Pb in the soil planted with the mining ecotype plants M. floridulus decreased significantly, which may be due to the strong ability of the roots of this ecotype to absorb Pb, leading to the decrease of the content of available Pb in the soil.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hector, A.; Bagchi, R. Biodiversity and ecosystem multifunctionality. Nature 2007, 448, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.D.J.; Neufeld, J.D. Ecology and exploration of the rare biosphere. Nature Reviews Microbiology. Nat. Rev. Micrlbiol. 2015, 13, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.H.; Luo, S.H.; Luo, H.Y.; Huang, H.Y.; Xu, F.; Feng, S.; Xu, H. Improved phytoremediation of heavy metal contaminated soils by Miscanthus floridulus under a varied rhizosphere ecological characteristic. Sci. Total Environ. 2021, 808, 151995. [Google Scholar] [CrossRef]
- Nie, G.; Zhong, M.Y.; Cai, J.B.; Yang, X.Y.; Zhou, J.; Appiah, C.; Tang, M.Y.; Wang, X.; Feng, G.Y.; Huang, L.K.; et al. Transcriptome characterization of candidate genes related to chromium uptake, transport and accumulation in Miscanthus sinensis. Ecotoxicol. Environ. Saf. 2021, 221, 112445. [Google Scholar] [CrossRef]
- Li, Q.F.; Du, W.B.; Li, Z.A.; Wang, Z.F.; Peng, S.L. Heavy metals accumulation in mining area’s Miscanthus sinensis populations and its relationship with soil characters. Chin. J. Ecol. 2006, 25, 255–258. [Google Scholar]
- Lenka, Š.; Juraj, F.; Danica, F. Transfer of Potentially Toxic Elements in the Soil-Plant System in Magnesite Mining and Processing Areas. Processes 2022, 10, 720. [Google Scholar]
- Xu, J.; Wang, S.; Yao, T.; She, X.; Gan, Z. Vertical Distributions and Bioavailabilities of Heavy Metals in Soil in An-Tea Plantations in Qimen County, China. Processes 2022, 10, 664. [Google Scholar] [CrossRef]
- Ke, W.S.; Xi, H.A.; Yang, Y. Analysis on characteristics of phytogeochemistry of Elsholtzia haichowensisin Daye Tonglushan copper mine. Acta Ecol. Sin. 2001, 21, 907–912. [Google Scholar]
- Xie, M.J.; Ke, W.S.; Wang, W.X. MDA accumulation and antioxidation capacity of two Elsholtzia splendens populations under copper stress. Chin. J. Ecol. 2005, 24, 935–938. [Google Scholar]
- Liu, Y.; Fang, Z.; Xie, C.; Zhang, N. Physiological responses of mining ecotypes and non-mining ecotypes of centipede grass to lead stress. J. Logist. Eng. Coll. 2014, 30, 52–58. (In Chinese) [Google Scholar]
- Wen, C.H.; Duan, C.Q.; Chang, X.X. Differentiation in Datura stramonium L.populations exposed to heavy-metal pollution at different durations: RAPD analysis. Acta Ecol. Sin. 2001, 21, 1239–1245. [Google Scholar]
- Peng, S.L.; Du, W.B.; Li, Z.A. A review of heavy metal accumulation and tolerance by plants of different ecotype. J. Jishou Univ. 2004, 25, 19–26. [Google Scholar]
- Yu, H.; Zheng, X.; Weng, W.; Yan, X.; Chen, P.; Liu, X.; Peng, T.; Zhong, Q.; Xu, K.; Wang, C.; et al. Synergistic effects of antimony and arsenic contaminations on bacterial, archaeal and fungal communities in the rhizosphere of Miscanthus sinensis: Insights for nitrification and carbon mineralization. J. Hazard. Mater. 2021, 411, 125094. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, B.; Boléo, S.; Sidella, S.; Costa, J.; Duarte, M.P.; Mendes, B.; Cosentino, S.L.; Fernando, A.L. Phytoremediation of heavy metal-contaminated soils using the perennial energy crops miscanthus spp. And Arundo donax L. Bioenerg. Res. 2015, 8, 1500–1511. [Google Scholar] [CrossRef]
- Zadel, U.; Nesme, J.; Michalke, B.; Vestergaard, G.; Plaza, G.A.; Schroder, P.; Radl, V.; Schloter, M. Changes induced by heavy metals in the plant-associated microbiome of miscanthus x giganteus. Sci. Total Environ. 2020, 711, 134433. [Google Scholar] [CrossRef]
- Wu, B.; Peng, H.; Sheng, M.; Luo, H.; Wang, X.; Zhang, R.; Xu, F.; Xu, H. Evaluation of phytoremediation potential of native dominant plants and spatial distribution of heavy metals in abandoned mining area in Southwest China. Ecotoxicol. Environ. Saf. 2021, 220, 112368. [Google Scholar] [CrossRef]
- Wu, B.; Luo, H.; Wang, X.; Liu, H.; Peng, H.; Sheng, M.; Xu, F.; Xu, H. Effects of environmental factors on soil bacterial community structure and diversity in different contaminated districts of Southwest China mine tailings. Sci. Total Environ. 2022, 802, 149899. [Google Scholar] [CrossRef]
- Ren, L.M.; Liu, P.; Zheng, Q.E. A survey of heavy metal content of plants growing on the soil polluted by manganese mine in Daxin County, Guangxi. Subtrop. Plant Sci. 2006, 35, 5–8. [Google Scholar]
- Sun, J.; Tie, B.Q.; Qin, P.F. Investigation of contaminated soil and plants by heavy metals in Pb-Zn mining area. J. Plant Resour. Environ. 2006, 15, 63–67. [Google Scholar]
- Chen, Z.J.; Tian, W.; Li, Y.J.; Sun, L.N.; Chen, Y.; Zhang, H.; Li, Y.Y.; Han, H. Responses of rhizosphere bacterial communities, their functions and their network interactions to Cd stress under phytostabilization by Miscanthus spp. Environ. Pollut. 2021, 287, 117663. [Google Scholar] [CrossRef]
- Zhu, Y.G. Microinterface processes in soil-plant systems and their eco-environmental effects. J. Environ. Sci. 2003, 23, 205–210. [Google Scholar]
- Wang, H.B.; Shu, W.S.; Lan, C. Ecology for heavy metal pollution: Recent advances and future prospects. Acta Ecol. Snica 2005, 25, 596–605. [Google Scholar]
- Zhou, S.; Deng, R.; Hursthouse, A. Risk Assessment of Potentially Toxic Elements Pollution from Mineral Processing Steps at Xikuangshan Antimony Plant, Hunan, China. Processes 2020, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Zhou, Y.; Zhao, Y.; Gao, Q.Z.; Peng, X.Z.; Dang, Z.; Zhang, C.B.; Yang, X.Q.; Yang, Z.J.; Dou, L.; et al. Study on heavy metals in soils contaminated by acid mine drainage from Dabaoshan Mine, Guangdong. Environ. Sci. 2007, 28, 805–812. [Google Scholar]
- Qin, J.Q.; Xia, B.C.; Hu, M.; Zhao, P.; Zhao, H.R.; Lin, X.F. Analysis of the vegetation succession of tailing wasteland of Dabaoshan Mine, Guangdong Province. J. Agro-Environ. Sci. 2009, 28, 2085–2091. [Google Scholar]
- Qin, J.Q.; Zhao, H.R.; Dai, M.; Zhao, P.; Chen, X.; Liu, H.; Lu, B.Z. Speciation Distribution and Influencing Factors of Heavy Metals in Rhizosphere Soil of Miscanthus Floridulus in the Tailing Reservoir Area of Dabaoshan Iron Polymetallic Mine in Northern Guangdong. Processes 2022, 10, 1217. [Google Scholar] [CrossRef]
- Zhao, H.R.; Xia, B.C.; Qin, J.Q.; Zhang, J. Hydrogeochemical and mineralogical characteristics related to heavy metal attenuation in a stream polluted by acid mine drainage: A case study in Dabaoshan mine, China. J. Environ. Sci. 2012, 24, 979–989. [Google Scholar] [CrossRef]
- Zhao, H.R.; Xia, B.C.; Fan, C.; Zhao, P.; Shen, S. Human health risk from soil heavy metal contamination under different land uses near Dabaoshan mine, southern China. Sci. Total Environ. 2012, 417, 45–54. [Google Scholar] [CrossRef]
- Cui, J.L.; Luo, C.L.; Tang, C.W.; Chang, T.; Li, X. Speciation and leaching of trace metal contaminants from e-waste contaminated soils. J. Hazard. Mater. 2017, 329, 150–158. [Google Scholar] [CrossRef]
- Lu, R. Methods of Soil and Agricultural Chemistry; Beijing Science and Technology Press: Beijing, China, 1999; pp. 235–285. [Google Scholar]
- Wenzel, W.W.; Jockwer, F. Accumulation of heavy metals in plants grown on mineralized soils of the Austrian Alps. Environ. Pollut. 1999, 104, 145–155. [Google Scholar] [CrossRef]
- Metwally, A.; Safronova, V.I.; Belimov, A.A.; Dietz, K.J. Genotypic variation of the response to cadmium toxicity in Pisum sativum L. J. Exp. Bot. 2005, 56, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Stoltz, E.; Greger, M. Accumulation properties ofAs, Cd, Cu, Pb and Zn by four wetland plant species growing on submerged mine tailing. Environ. Exp. Bot. 2002, 47, 271–280. [Google Scholar] [CrossRef]
- Brun, L.A.; Maillet, J.; Hinsinge, P.; Pepin, M. Evalution of copper availability to plants in copper-contaminated vineyard soils. Environ. Pollut. 2001, 111, 293–302. [Google Scholar] [CrossRef]
- Li, J.Y.; Zheng, B.H.; He, Y.Z.; Zhou, Y.Y.; Chen, X.; Ruan, S.; Yang, Y.; Dai, C.H.; Tang, L. Antimony contamination, consequences and removal techniques: A review. Ecotoxicol. Environ. Saf. 2018, 156, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.H.; Yang, X.E.; Ye, Z.Q.; He, B. Southeast scene days to respond to the growth of cadmium, lead and accumulation characteristics comparison. J. Northwest Agric. For. Univ. Sci. Technol. 2004, 32, 101–106. [Google Scholar]
- Duan, C.Q. Adaptation and microevolution of plants on contaminated environment. Chin. J. Ecol. 1995, 14, 43–50. [Google Scholar]
- Han, Y.-Y.; Zhou, S.; Chen, Y.-H.; Kong, X.; Xu, Y.; Wang, W. The involvement of expansins in responses to phosphorus availability in wheat, and its potentials in improving phosphorus efficiency of plants. Plant Physiol. Biochem. 2014, 78, 53–62. [Google Scholar] [CrossRef]
- Fayiga, A.O.; Ma, L.Q.; Cao, X.; Rathinasabapathi, B. Effects of heavy metals on growth and arsenic accumulation in the arsenic hyperaccumulator Pteris vittata L. Environ. Pollut. 2004, 132, 289–296. [Google Scholar] [CrossRef]
- He, J.; Li, H.; Luo, J.; Ma, C.; Li, S.; Qu, L.; Gai, Y.; Jiang, X.; Janz, D.; Polle, A. A transcriptomic network underlies microstructural and physiological responses to cadmium in Populus×canescens. Plant Physiol. 2013, 162, 424–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.T. Pollution-Resistant Evolution in Plants and Its Genecological Costs. J. Ecol. 1997, 16, 53–57. [Google Scholar]
- Macnair, M.R. The genetics of metal tolerance on vascular plants. New Phytol. 1993, 124, 541–559. [Google Scholar] [CrossRef]
- Jiang, L.Y.; Yang, X.E.; Shi, W.Y.; Ye, Z.Q.; He, Z.L. Copper uptake and tolerance in two contrasting ecotypes of Elsholtzia argyi. J. Plant Nutr. 2004, 27, 2067–2083. [Google Scholar] [CrossRef]
- Zlobin, I.E.; Kartashov, A.V.; Shpakovski, G.V. Different roles of glutathione in copper and zinc chelation in Brassica napus roots. Plant Physiol. Biochem. 2017, 118, 333–341. [Google Scholar] [CrossRef]
- Poschenrieder, C.; i Coll, J.B. Phytoremediation: Principles and perspectives. Contrib. Sci. 2003, 2, 333–344. [Google Scholar]
- Baker, A.J. Accumulators and excluders-strategies in the response of plants to heavy metals. J. Plant Nutr. 1981, 3, 643–654. [Google Scholar] [CrossRef]
- Yang, Q.; Tu, S.; Wang, G.; Liao, X.; Yan, X. Effectiveness of applying arsenate reducing bacteria to enhance arsenic removal from polluted soils by Pteris vittata L. Int. J. Phytoremediation 2012, 14, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhou, Z.; Rao, M.; Sun, Z. Assessment of heavy metals and arsenic pollution in surface sediments from rivers around a uranium mining area in East China. Environ. Geochem. Health 2020, 42, 1401–1413. [Google Scholar] [CrossRef] [PubMed]
- Xiao, E.; Ning, Z.; Xiao, T.; Sun, W.; Qiu, Y.; Zhang, Y.; Chen, J.; Gou, Z.; Chen, Y. Variation in rhizosphere microbiota correlates with edaphic factor in an abandoned antimony tailing dump. Environ. Pollut. 2019, 253, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yuan, Y.; Jiao, H.; Liu, X.; Su, S.; Tian, S. Study on heavy metal absorption and enrichment characteristics by 8 plants species settled naturally in Xiangsigu copper tailings. Ecol. Environ. Sci. 2015, 5, 25. [Google Scholar]
- Zhu, H.; Teng, Y.; Wang, X.; Zhao, L.; Ren, W.; Luo, Y.; Christie, P. Changes in clover rhizosphere microbial community and diazotrophs in mercury-contaminated soils. Sci.Total Environ. 2021, 767, 145473. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Sample Point | Organic C (mg·kg−1) | Available P (mg·kg−1) | Available N (mg·kg−1) | Heavy Metal Contents (mg·kg−1) | |||
|---|---|---|---|---|---|---|---|
| Zn | Pb | Cu | Cd | ||||
| Dabaoshan Mining Area | 14.7 ± 0.9 b | 32.2 ± 2.0 b | 30.2 ± 1.9 b | 1768.7 ± 91.1 a | 1253.3 ± 71.3 a | 1701.3 ± 77.5 a | 9.1 ± 0.9 a |
| Boluo County | 13.8 ± 0.9 b | 26.6 ± 1.8 b | 28.4 ± 2.7 b | 135.2 ± 13.1 b | 242.6 ± 44.1 b | 48.4 ± 9.5 b | 1.1 ± 0.2 b |
| Soil samples tested | 36.2 ± 1.1 a | 60.5 ± 9.9 a | 61.5 ± 10.9 a | 60.5 ± 9.9 b | 35.2 ± 7.2 c | 6.3 ± 1.7 b | 0.13 ± 0.1 b |
| Pb Treatment Concentration (mg·kg−1) | Before Planting | After Harvest | |
|---|---|---|---|
| Non-Mining Ecotype | Mining Ecotype | ||
| CK | 1.209 ± 0.101 a | 0.13 ± 0.01 b | 0.12 ± 0.000 b |
| 100 | 30.060 ± 0.77 a | 12.098 ± 0.199 b | 9.799 ± 0.300 c |
| 300 | 120.094 ± 6.032 a | 63.163 ± 6.205 b | 53.509 ± 4.160 c |
| 500 | 152.877 ± 8.093 a | 78.396 ± 8.105 b | 66.66 ± 5.103 c |
| 1000 | 317.817 ± 18.166 a | 159.500 ± 11.127 b | 127.785 ± 11.100 c |
| 2000 | 788.833 ± 30.955 a | 442.530 ± 20.951 b | 254.674 ± 38.852 c |
| Item | Shoot Biomass | Root Biomass | Pb Content in Shoot | Pb Content in Root | Pb Accumulation in Shoot | Pb Accumulation in Root | Translocation Factor (TF) |
|---|---|---|---|---|---|---|---|
| Root biomass | 0.551 ** | 1 | |||||
| Pb content in shoot | −0.481 ** | 0.194 | 1 | ||||
| Pb content in root | −0.508 ** | 0.109 | 0.967 ** | 1 | |||
| Pb accumulation in shoot | −0.299 | 0.428 ** | 0.952 ** | 0.919 ** | 1 | ||
| Pb accumulation in root | −0.329 | 0.369 * | 0.945 ** | 0.949 ** | 0.983 ** | 1 | |
| Translocation factor (TF) | 0.936 ** | 0.459 ** | 0.548 ** | −0.598 ** | −0.393 * | −0.418 * | 1 |
| Tolerance index (TI) | 0.993 ** | 0.558 ** | −0.483 ** | −0.505 ** | −0.299 | −0.316 | 0.943 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, J.; Zhao, H.; Liu, H.; Dai, M.; Zhao, P.; Chen, X.; Wu, X. The Difference of Lead Accumulation and Transport in Different Ecotypes of Miscanthus floridulus. Processes 2022, 10, 2219. https://doi.org/10.3390/pr10112219
Qin J, Zhao H, Liu H, Dai M, Zhao P, Chen X, Wu X. The Difference of Lead Accumulation and Transport in Different Ecotypes of Miscanthus floridulus. Processes. 2022; 10(11):2219. https://doi.org/10.3390/pr10112219
Chicago/Turabian StyleQin, Jianqiao, Huarong Zhao, Hao Liu, Min Dai, Peng Zhao, Xi Chen, and Xiange Wu. 2022. "The Difference of Lead Accumulation and Transport in Different Ecotypes of Miscanthus floridulus" Processes 10, no. 11: 2219. https://doi.org/10.3390/pr10112219