
The Broad-Spectrum Antitrypanosomal Inhibitory Efficiency of the Antimetabolite/Anticancer Drug Raltitrexed


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Evaluation of Trypanocidal Activity In Vitro
2.1.1. Materials
2.1.2. Antitrypanosomal Assay
2.2. Docking Studies
2.2.1. Macromolecule and Compounds Retrieval and Preparation
2.2.2. Docking Runs
2.3. Molecular Dynamics Simulation
2.4. Statistical Analysis
3. Results and Discussion
3.1. Trypanocidal Assay
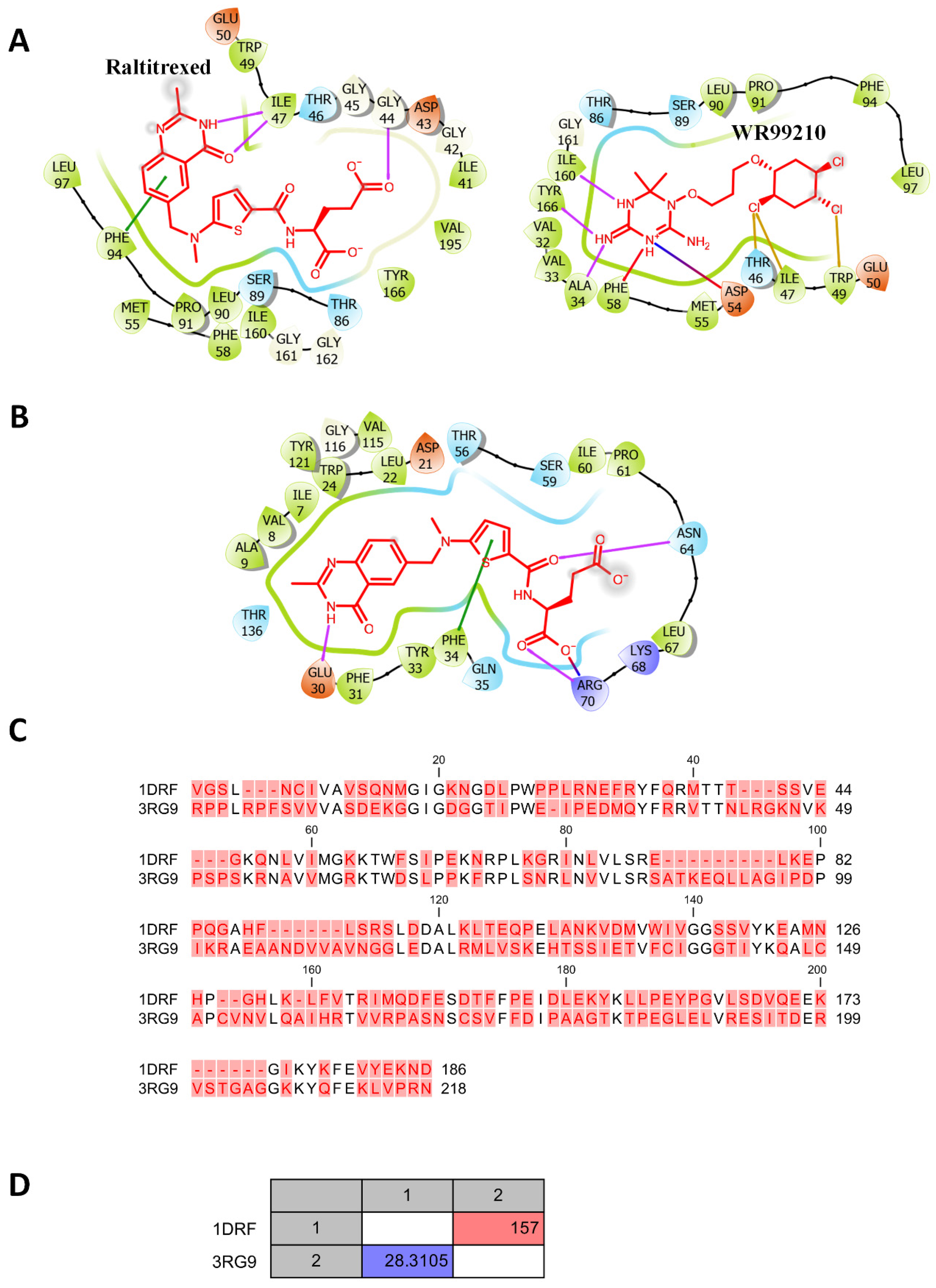
3.2. Molecular Docking
3.3. Molecular Dynamics Simulations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaye, S.B. New antimetabolites in cancer chemotherapy and their clinical impact. Br. J. Cancer 1998, 78, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gangjee, A.; Jain, H.D.; Phan, J.; Lin, X.; Song, X.; McGuire, J.J.; Kisliuk, R.L. Dual inhibitors of thymidylate synthase and dihydrofolate reductase as antitumor agents: Design, synthesis, and biological evaluation of classical and nonclassical pyrrolo [2, 3-d] pyrimidine antifolates. J. Med. Chem. 2006, 49, 1055–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimondi, M.V.; Randazzo, O.; La Franca, M.; Barone, G.; Vignoni, E.; Rossi, D.; Collina, S. DHFR inhibitors: Reading the past for discovering novel anticancer agents. Molecules 2019, 24, 1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wróbel, A.; Arciszewska, K.; Maliszewski, D.; Drozdowska, D. Trimethoprim and other nonclassical antifolates an excellent template for searching modifications of dihydrofolate reductase enzyme inhibitors. J. Antibiot. 2020, 73, 5–27. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Scocchera, E.W.; Wright, D.L.; Anderson, A.C. Antifolates as effective antimicrobial agents: New generations of trimethoprim analogs. MedChemComm 2013, 4, 908–915. [Google Scholar] [CrossRef]
- He, J.; Qiao, W.; An, Q.; Yang, T.; Luo, Y. Dihydrofolate reductase inhibitors for use as antimicrobial agents. Eur. J. Med. Chem. 2020, 195, 112268. [Google Scholar] [CrossRef]
- Cerny, N.; Bivona, A.E.; Sanchez Alberti, A.; Trinitario, S.N.; Morales, C.; Cardoso Landaburu, A.; Cazorla, S.I.; Malchiodi, E.L. Cruzipain and Its Physiological Inhibitor, Chagasin, as a DNA-Based Therapeutic Vaccine Against Trypanosoma cruzi. Front. Immunol. 2020, 11, 565142. [Google Scholar] [CrossRef]
- Kennedy, P.G. Clinical features, diagnosis, and treatment of human African trypanosomiasis (sleeping sickness). Lancet Neurol. 2013, 12, 186–194. [Google Scholar] [CrossRef]
- Kourbeli, V.; Chontzopoulou, E.; Moschovou, K.; Pavlos, D.; Mavromoustakos, T.; Papanastasiou, I.P. An overview on target-based drug design against kinetoplastid protozoan infections: Human African trypanosomiasis, Chagas disease and leishmaniases. Molecules 2021, 26, 4629. [Google Scholar] [CrossRef]
- Abdeta, D.; Kebede, N.; Giday, M.; Terefe, G.; Abay, S.M. In Vitro and In Vivo Antitrypanosomal Activities of Methanol Extract of Echinops kebericho Roots. Evid. Based Complementary Altern. Med. eCAM 2020, 2020, 8146756. [Google Scholar] [CrossRef]
- Fernández-Villa, D.; Aguilar, M.R.; Rojo, L. Folic acid antagonists: Antimicrobial and immunomodulating mechanisms and applications. Int. J. Mol. Sci. 2019, 20, 4996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scocchera, E.; Reeve, S.M.; Keshipeddy, S.; Lombardo, M.N.; Hajian, B.; Sochia, A.E.; Alverson, J.B.; Priestley, N.D.; Anderson, A.C.; Wright, D.L. Charged nonclassical antifolates with activity against gram-positive and gram-negative pathogens. ACS Med. Chem. Lett. 2016, 7, 692–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Paula Reis, M.; de Lima, D.A.; Pauli, K.B.; Andreotti, C.E.L.; de Moraes, A.L.S.; Gonçalves, D.D.; Navarro, I.T.; Bueno, P.S.A.; Seixas, F.A.V.; Gasparotto Junior, A.; et al. Molecular docking to Toxoplasma gondii thymidylate synthase-dihydrofolate reductase and efficacy of raltitrexed in infected mice. Parasitol. Res. 2018, 117, 1465–1471. [Google Scholar] [CrossRef]
- Gangjee, A.; Li, W.; Yang, J.; Kisliuk, R.L. Design, synthesis, and biological evaluation of classical and nonclassical 2-amino-4-oxo-5-substituted-6-methylpyrrolo [3, 2-d] pyrimidines as dual thymidylate synthase and dihydrofolate reductase inhibitors. J. Med. Chem. 2008, 51, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Hirumi, H.; Hirumi, K. Continuous cultivation of Trypanosoma brucei blood stream forms in a medium containing a low concentration of serum protein without feeder cell layers. J. Parasitol. 1989, 75, 985–989. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Fujioka, S.; Fukumoto, S.; Inoue, N.; Sakamoto, K.; Hirata, H.; Kido, Y.; Yabu, Y.; Suzuki, T.; Watanabe, Y.-i.; et al. Trypanosome alternative oxidase, a potential therapeutic target for sleeping sickness, is conserved among Trypanosoma brucei subspecies. Parasitol. Int. 2010, 59, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Suganuma, K.; Narantsatsral, S.; Battur, B.; Yamasaki, S.; Otgonsuren, D.; Musinguzi, S.P.; Davaasuren, B.; Battsetseg, B.; Inoue, N. Isolation, cultivation and molecular characterization of a new Trypanosoma equiperdum strain in Mongolia. Parasites Vectors 2016, 9, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, W. The origins of the trypanosome genome strains Trypanosoma brucei brucei TREU 927, T. b. gambiense DAL 972, T. vivax Y486 and T. congolense IL3000. Parasites Vectors 2012, 5, 71. [Google Scholar] [CrossRef] [Green Version]
- Suganuma, K.; Allamanda, P.; Hakimi, H.; Zhou, M.; Angeles, J.M.; Kawazu, S.; Inoue, N. Establishment of ATP-based luciferase viability assay in 96-well plate for Trypanosoma congolense. J. Vet. Med. Sci. 2014, 76, 1437–1441. [Google Scholar] [CrossRef] [Green Version]
- Burayk, S.; Oh-Hashi, K.; Kandeel, M. Drug Discovery of New Anti-Inflammatory Compounds by Targeting Cyclooxygenases. Pharmaceuticals 2022, 15, 282. [Google Scholar] [CrossRef]
- Kandeel, M.; Park, B.K.; Morsy, M.A.; Venugopala, K.N.; Oh-hashi, K.; Al-Nazawi, M.; Kwon, H.-J. Virtual Screening and Inhibition of Middle East Respiratory Syndrome Coronavirus Replication by Targeting Papain-like Protease. Dr. Sulaiman Al Habib Med. J. 2021, 3, 179–187. [Google Scholar] [CrossRef]
- Vanichtanankul, J.; Taweechai, S.; Yuvaniyama, J.; Vilaivan, T.; Chitnumsub, P.; Kamchonwongpaisan, S.; Yuthavong, Y. Trypanosomal dihydrofolate reductase reveals natural antifolate resistance. ACS Chem. Biol. 2011, 6, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Kandeel, M.; Kitade, Y.; Almubarak, A. Repurposing FDA-approved phytomedicines, natural products, antivirals and cell protectives against SARS-CoV-2 (COVID-19) RNA-dependent RNA polymerase. PeerJ 2020, 8, e10480. [Google Scholar] [CrossRef]
- Gibson, M.W.; Dewar, S.; Ong, H.B.; Sienkiewicz, N.; Fairlamb, A.H. Trypanosoma brucei DHFR-TS Revisited: Characterisation of a Bifunctional and Highly Unstable Recombinant Dihydrofolate Reductase-Thymidylate Synthase. PLoS Negl. Trop. Dis. 2016, 10, e0004714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullia, G.; Tamborini, L.; Conti, P.; De Micheli, C.; Pinto, A. Folates in Trypanosoma brucei: Achievements and opportunities. ChemMedChem 2018, 13, 2150–2158. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.F.; Mobashery, S.; Miller, M.J. Antibacterials; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Palomino, J.C.; Martin, A. The potential role of trimethoprim-sulfamethoxazole in the treatment of drug-resistant tuberculosis. Future Microbiol. 2016, 11, 539–547. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trypanosome | Raltitrexed | |
|---|---|---|
| Inhibition Rate at 25 µg/mL * | Inhibition Rate at 0.25 µg/mL * | |
| TcIL3000 | 1.77 ± 2.5 | 0 |
| TbbGUTat3.1 | 99.47 ± 0.07 | 0 |
| TbrIL1501 | 99.68 ± 0.16 | 1.01 ± 1.4 |
| TbgIL1922 | 99.6 ± 0.33 | 0 |
| Tev Tansui | 99.9 ± 0.12 | 0 |
| Teq IVM-t1 | 99.8 ± 0.027 | 0 |
| Raltitrexed | |
|---|---|
| Trypanosome | IC50 (µg/mL) * |
| TcIL3000 | >25 |
| TbbGUTat3.1 | 24.13 ± 0.01 |
| TbrIL1501 | 12.46 ± 0.05 |
| TbgIL1922 | 23.98 ± 0.14 |
| Tev Tansui | 11.57 ± 0.24 |
| Teq IVM-t1 | 5.18 ± 0.53 |
| Title | Docking Score | Glide Ligand Efficiency | Glide Hbond | Glide Lipo |
|---|---|---|---|---|
| Raltitrexed | −7.78 | −0.24 | −0.3 | −3.1 |
| WR99210 | −4.6 | −0.19 | −0.1 | −1.7 |
| Title | Docking Score | Glide Ligand Efficiency | Glide Hbond | Glide Lipo |
|---|---|---|---|---|
| T. brucei | −7.78 | −0.24 | −0.3 | −3.1 |
| Human | −7.2 | −0.22 | −0.32 | −2.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kandeel, M.; Suganuma, K. The Broad-Spectrum Antitrypanosomal Inhibitory Efficiency of the Antimetabolite/Anticancer Drug Raltitrexed. Processes 2022, 10, 2158. https://doi.org/10.3390/pr10112158
Kandeel M, Suganuma K. The Broad-Spectrum Antitrypanosomal Inhibitory Efficiency of the Antimetabolite/Anticancer Drug Raltitrexed. Processes. 2022; 10(11):2158. https://doi.org/10.3390/pr10112158
Chicago/Turabian StyleKandeel, Mahmoud, and Keisuke Suganuma. 2022. "The Broad-Spectrum Antitrypanosomal Inhibitory Efficiency of the Antimetabolite/Anticancer Drug Raltitrexed" Processes 10, no. 11: 2158. https://doi.org/10.3390/pr10112158