
Effects of Polyethylene Microplastics and Phenanthrene on Soil Properties, Enzyme Activities and Bacterial Communities

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Soil Incubation Experiment
2.3. Soil Chemical Properties and Enzymatic Activities Analysis
2.4. Soil DNA Extraction and 16S Amplicon Sequencing
2.5. Analysis of Microbial Community Diversity, Composition and Function
3. Results
3.1. Soil Chemical Properties
3.2. Soil Enzymatic Activities
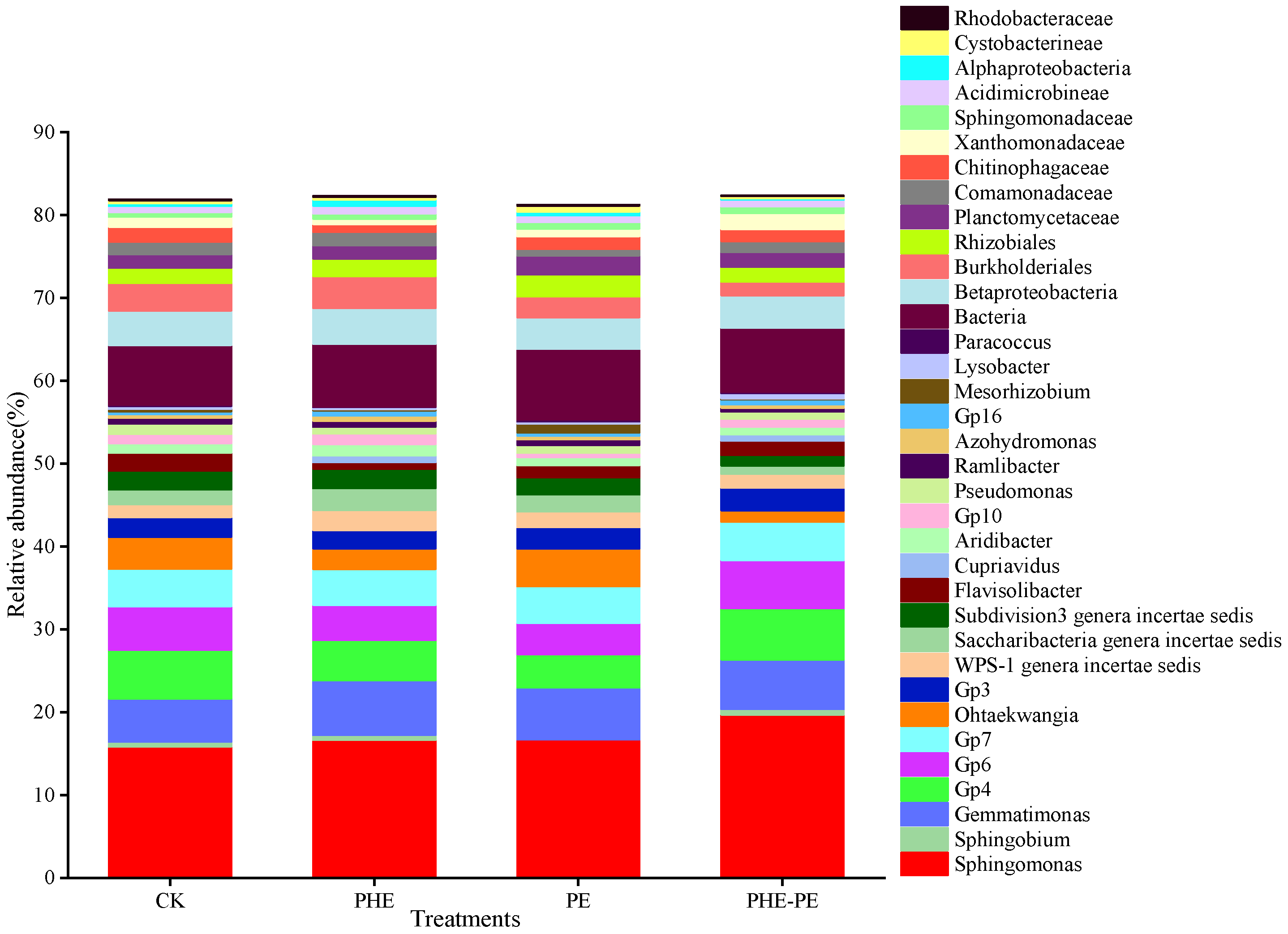
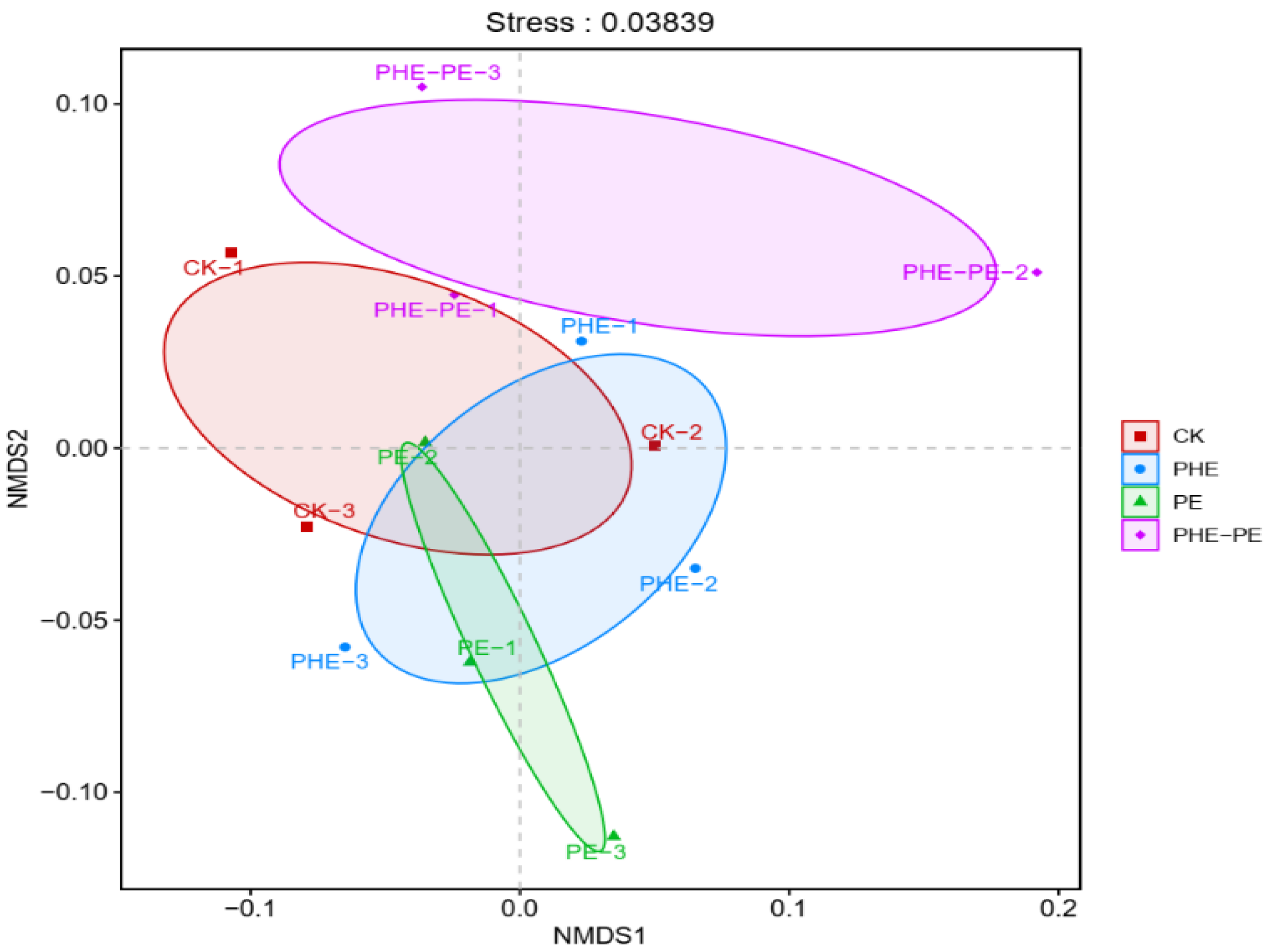
3.3. Microbial Community Diversity and Composition
3.4. Functional Prediction of Soil Microbial Communities
3.4.1. BugBase
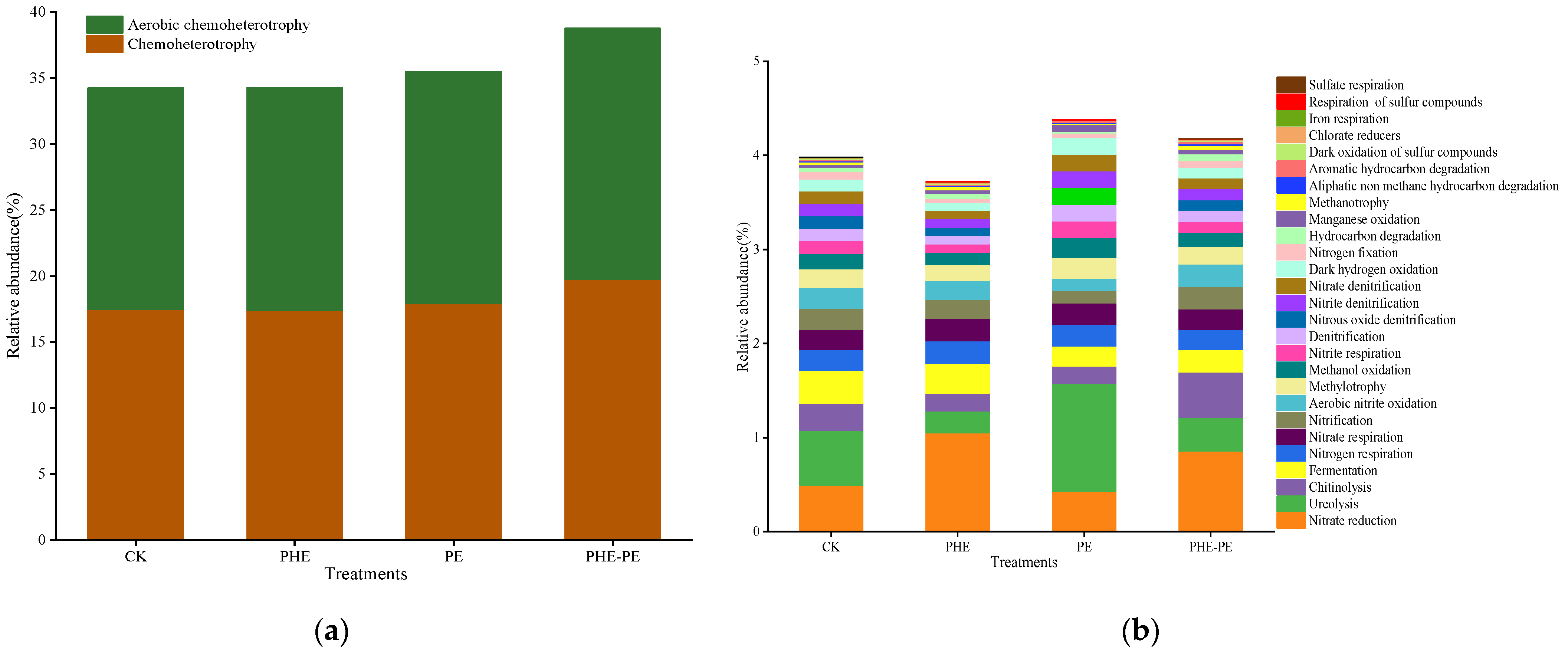
3.4.2. FAPROTAX
3.4.3. KEGG
4. Discussion
4.1. Effects of PHE and PE on Soil Chemical Properties
4.2. Effects of PHE and PE on Enzymatic Activities
4.3. Effects of PHE and PE on Microbial Diversity and Community Composition
4.4. Effects of PHE and PE on Soil Microbial Community Function
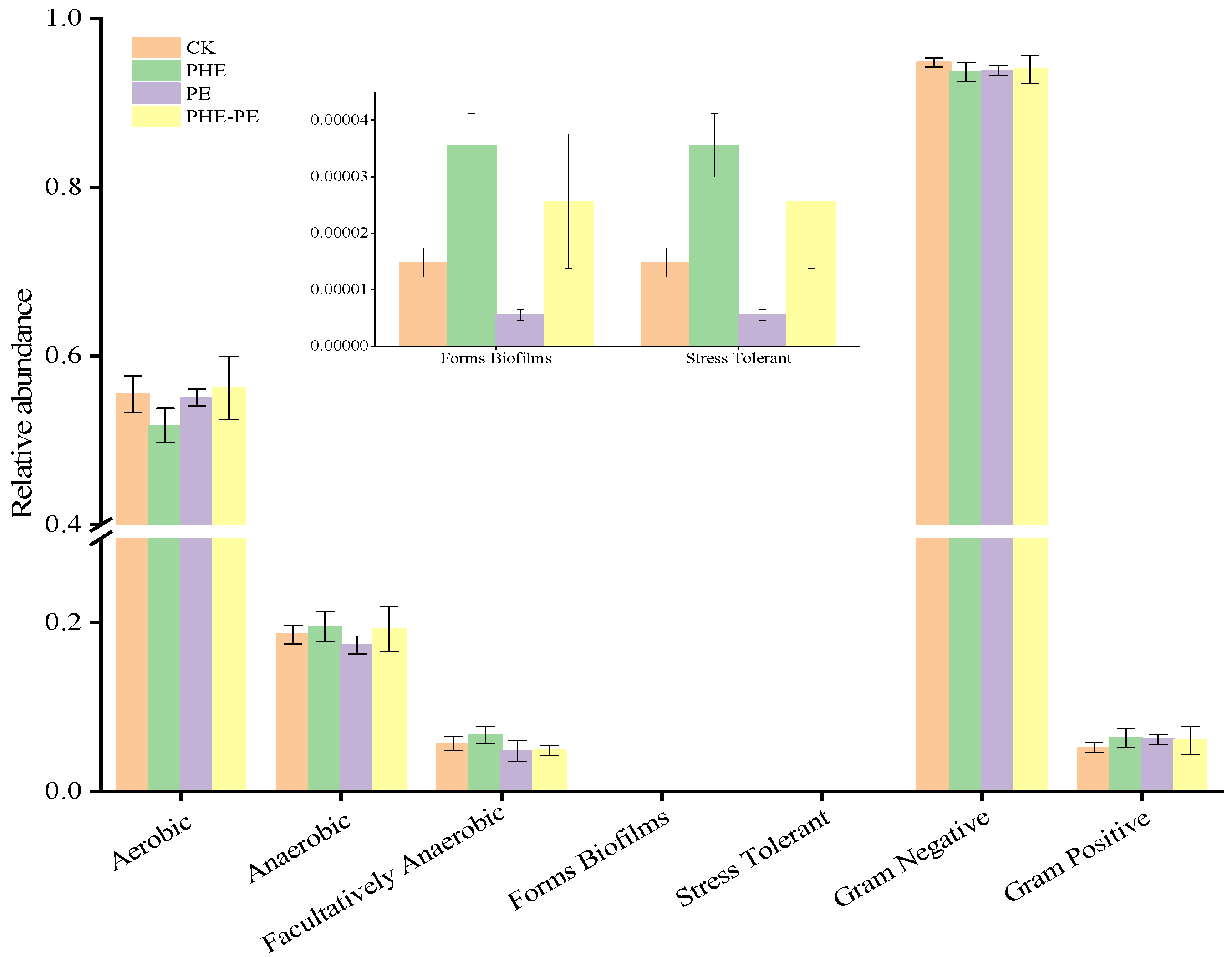
4.4.1. Microbial Phenotype Prediction Based on BugBase
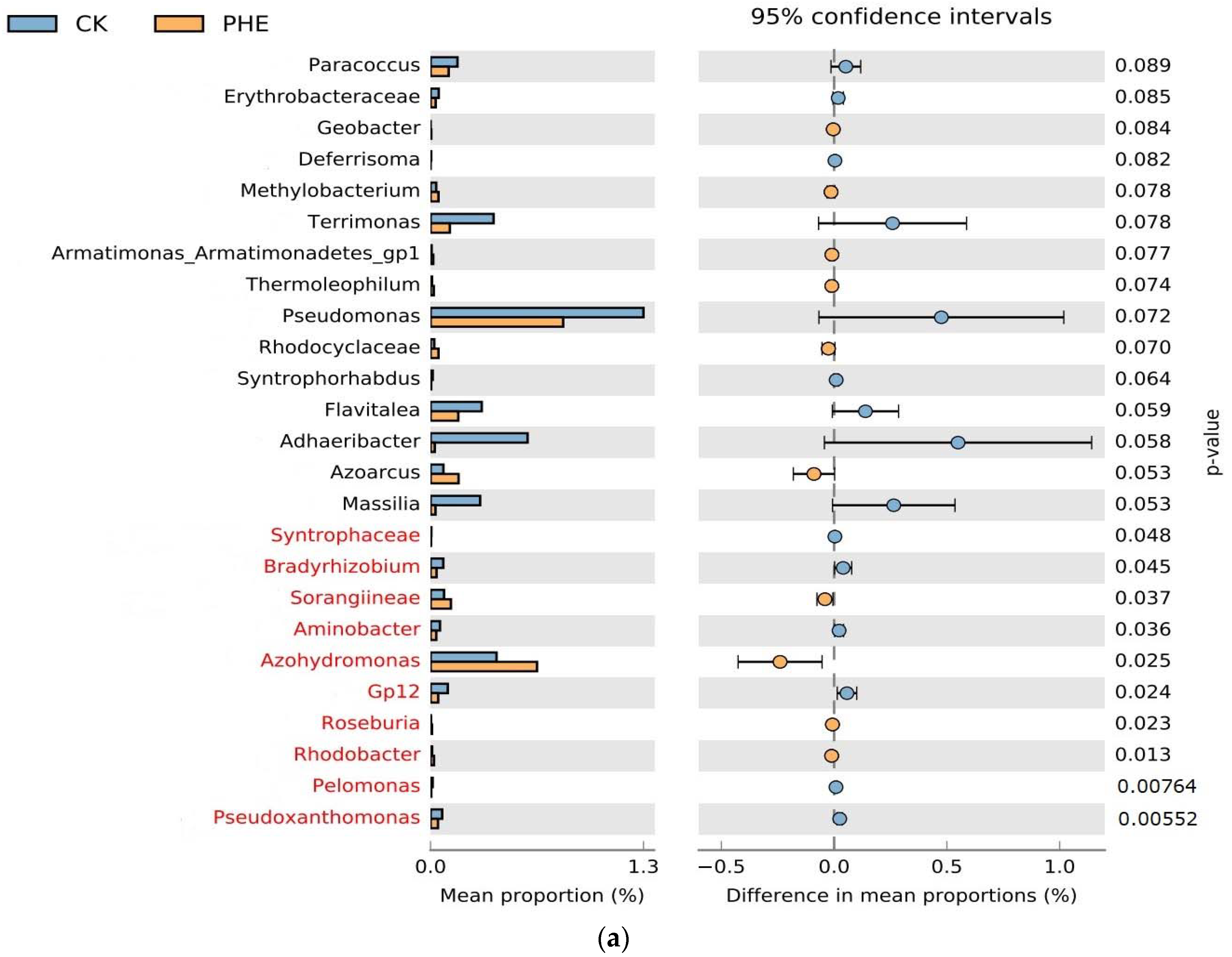
4.4.2. Functional Prediction of Bacterial Community by FAPROTAX
4.4.3. Predictive Metabolic Pathways Using KEGG
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Isobe, A.; Uchiyama–Matsumoto, K.; Uchida, K.; Tokai, T. Microplastics in the Southern Ocean. Mar. Pollut. Bull. 2017, 114, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Napper, I.E.; Bakir, A.; Rowland, S.J.; Thompson, R.C. Characterisation, quantity and sorptive properties of microplastics extracted from cosmetics. Mar. Pollut. Bull. 2015, 99, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Karbalaei, S.; Hanachi, P.; Walker, T.R.; Cole, M. Occurrence, sources, human health impacts and mitigation of microplastic pollution. Environ. Sci. Pollut. Res. 2018, 25, 36046–36063. [Google Scholar] [CrossRef] [PubMed]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.; Zhou, W.; Lu, S.; Huang, W.; Yuan, Q.; Tian, M.; Lv, W.; He, D. Microplastic pollution in rice–fish co–culture system: A report of three farmland stations in Shanghai, China. Sci. Total Environ. 2019, 652, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Liu, Y. The distribution of microplastics in soil aggregate fractions in southwestern China. Sci. Total Environ. 2018, 642, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Trilla-Prieto, N.; Vila-Costa, M.; Casas, G.; Jiménez, B.; Dachs, J. Dissolved black carbon and semivolatile aromatic hydrocarbons in the ocean: Two entangled biogeochemical cycles? Environ. Sci. Technol. Lett. 2021, 8, 918–923. [Google Scholar] [CrossRef]
- Wang, J.; Liu, J.; Ling, W.; Huang, Q.; Gao, Y. Composite of PAH–degrading endophytic bacteria reduces contamination and health risks caused by PAHs in vegetables. Sci. Total Environ. 2017, 598, 471–478. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, X.; Ling, W.; Liu, R.; Liu, J.; Kang, F.; Gao, Y. Contamination and health risk assessment of PAHs in soils and crops in industrial areas of the Yangtze River Delta region, China. Chemosphere 2017, 168, 976–987. [Google Scholar] [CrossRef]
- Chen, J.; Wu, Q.; Li, S.; Ge, J.; Liang, C.; Qin, H.; Xu, Q.; Fuhrmann, J.J. Diversity and function of soil bacterial communities in response to long–term intensive management in a subtropical bamboo forest. Geoderma 2019, 354, 113894. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, P.; Chen, Y. Polycyclic aromatic hydrocarbons contamination in surface soil of China: A review. Sci. Total Environ. 2017, 605, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Huang, S.; Zhang, H.; Tong, Y.; Barceló, D. Response of soil enzyme activities and bacterial communities to the accumulation of microplastics in an acid cropped soil. Sci. Total Environ. 2020, 707, 135634. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, L.; Rogers, E.; Altin, D.; Salaberria, I.; Booth, A.M. Sorption of PAHs to microplastic and their bioavailability and toxicity to marine copepods under co–exposure conditions. Environ. Pollut. 2020, 258, 113844. [Google Scholar] [CrossRef] [PubMed]
- Delgado–Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Su, Y.; Zhu, Y.; Chen, M.; Chen, B.; Liu, Y. Influences of polycyclic aromatic hydrocarbons (PAHs) on soil microbial community composition with or without Vegetation. J. Environ. Sci. Health Part A 2007, 42, 65–72. [Google Scholar] [CrossRef]
- Sutton, N.B.; Maphosa, F.; Morillo, J.A.; Al–Soud, W.A.; Langenhoff, A.A.M.; Grotenhuis, T.; Rijnaarts, H.H.M.; Smidt, H. Impact of long–term diesel contamination on soil microbial community structure. Appl. Environ. Microb. 2012, 79, 619–630. [Google Scholar] [CrossRef]
- Picariello, E.; Baldantoni, D.; De Nicola, F. Acute effects of PAH contamination on microbial community of different forest soils. Environ. Pollut. 2020, 262, 114378. [Google Scholar] [CrossRef]
- Picariello, E.; Baldantoni, D.; Muniategui-Lorenzo, S.; Concha-Grana, E.; De Nicola, F. A synthetic quality index to evaluate the functional stability of soil microbial communities after perturbations. Ecol. Indic. 2021, 128, 107844. [Google Scholar] [CrossRef]
- Picariello, E.; Baldantoni, D.; De Nicola, F. Investigating natural attenuation of PAHs by soil microbial communities: Insights by a machine learning approach. Restor. Ecol. 2022, 30, e13655. [Google Scholar] [CrossRef]
- Lee, D.W.; Lee, H.; Lee, A.H.; Kwon, B.; Khim, J.S.; Yim, U.H.; Kim, B.S.; Kima, J.J. Microbial community composition and PAHs removal potential of indigenous bacteria in oil contaminated sediment of Taean coast, Korea. Environ. Pollut. 2018, 234, 503–512. [Google Scholar] [CrossRef]
- Liu, H.; Yang, X.; Liu, G.; Liang, C.; Xue, S.; Chen, H.; Ritsema, C.J.; Geissen, V. Response of soil dissolved organic matter to microplastic addition in Chinese loess soil. Chemosphere 2017, 185, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Rillig, M.C. Microplastic disguising as soil carbon storage. Environ. Sci. Technol. 2018, 52, 6079–6080. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Yao, H.; Li, Y.; Zhu, Y. Microplastic addition alters the microbial community structure and stimulates soil carbon dioxide emissions in vegetable-growing soil. Environ. Toxicol. Chem. 2021, 40, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Ossowicki, A.; Yang, X.; Lwanga, E.H.; Dini–Andreote, F.; Geissen, V.; Garbeva, P. Effects of plastic mulch film residues on wheat rhizosphere and soil properties. J. Hazard. Mater. 2020, 387, 121711. [Google Scholar] [CrossRef]
- Ren, X.; Tang, J.; Liu, X.; Liu, Q. Effects of microplastics on greenhouse gas emissions and the microbial community in fertilized soil. Environ. Pollut. 2020, 256, 113347. [Google Scholar] [CrossRef]
- Lu, R. Soil and Agricultural Chemistry Analysis Methods; China Agricultural Science and Technology Press: Beijing, China, 2000; pp. 146–187. (In Chinese) [Google Scholar]
- Yi, M.; Zhou, S.; Zhang, L.; Ding, S. The effects of three different microplastics on enzyme activities and microbial communities in soil, Water. Environ. Res. 2020, 93, 24–32. [Google Scholar]
- Wallenstein, M.D.; Burns, R.G. Ecology of extracellular enzyme activities and organic matter degradation in soil: A complex community-driven process. In Methods of Soil Enzymology; Dick, R.P., Ed.; Soil Science Society of America: Inc.: Madison, WI, USA, 2011; Volume 9, pp. 35–55. [Google Scholar]
- Zeng, F.; Ali, S.; Zhang, H.; Ouyang, Y.; Qiu, B.; Wu, F.; Zhang, G.P. The influence of pH and organic matter content in paddy soil on heavy metal availability and their uptake by rice plants. Environ. Pollut. 2011, 159, 84–91. [Google Scholar] [CrossRef]
- Boots, B.; Russell, C.W.; Green, D.S. Effects of microplastics in soil ecosystems: Above and below ground. Environ. Sci. Technol. 2019, 53, 11496–11506. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Q.; Adams, C.A.; Sun, Y.; Zhang, S. Effects of microplastics on soil properties: Current knowledge and future perspectives. J. Hazard. Mater. 2022, 424, 127531. [Google Scholar] [CrossRef]
- Wang, Q.; Feng, X.; Liu, Y.; Cui, W.; Sun, Y.; Zhang, S.; Wang, F. Effects of microplastics and carbon nanotubes on soil geochemical properties and bacterial communities. J. Hazard. Mater. 2022, 433, 128826. [Google Scholar] [CrossRef]
- Xiao, M.; Ding, J.; Luo, Y.; Zhang, H.; Yu, Y.; Yao, H.; Zhu, Z.; Chadwick, D.R.; Jones, D.; Chen, J. Microplastics shape microbial communities affecting soil organic matter decomposition in paddy soil. J. Hazard. Mater. 2022, 431, 128589. [Google Scholar] [CrossRef] [PubMed]
- Coche, A.; Babey, T.; Rapaport, A.; Gonod, L.V.; Garnier, P.; Nunan, N.; de Dreuzy, J.R. Competition within low–density bacterial populations as an unexpected factor regulating carbon decomposition in bulk soil. Soil Biol. Biochem. 2022, 164, 108423. [Google Scholar] [CrossRef]
- Xiao, M.; Shahbaz, M.; Liang, Y.; Yang, J.; Wang, S.; Chadwicka, D.R.; Jones, D.; Chen, J.; Ge, T. Effect of microplastics on organic matter decomposition in paddy soil amended with crop residues and labile C: A three–source–partitioning study. J. Hazard. Mater. 2021, 416, 126221. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Gui, H.; Banfield, C.C.; Wen, Y.; Zang, H.; Dippold, M.A.; Charlton, A.; Jones, D.L. The microplastisphere: Biodegradable microplastics addition alters soil microbial community structure and function. Soil Biol. Biochem. 2021, 156, 108211. [Google Scholar] [CrossRef]
- Kielak, A.M.; Barreto, C.C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E.E. The ecology of acidobacteria: Moving beyond genes and genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef]
- He, T.; Li, Z.; Xie, D.; Sun, Q.; Xu, Y.; Ye, Q. Simultaneous nitrification and denitrification with different mixed nitrogen loads by a hypothermia aerobic bacterium. Biodegradation 2018, 29, 159–170. [Google Scholar] [CrossRef]
- Dong, Y.; Gao, M.; Qiu, W.; Song, Z.G. Effect of microplastics and arsenic on nutrients and microorganisms in rice rhizosphere soil. Ecotoxicol. Environ. Saf. 2021, 211, 111899. [Google Scholar] [CrossRef]
- Satyaprakash, M.; Nikitha, T.; Reddi, E.; Sadhana, B.; Vani, S.S. Phosphorous and phosphate solubilising bacteria and their role in plant nutrition. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2133–2144. [Google Scholar]
- Qu, Q.; Zhang, Z.; Peijnenburg, W.; Liu, W.; Lu, T.; Hu, B.; Chen, J.; Chen, J.; Lin, Z.; Qian, H. Rhizosphere microbiome assembly and its impact on plant growth. J. Agric. Food Chem. 2020, 68, 5024–5038. [Google Scholar] [CrossRef]
- Bergkemper, F.; Schöler, A.; Engel, M.; Lang, F.; Krüger, J.; Schloter, M. Phosphorus depletion in forest soils shapes bacterial communities towards phosphorus recycling systems. Environ. Microbiol. 2016, 18, 2767. [Google Scholar] [CrossRef]
- Yan, Y.; Chen, Z.; Zhu, F.; Zhu, C.; Wang, C.; Gu, C. Effect of polyvinyl chloride microplastics on bacterial community and nutrient status in two agricultural soils. Bull. Environ. Contam. Toxicol. 2020, 107, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhao, Y.; Wang, J.; Zhang, M.; Jia, W.; Qin, X. LDPE microplastic films alter microbial community composition and enzymatic activities in soil. Environ. Pollut. 2019, 254, 112983. [Google Scholar] [CrossRef] [PubMed]
- Kompaa–Bba, A.; Bierza, W.; Sierka, E.; Błońska, A.; Besenyei, L.; Woźniak, G. The role of plants and soil properties in the enzyme activities of substrates on hard coal mine spoil heaps. Sci. Rep. 2021, 11, 5155. [Google Scholar] [CrossRef] [PubMed]
- Perucci, P. Enzyme–activity and microbial biomass in a field soil amended with municipal refuse. Biol. Fertil. Soils 1992, 14, 54–60. [Google Scholar] [CrossRef]
- Zhang, F.; Li, M.; Qi, J.; Li, F.; Sun, G. Plastic film mulching increases soil respiration in ridge–furrow maize management. Arid Land Res. Manag. 2015, 29, 432–453. [Google Scholar] [CrossRef]
- Rostami, S.; Azhdarpoor, A.; Baghapour, M.A.; Dehghani, M.; Samaei, M.R.; Jaskulak, M.; Jafarpour, S.; Samare–Najaf, M. The effects of exogenous application of melatonin on the degradation of polycyclic aromatic hydrocarbons in the rhizosphere of Festuca. Environ. Pollut. 2021, 274, 116559. [Google Scholar] [CrossRef]
- Peng, D.; Ping, W.; Zhang, J.; Cui, M.; Liu, H. Damage of anodic biofilms by high salinity deteriorates PAHs degradation in single–chamber microbial electrolysis cell reactor. Sci. Total. Environ. 2021, 777, 145752. [Google Scholar]
- Condron, L.M.; Turner, B.L.; Cade–Menun, B.J.; Sims, J.; Sharpley, A. Chemistry and dynamics of soil organic phosphorus. Talnta 2005, 66, 87–121. [Google Scholar]
- Piotrowska–Dlugosz, A.; Charzynski, P. The impact of the soil sealing degree on microbial biomass, enzymatic activity, and physicochemical properties in the Ekranic Technosols of Toruń (Poland). J. Soil. Sediment. 2015, 15, 47–59. [Google Scholar] [CrossRef]
- Sun, Y.; Duan, C.; Cao, N.; Li, X.; Li, X.; Chen, Y.; Huang, Y.; Wang, J. Effects of microplastics on soil microbiome: The impacts of polymer type, shape, and concentration. Sci. Total Environ. 2022, 806, 150516. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Chen, Y.; Li, C.; Zhou, Z. Microbial community structure and co–occurrence are essential for methanogenesis and its contribution to phenanthrene degradation in paddy soil. J. Hazard. Mater. 2021, 417, 126086. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Luo, C.; Zhang, D.; Song, M.; Cai, X.; Jiang, L.; Zhang, G. Autochthonous bioaugmentation modified bacterial diversity of phenanthrene degraders in PAH–contaminated wastewater as revealed by DNA–stable isotope probing. Environ. Sci. Technol. 2018, 52, 2934–2944. [Google Scholar] [CrossRef]
- Judy, J.D.; Williams, M.; Gregg, A.; Oliver, D.; Kumar, A.; Kookana, R.; Kirbya, J.K. Microplastics in municipal mixed–waste organic outputs induce minimal short to long–term toxicity in key terrestrial biota. Environ. Pollut. 2019, 252, 522–531. [Google Scholar] [CrossRef]
- Xu, M.; Du, W.; Ai, F.; Xu, F.; Zhu, J.G.; Yin, Y.; Ji, R.; Guo, H. Polystyrene microplastics alleviate the effects of sulfamethazine on soil microbial communities at different CO2 concentrations. J. Hazard. Mater. 2021, 413, 125286. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Liang, S.; Yang, X. Isolation and characterization of catechol 2,3–dioxygenase genes from phenanthrene degraders Sphingomonas sp. ZP1 and Pseudomonas sp. ZP2. Environ. Technol. 2011, 32, 1895–1901. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Huang, Y.; Rene, E.R.; Kumar, A.J.; Chen, S. Mechanism of allethrin biodegradation by a newly isolated Sphingomonas trueperi strain CW3 from wastewater sludge. Bioresour. Technol. 2020, 305, 123074. [Google Scholar]
- Kalam, S.; Basu, A.; Ahmad, I.; Kumar, A.J.; Chen, S. Recent understanding of soil acidobacteria and their ecological significance: A critical review. Front. Microbiol. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Liu, M.; Lu, S.; Song, Y.; Lei, L.; Hu, J.; Lv, W.; Zhou, W.; Cao, C.; Shi, H.; Yang, X.; et al. Microplastic and mesoplastic pollution in farmland soils in suburbs of Shanghai, China. Environ. Pollut. 2018, 242, 855–862. [Google Scholar] [CrossRef]
- Dahal, R.H.; Chaudhary, D.K.; Kim, D.U.; Kim, J. Azohydromonas caseinilytica sp. nov. a nitrogen–fixing bacterium isolated from forest soil by using optimized culture method. Front. Microbiol. 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Flores–Núez, V.M.; Fonseca, C.; Desgarennes, D.; Eloe–Fadrosh, E.; Woyke, T.; Partida–Martínez, L.P. Functional signatures of the epiphytic prokaryotic microbiome of agaves and cacti. Front. Microbiol. 2020, 10, 3044. [Google Scholar] [CrossRef]
- Negi, S.; Kumar, P.; Kumar, J.; Singh, A.; Dubey, R.C. Indigenous nitrogen fixing microbes engineer rhizosphere and enhance nutrient availability and plant growth. In Rhizosphere Engineering; Dubey, R.C., Pankaj, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; Volume 21, pp. 19–43. [Google Scholar]
- Wang, W.; Wang, N.; Dang, K.; Dai, W.; Guan, L.; Wang, B.; Gao, J.; Cui, Z.; Dong, Y.; Wang, H. Long–term nitrogen application decreases the abundance and copy number of predatory myxobacteria and alters the myxobacterial community structure in the soil. Sci. Total Environ. 2019, 708, 135114. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.; Pistorius, D.; Stadler, M.; Müller, R. Fatty acid–Related phylogeny of myxobacteria as an approach to discover polyunsaturated omega–3/6 fatty acids. J. Bacteriol. 2011, 193, 1930–1942. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Chen, K.; Han, D.; Zhao, J.; Lu, Y.; Yang, G.F.; Mu, J.; Zhao, X. Comparison of nitrogen removal and microbial properties in solid–phase denitrification systems for water purification with various pretreated lignocellulosic carriers. Bioresour. Technol. 2017, 224, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Sur, I.M.; Micle, V.; Polyak, E.T.; Gabor, T. Assessment of soil quality status and the ecological risk in the Baia Mare, Romania Area. Sustainability 2022, 14, 3739. [Google Scholar] [CrossRef]
- Sur, I.M.; Micle, V.; Hegyi, A.; Lăzărescu, A.V. Extraction of metals from polluted soils by bioleaching in relation to environmental risk assessment. Materials 2022, 15, 3973. [Google Scholar] [CrossRef]
- de Souza Machado, A.A.; Lau, C.W.; Till, J.; Kloas, W.; Lehmann, A.; Becker, R.; Rillig, M.C. Impacts of microplastics on the soil biophysical environment. Environ. Sci. Technol. 2018, 52, 9656–9665. [Google Scholar] [CrossRef]
- Qi, Y.; Beriot, N.; Gort, G.; Huerta Lwanga, E.; Gooren, H.; Yang, X.; Geissen, V. Impact of plastic mulch film debris on soil physicochemical and hydrological properties. Environ. Pollut. 2020, 266, 115097. [Google Scholar] [CrossRef]
- Akhavan, O.; Ghaderi, E. Toxicity of graphene and graphene oxide nanowalls against bacteria. ACS Nano 2010, 4, 5731–5736. [Google Scholar] [CrossRef]
- Liang, S.; Deng, J.; Jiang, Y.; Wu, S. Functional distribution of bacterial community under different land use patterns based on FaProTax function prediction. Pol. J. Environ. Stud. 2020, 29, 1245–1261. [Google Scholar] [CrossRef]
- Hubert, L.; Sutton, V.R. Disorders of purine and pyrimidine metabolism. In Biomarkers in Inborn Errors of Metabolism; Garg, U., Smith, L.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 283–299. [Google Scholar]
- Rahmawati, S.I.; Izzati, F.N.; Hapsari, Y.; Septiana, E.; Rachman, F.; Bustanussalam, B.; Simanjuntak, P. Endophytic microbes and antioxidant activities of secondary metabolites from mangroves Avicennia marina and Xylocarpus granatum. IOP Conf. Ser. Earth Environ. Sci. 2019, 278, 012065. [Google Scholar] [CrossRef]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell. Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D. A nearest neighbor approach for automated transporter prediction and categorization from protein sequences. Bioinformatics 2008, 24, 1129–1136. [Google Scholar]
- Hessel, S.; John, A.; Seidel, A.; Lampen, A. Multidrug resistance–associated proteins are involved in the transport of the glutathione conjugates of the ultimate carcinogen of benzo[a]pyrene in human Caco–2 cells. Arch. Toxicol. 2013, 87, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Pal, D. Clusters of hairpins induce intrinsic transcription termination in bacteria. Sci. Rep. 2021, 11, 16194. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, L.; Hou, J.; Wang, X.; Liu, H.; Zheng, D.; Liang, R. iTRAQ–based quantitative proteomic analysis of the global response to 17β–estradiol in estrogen–degradation strain Pseudomonas putida SJTE–1. Sci. Rep. 2017, 7, 41682. [Google Scholar] [CrossRef]
- Piattelli, E.; Peltier, J.; Soutourina, O. Interplay between regulatory RNAs and signal transduction systems during bacterial infection. Genes 2020, 11, 1209. [Google Scholar] [CrossRef]
- Kisand, V.; Valente, A.; Lahm, A.; Tanet, G.; Lettieri, T. Phylogenetic and functional metagenomic profiling for assessing microbial biodiversity in environmental monitoring. PLoS ONE 2012, 7, e43630. [Google Scholar] [CrossRef]
- Sur, I.M.; Micle, V.; Damian, G.E. Assessment of heavy metal contamination and bioremediation potential of Thiobacillus ferrooxidans in soils around copper quarry. J. Environ. Prot. Ecol. 2020, 21, 56–62. [Google Scholar]
- Micle, V.; Sur, I.M. Experimental investigation of a pilot-scale concerning ex-situ bioremediation of petroleum hydrocarbons contaminated soils. Sustainability 2021, 13, 8165. [Google Scholar] [CrossRef]
- Evans, M.J.; Cravatt, B.F. Mechanism–based profiling of enzyme families. Chem. Rev. 2006, 106, 3279–3301. [Google Scholar] [CrossRef]
- Kandlinger, F.; Plach, M.G.; Merkl, R. AGeNNT: Annotation of enzyme families by means of refined neighborhood networks. BMC Bioinform. 2017, 18, 274. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Chemical Properties | CK | PHE | PE | PHE- PE |
|---|---|---|---|---|
| pH | 7.427 ± 0.012 | 7.440 ± 0.017 | 7.270 ± 0.010 * | 7.320 ± 0.036 * |
| SOM (g kg−1) | 9.189 ± 0.439 | 9.358 ± 0.818 | 8.085 ± 0.147 * | 9.869 ± 0.365 |
| AN (mg kg−1) | 37.157 ± 0.001 | 34.906 ± 3.906 | 30.402 ± 0.002 * | 32.652 ± 3.894 |
| AP (mg kg−1) | 2.118 ± 0.066 | 2.018 ± 0.152 | 1.986 ± 0.095 | 2.024 ± 0.178 |
| Soil Enzymatic Activities | CK | PHE | PE | PHE- PE |
|---|---|---|---|---|
| Urease (mg NH3-N g−1 soil) | 1.949 ± 0.179 | 1.712 ± 0.098 | 2.700 ± 0.302 * | 1.918 ± 0.289 |
| FDAse (μg FDA g−1 soil) | 7.119 ± 1.351 | 7.224 ± 0.685 | 9.630 ± 0.293 * | 8.555 ± 3.871 |
| Dehydrogenase (μL H+ 20 g−1 soil) | 3.413 ± 0.451 | 3.613 ± 0.230 | 3.463 ± 0.087 | 5.618 ± 1.206 * |
| Phosphatase (mg phenol g−1 soil) | 0.103 ± 0.006 | 0.089 ± 0.011 | 0.085 ± 0.016 | 0.089 ± 0.011 |
| Treatments | Shannon | Chao | Ace | Simpson |
|---|---|---|---|---|
| CK | 5.968 ± 0.076 | 2391.022 ± 116.673 | 2377.793 ± 88.873 | 0.008 ± 0.002 |
| PHE | 5.944 ± 0.071 | 2401.953 ± 120.764 | 2386.161 ± 91.593 | 0.010 ± 0.001 |
| PE | 6.017 ± 0.111 | 2378.052 ± 13.755 | 2363.966 ± 19.253 | 0.008 ± 0.001 |
| PHE-PE | 5.894 ± 0.206 | 2413.304 ± 44.935 | 2395.093 ± 35.803 | 0.011 ± 0.004 |
| Pathways | CK | PHE | PE | PHE-PE |
|---|---|---|---|---|
| Amino acid metabolism | 5,687,523 ± 1,106,040 | 6,008,889 ± 939,065 | 9,677,218 ± 1,179,844 * | 6,773,054 ± 862,218 |
| Carbohydrate metabolism | 5,327,248 ± 1,022,491 | 5,593,268 ± 830,559 | 8987628 ± 1,112,622 * | 6,320,786 ± 715,952 |
| Membrane transport | 4,823,873 ± 1,014,658 | 5,134,273 ± 814,056 | 8,333,681 ± 945,512 * | 5,651,892 ± 713,316 |
| Replication and repair | 3,808,074 ± 734,029 | 3,998,707 ± 625,335 | 6,377,432 ± 797,193 * | 4,478,773 ± 484,227 |
| Energy metabolism | 3,138,171 ± 627,139 | 3,325,013 ± 500,374 | 5,276,552 ± 642,955 * | 3,686,084 ± 370,698 |
| Lipid metabolism | 2,099,475 ± 402,180 | 2,219,049 ± 354,419 | 3,577,720 ± 445,975 * | 2,518,405 ± 332,718 |
| Translation | 2,377,536 ± 473,274 | 2,512,310 ± 383,629 | 3,992,523 ± 495,321 * | 2,783,287 ± 282,147 |
| Metabolism of cofactors and vitamins | 2,267,936 ± 443,783 | 2,392,752 ± 370,863 | 3,831,557 ± 473,485 * | 2,673,111 ± 314,202 |
| Cellular processes and signaling | 1,950,477 ± 363,800 | 2,031,076 ± 326,293 | 3,248,166 ± 388,559 * | 2,296,525 ± 278,658 |
| Xenobiotics biodegradation and metabolism | 1,800,127 ± 337,033 | 1,938,425 ± 331,984 | 3,105,698 ± 359,437 * | 2,240,873 ± 469,536 |
| Cell motility | 1,835,961 ± 312,990 | 1,957,392 ± 340,056 | 2,997,769 ± 368,876 * | 2,227,334 ± 296,725 |
| Nucleotide metabolism | 1,727,368 ± 342,220 | 1,816,148 ± 279,130 | 2,905,554 ± 355,955 * | 2,022,893 ± 209,472 |
| Genetic information processing | 1,282,896 ± 249,837 | 1,347,077 ± 206,965 | 2,175,475 ± 269,490 * | 1,504,842 ± 161,464 |
| Folding, sorting and degradation | 1,270,334 ± 247,722 | 1,339,382 ± 206,626 | 2,123,214 ± 265,497 * | 1,486,413 ± 155,618 |
| Transcription | 1,270,122 ± 236,126 | 1,327,185 ± 212,216 | 2,139,723 ± 277,642 * | 1,504,305 ± 158,066 |
| Glycan biosynthesis and metabolism | 1,214,360 ± 240,003 | 1,247,544 ± 193,799 | 1,997,518 ± 283,128 * | 1,393,183 ± 62,631 |
| Signal transduction | 1,208,537 ± 220,584 | 1,280,745 ± 221,183 | 1,998,523 ± 242,461 * | 1,435,260 ± 178,339 |
| Metabolism of terpenoids and polyketides | 1,119,529 ± 217,912 | 1,192,808 ± 189,649 | 1,909,573 ± 235,880 * | 1,340,870 ± 186,128 |
| Enzyme families | 1,082,715 ± 208,021 | 1,134,254 ± 192,093 | 1,813,880 ± 243,363 * | 1,283,329 ± 109,820 |
| Cell growth and death | 311,845 ± 62,262 | 332,013 ± 51,409 | 541,832 ± 58,802 * | 373,824 ± 53,605 |
| Transport and catabolism | 189,182 ± 34,903 | 195,612 ± 28,236 | 319,386 ± 42,214 * | 224,248 ± 28,276 |
| Signaling molecules and interaction | 103,635 ± 18,364 | 103,044 ± 16,871 | 168,531 ± 25,056 * | 1,22,624 ± 11,234 |
| Environmental adaptation | 85,022 ± 16,316 | 89,595 ± 13,882 | 140,888 ± 16,832 * | 99,957 ± 9358 |
| Immune system | 21,286 ± 5060 | 22,507 ± 4475 | 36,422 ± 4323 * | 23,761 ± 1891 |
| Excretory system | 20,762 ± 4345 | 21,552 ± 3309 | 35,427 ± 4640 * | 23,473 ± 1343 |
| Biosynthesis of secondary metabolites | 589,242 ± 109,571 | 611,122 ± 94,164 | 991,848 ± 133,240 * | 697,493 ± 54,578 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Huang, K.; Yuan, G.; Yang, C. Effects of Polyethylene Microplastics and Phenanthrene on Soil Properties, Enzyme Activities and Bacterial Communities. Processes 2022, 10, 2128. https://doi.org/10.3390/pr10102128
Liu S, Huang K, Yuan G, Yang C. Effects of Polyethylene Microplastics and Phenanthrene on Soil Properties, Enzyme Activities and Bacterial Communities. Processes. 2022; 10(10):2128. https://doi.org/10.3390/pr10102128
Chicago/Turabian StyleLiu, Shasha, Kaibo Huang, Guodong Yuan, and Chengfang Yang. 2022. "Effects of Polyethylene Microplastics and Phenanthrene on Soil Properties, Enzyme Activities and Bacterial Communities" Processes 10, no. 10: 2128. https://doi.org/10.3390/pr10102128