
Defective Leukocyte β2 Integrin Expression and Reactive Oxygen Species Production in Neonates

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Integrin Cell Surface Expression
2.3. Superoxide Radical Production
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wynn, J.L. Defining neonatal sepsis. Curr. Opin. Pediatr. 2016, 28, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Carr, R.; Huizinga, T.W. Low soluble FcRIII receptor demonstrates reduced neutrophil reserves in preterm neonates. Arch. Dis. Child. Fetal Neonatal Ed. 2000, 83, 160–164. [Google Scholar] [CrossRef] [Green Version]
- Källman, J.; Schollin, J.; Schalèn, C.; Erlandsson, A.; Kihlström, E. Impaired phagocytosis and opsonisation towards group B streptococci in preterm neonates. Arch. Dis. Child. Fetal Neonatal Ed. 1998, 78, 46–50. [Google Scholar] [CrossRef]
- Drossou, V.; Kanakoudi, F.; Tzimouli, V.; Sarafidis, K.; Taparkou, A.; Bougiouklis, D.; Petropoulou, T.; Kremenopoulos, G. Impact of prematurity, stress and sepsis on the neutrophil respiratory burst activity of neonates. Biol. Neonate 1997, 72, 201–209. [Google Scholar] [CrossRef]
- Badolato, R. Defects of leukocyte migration in primary immunodeficiencies. Eur. J. Immunol. 2013, 43, 1436–1440. [Google Scholar] [CrossRef]
- De Rose, D.U.; Giliani, S.; Notarangelo, L.D.; Lougaris, V.; Lanfranchi, A.; Moratto, D.; Martire, B.; Specchia, F.; Tommasini, A.; Plebani, A.; et al. Long term outcome of eight patients with type 1 Leukocyte Adhesion Deficiency (LAD-1): Not only infections, but high risk of autoimmune complications. Clin. Immunol. 2018, 191, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.M.; Babensee, J.E.; Simon, S.I.; Lu, H.; Perrard, J.L.; Bullard, D.C.; Dai, X.Y.; Bromley, S.K.; Dustin, M.L.; Entman, M.L.; et al. Relative contribution of LFA-1 and Mac-1 to neutrophil adhesion and migration. J. Immunol. 1999, 163, 5029–5038. [Google Scholar]
- Thornton, B.; Vĕtvicka, V.; Pitman, M.; Goldman, R.C.; Ross, G.D. Analysis of the sugar specificity and molecular location of the beta-glucan-binding lectin site of complement receptor type 3 (CD11b/CD18). J. Immunol. 1996, 156, 1235–1246. [Google Scholar]
- Babior, B.M. NADPH oxidase. Curr. Opin. Immunol. 2004, 16, 42–47. [Google Scholar] [CrossRef]
- Yan, J.; Vetvicka, V.; Xia, Y.; Hanikýrová, M.; Mayadas, T.N.; Ross, G.D. Critical role of Kupffer cell CR3 (CD11b/CD18) in the clearance of IgM-opsonized erythrocytes or soluble β-glucan. Immunopharmacology 2000, 46, 39–54. [Google Scholar] [CrossRef]
- Yan, J.; Vetvicka, V.; Xia, Y.; Coxon, A.; Carroll, M.C.; Mayadas, T.N.; Ross, G.D. B-glucan, a “specific” biologic response modifier that uses antibodies to target tumors for cytotoxic recognition by Leukocyte Complement Receptor Type 3 (CD11b/CD18). J. Immunol. 1999, 163, 3045–3052. [Google Scholar] [PubMed]
- Bannister, J.V.; Bellavite, P.; Davoli, A.; Thornalley, P.J.; Rossi, F. The generation of hydroxyl radicals following superoxide production by neutrophil NADPH oxidase. FEBS Lett. 1982, 150, 300–302. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.C.; Khodadadi, H.; Malik, A.; Davidson, B.; Salles, É.; Bhatia, J.; Hale, V.L.; Baban, B. Innate immunity of neonates and infants. Front. Immunol. 2018, 9, 1759. [Google Scholar] [CrossRef]
- Yektaei-Karin, E.; Moshfegh, A.; Lundahl, J.; Berggren, V.; Hansson, L.O.; Marchini, G. The stress of birth enhances in vitro spontaneous and IL-8-induced neutrophil chemotaxis in the human newborn. Pediatr. Allergy Immunol. 2007, 18, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Stålhammar, M.E.; Sindelar, R.; Douhan Håkansson, L. Neutrophil receptor response to bacterial n-formyl peptides is similar in term newborn infants and adults in contrast to IL-8. Scand. J. Immunol. 2016, 84, 332–337. [Google Scholar] [CrossRef]
- Yuki, K.; Hou, L. Role of B2 integrins in neutrophils and sepsis. Infect. Immun. 2020, 88, e00031-20. [Google Scholar] [CrossRef]
- O’Hare, F.M.; Watson, W.; O’Neill, A.; Grant, T.; Onwuneme, C.; Donoghue, V.; Mooney, E.; Downey, P.; Murphy, J.; Twomey, A.; et al. Neutrophil and monocyte toll-like receptor 4, CD11b and reactive oxygen intermediates, and neuroimaging outcomes in preterm infants. Pediatr. Res. 2015, 78, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.M.; Steadman, R.; Williams, J.D. Human neutrophils are selectively activated by independent ligation of the subunits of the CD11b/CD18 integrin. J. Leukoc. Biol. 1994, 56, 708–713. [Google Scholar] [CrossRef]
- Nixon, J.B.; McPhail, L.C. Protein kinase C (PKC) isoforms translocate to Triton-insoluble fractions in stimulated human neutrophils: Correlation of conventional PKC with activation of NADPH oxidase. J. Immunol. 1999, 163, 4574–4582. [Google Scholar]
- Cosentino-Gomes, D.; Rocco-Machado, N.; Meyer-Fernandes, J.R. Cell signaling through protein kinase C oxidation and activation. Int. J. Mol. Sci. 2012, 13, 10697–10721. [Google Scholar] [CrossRef] [Green Version]
- Le Cabec, V.; Carréno, S.; Moisand, A.; Bordier, C.; Maridonneau-Parini, I. Complement receptor 3 (CD11b/CD18) mediates type i and type ii phagocytosis during nonopsonic and opsonic phagocytosis, respectively. J. Immunol. 2002, 169, 2003–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, G.C.; Chan, W.K.; Sze, D.M. The effects of beta-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Ross, G.D. Generation of recombinant fragments of CD11b expressing the functional beta-glucan-binding lectin site of CR3 (CD11b/CD18). J. Immunol. 1999, 162, 7285–7293. [Google Scholar] [PubMed]
- Xia, Y.; Borland, G.; Huang, J.; Mizukami, I.F.; Petty, H.R.; Todd, R.F.; Ross, G.D. Function of the lectin domain of mac-1/complement receptor type 3 (CD11b/CD18) in regulating neutrophil adhesion. J. Immunol. 2002, 169, 6417–6426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geelen, S.P.; Fleer, A.; Bezemer, A.C.; Gerards, L.J.; Rijkers, G.T.; Verhoef, J. Deficiencies in opsonic defense to pneumococci in the human newborn despite adequate levels of complement and specific IgG antibodies. Pediatr. Res. 1990, 27, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Chudgar, U.H.; Thurman, G.W.; Ambruso, D.R. Oxidase activity in cord blood neutrophils: A balance between increased membrane associated cytochrome b558 and deficient cytosolic components. Pediatr. Blood Cancer 2005, 45, 311–317. [Google Scholar] [CrossRef]
- Osiak, W.; Wątroba, S.; Kapka-Skrzypczak, L.; Kurzepa, J. Two Faces of heme catabolic pathway in newborns: A potential role of bilirubin and carbon monoxide in neonatal inflammatory diseases. Oxid. Med. Cell. Longev. 2020, 2020, 7140496. [Google Scholar] [CrossRef]
- Nakasone, R.; Ashina, M.; Abe, S.; Tanimura, K.; Van Rostenberghe, H.; Fujioka, K. The role of heme oxygenase-1 promoter polymorphisms in perinatal disease. Int. J. Environ. Res. Public Health 2021, 18, 3520. [Google Scholar] [CrossRef]
- Hayashi, S.; Takamiya, R.; Yamaguchi, T.; Matsumoto, K.; Tojo, S.J.; Tamatani, T.; Kitajima, M.; Makino, N.; Ishimura, Y.; Suematsu, M. Induction of heme oxygenase-1 suppresses venular leukocyte adhesion elicited by oxidative stress: Role of bilirubin generated by the enzyme. Circ. Res. 1999, 85, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Andersson, J.A.; Egesten, A.; Cardell, L.O. Hemin, a heme oxygenase substrate analog, inhibits the cell surface expression of CD11b and CD66b on human neutrophils. Allergy 2002, 57, 718–722. [Google Scholar] [CrossRef]
- Morisaki, H.; Katayama, T.; Kotake, Y.; Ito, M.; Handa, M.; Ikeda, Y.; Takeda, J.; Suematsu, M. Carbon monoxide modulates endotoxin-induced microvascular leukocyte adhesion through platelet-dependent mechanisms. Anesthesiology 2002, 97, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Klimova, N.G.; Hanna, N.; Peltier, M.R. Does carbon monoxide inhibit proinflammatory cytokine production by fetal membranes? J. Perinat. Med. 2013, 41, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Li, J.; Yang, X.; Tang, J.; Shi, J.; Tong, Y.; Qu, Y.; Mu, D. Is neutrophil CD11b a special marker for the early diagnosis of sepsis in neonates? A systematic review and meta-analysis. BMJ Open 2019, 9, e025222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, H.E.; Ibrahim, Z.H.; Ahmed, W.O. Diagnostic, prognostic, predictive and monitoring role of neutrophil CD11b and monocyte CD14 in neonatal sepsis. Dis. Markers 2021, 2021, 4537760. [Google Scholar] [CrossRef]


{kind=link}
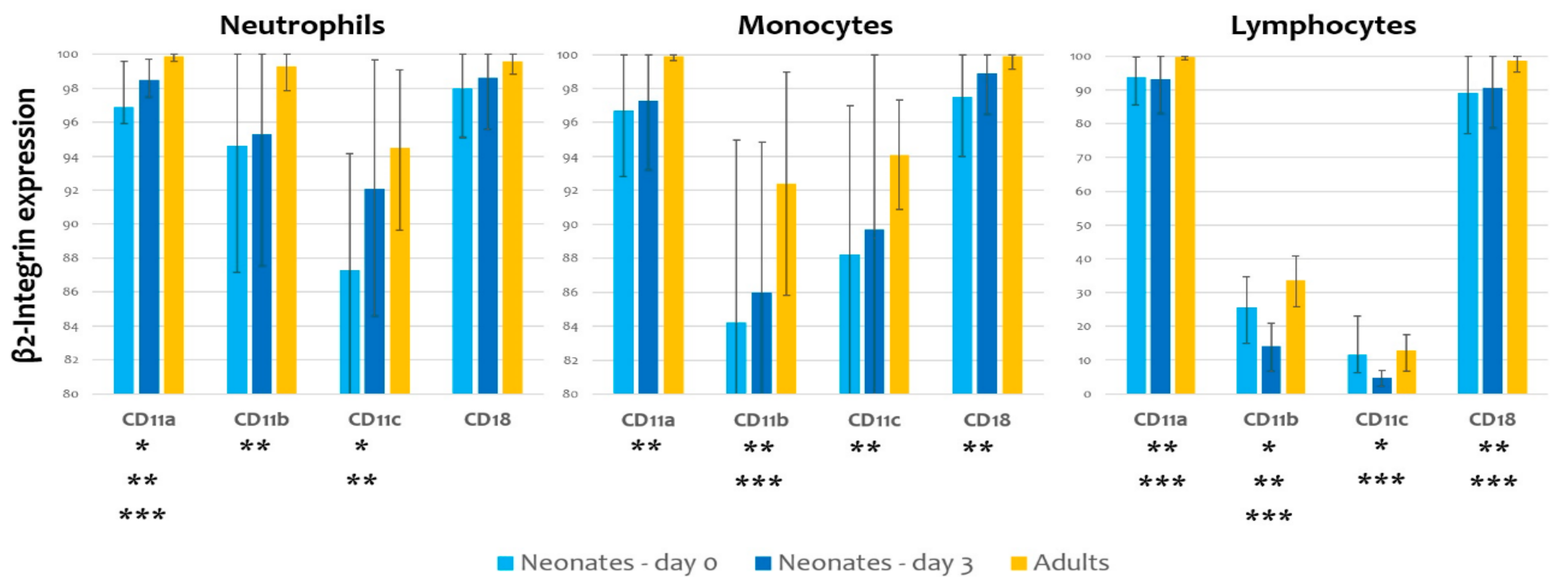
| Neonates—Day 0 | Neonates—Day 3 | Adults | p-Value (Neonates Day 0 vs. Day 3) | p-Value (Neonates Day 0 vs. Adults) | p-Value (Neonates Day 3 vs. Adults) | ||
|---|---|---|---|---|---|---|---|
| Neutrophils | CD11a | 96.9 ± 2.7 | 98.5 ± 1.3 | 99.9 ± 0.1 | <0.01 * | <0.01 * | <0.01 * |
| CD11b | 94.6 ± 7.5 | 95.3 ± 7.6 | 99.3 ± 1.4 | 0.70 | 0.04 * | 0.08 | |
| CD11c | 87.3 ± 7.0 | 92.1 ± 7.6 | 94.5 ± 4.7 | <0.01 * | <0.01 * | 0.31 | |
| CD18 | 98.0 ± 2.7 | 98.6 ± 2.9 | 99.6 ± 1.1 | 0.37 | 0.05 | 0.25 | |
| Monocytes | CD11a | 96.7 ± 4.0 | 97.3 ± 4.3 | 99.9 ± 0.1 | 0.55 | <0.01 * | 0.12 |
| CD11b | 84.2 ± 10.9 | 86.0 ± 9.0 | 92.4 ± 6.5 | 0.45 | 0.02 * | 0.03 * | |
| CD11c | 88.2 ± 8.9 | 89.7 ± 10.6 | 94.1 ± 3.4 | 0.52 | 0.03 * | 0.17 | |
| CD18 | 97.5 ± 3.5 | 98.9 ± 2.5 | 99.9 ± 0.2 | 0.06 | 0.02 * | 0.18 | |
| Lymphocytes | CD11a | 93.8 ± 7.0 | 93.1 ± 10.2 | 99.8 ± 0.3 | 0.74 | <0.01 * | 0.03 * |
| CD11b | 25.5 ± 9.9 | 14.1 ± 6.6 | 33.6 ± 7.5 | <0.01 * | <0.01 * | <0.01 * | |
| CD11c | 11.5 ± 6.0 | 4.8 ± 2.6 | 12.8 ± 5.5 | <0.01 * | 0.51 | <0.01 * | |
| CD18 | 89.2 ± 12.3 | 90.7 ± 11.4 | 98.6 ± 4.6 | 0.60 | 0.01 * | 0.03 * |
| Resting | After Zymosan (Z) | After Opsonized Zymosan (OZ) | After PMA | |
|---|---|---|---|---|
| Day 0 | 4.39 ± 4.00 | 18.42 ± 17.56 | 59.53 ± 27.77 | 72.99 ± 29.06 |
| Day 3 | 2.28 ± 3.58 | 6.22 ± 10.97 | 47.80 ± 22.97 | 63.12 ± 30.27 |
| Adults | 2.61 ± 1.73 | 48.78 ± 34.20 | 69.02 ± 43.11 | 52.21 ± 28.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capolupo, I.; De Rose, D.U.; Pascone, R.; Danhaive, O.; Orzalesi, M. Defective Leukocyte β2 Integrin Expression and Reactive Oxygen Species Production in Neonates. Children 2022, 9, 494. https://doi.org/10.3390/children9040494
Capolupo I, De Rose DU, Pascone R, Danhaive O, Orzalesi M. Defective Leukocyte β2 Integrin Expression and Reactive Oxygen Species Production in Neonates. Children. 2022; 9(4):494. https://doi.org/10.3390/children9040494
Chicago/Turabian StyleCapolupo, Irma, Domenico Umberto De Rose, Roberto Pascone, Olivier Danhaive, and Marcello Orzalesi. 2022. "Defective Leukocyte β2 Integrin Expression and Reactive Oxygen Species Production in Neonates" Children 9, no. 4: 494. https://doi.org/10.3390/children9040494