
Allergen Exposure in Murine Neonates Promoted the Development of Asthmatic Lungs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. OVA Exposure and Challenge in Mice
2.3. Measurement of Lung Function
2.4. Eosinophil Enumeration in Bronchoalveolar Lavage Fluid (BALF) and Lung Histology
2.5. Measurement of OVA-Specific IgE (OVA-IgE)
2.6. Cell Culture and Cytokine Assays
2.7. RNA Isolation and Quantitative PCR
2.8. Statistical Analysis
3. Results
3.1. Immunological Consequences of Neonatal Exposure to OVA
3.2. Lung Parameters in Mice Subjected to Neonatal OVA Exposure
3.3. Heightened Recall Immune Responses and Hyperresponsive Airways in Neonatally OVA-Exposed Mice Following Aerosolized OVA Challenge in Adulthood
3.4. Immunological Consequences of Aerosolized OVA Exposure in Adulthood
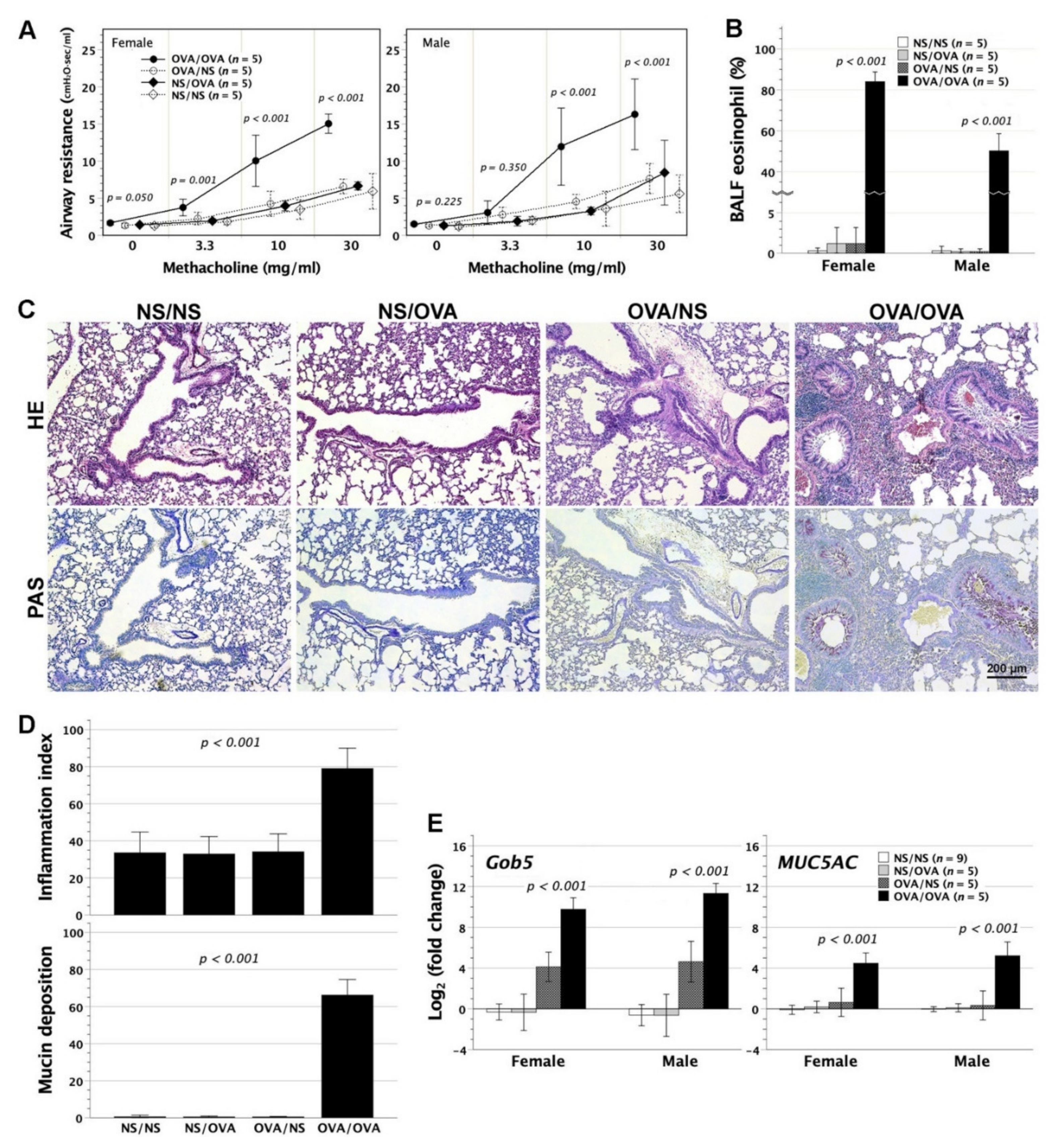
3.5. Asthmatic Lungs in Neonatally OVA-Exposed Mice Subjected to Aerosolized OVA Stress
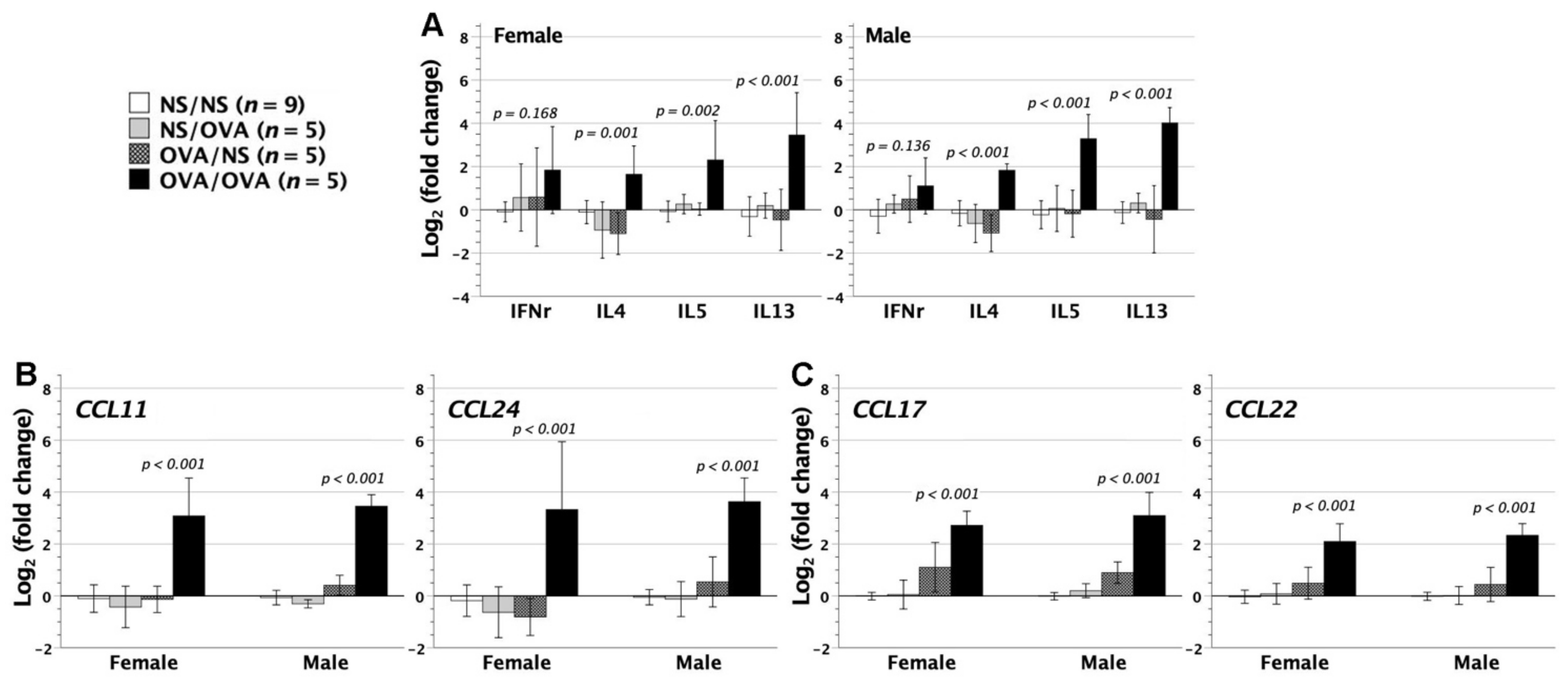
3.6. Cytokine and Chemokine Gene Expression in the Lungs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Billingham, R.E.; Brent, L.; Medawar, P.B. ‘Actively Acquired Tolerance’ of Foreign Cells. Nat. Cell Biol. 1953, 172, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-C.; Chan, C.-C.; Wu, C.-J.; Ou, L.-S.; Yu, H.-Y.; Chang, H.-L.; Tseng, L.-Y.; Kuo, M.-L. Fetal Phagocytes Take up Allergens to Initiate T-Helper Cell Type 2 Immunity and Facilitate Allergic Airway Responses. Am. J. Respir. Crit. Care Med. 2016, 194, 934–947. [Google Scholar] [CrossRef]
- Chen, J.-C.; Kuo, M.-L.; Ou, L.-S.; Chang, P.-Y.; Muench, M.O.; Shen, C.-R.; Chang, H.-L.; Yu, H.-Y.; Fu, R.-H. Characterization of Tolerance Induction through Prenatal Marrow Transplantation: The Requirement for a Threshold Level of Chimerism to Establish Rather than Maintain Postnatal Skin Tolerance. Cell Transplant. 2010, 19, 1609–1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-C. Immunological Consequences of In Utero Exposure to Foreign Antigens. Front. Immunol. 2021, 12, 638435. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.T.; Bridges, R.A.; Thomson, E. Immunological unresponsiveness in rabbits produced by neonatal injection of defined antigens. J. Exp. Med. 1958, 108, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Terres, G.; Hughes, W.L. Acquired immune tolerance in mice to crystalline bovine serum albumin. J. Immunol. 1959, 83, 459–467. [Google Scholar] [PubMed]
- Felton, L.D.; Kauffmann, G.; Prescott, B.; Ottinger, B. Studies on the mechanism of the immunological paralysis induced in mice by pneumococcal polysaccharides. J. Immunol. 1955, 74, 17–26. [Google Scholar] [PubMed]
- Forsthuber, T.; Yip, H.C.; Lehmann, P.V. Induction of TH1 and TH2 Immunity in Neonatal Mice. Science 1996, 271, 1728–1730. [Google Scholar] [CrossRef]
- Adkins, B.; Du, R.Q. Newborn mice develop balanced Th1/Th2 primary effector responses in vivo but are biased to Th2 secondary responses. J. Immunol. 1998, 160, 4217–4224. [Google Scholar]
- Rose, S.; Lichtenheld, M.; Foote, M.R.; Adkins, B. Murine Neonatal CD4+ Cells Are Poised for Rapid Th2 Effector-Like Function. J. Immunol. 2007, 178, 2667–2678. [Google Scholar] [CrossRef] [Green Version]
- Adkins, B.; Bu, Y.; Vincek, V.; Guevara, P. The Primary Responses of Murine Neonatal Lymph Node CD4+Cells are Th2-skewed and are Sufficient for the Development of Th2-biased Memory. Clin. Dev. Immunol. 2003, 10, 43–51. [Google Scholar] [CrossRef]
- Sarzotti, M.; Robbins, D.S.; Hoffman, P.M. Induction of Protective CTL Responses in Newborn Mice by a Murine Retrovirus. Science 1996, 271, 1726–1728. [Google Scholar] [CrossRef]
- Adkins, B.; Leclerc, C.; Marshall-Clarke, S. Neonatal adaptive immunity comes of age. Nat. Rev. Immunol. 2004, 4, 553–564. [Google Scholar] [CrossRef]
- Ridge, J.P.; Fuchs, E.J.; Matzinger, P. Neonatal Tolerance Revisited: Turning on Newborn T Cells with Dendritic Cells. Science 1996, 271, 1723–1726. [Google Scholar] [CrossRef] [Green Version]
- Gabet, S.; Just, J.; Couderc, R.; Seta, N.; Momas, I. Allergic sensitisation in early childhood: Patterns and related factors in PARIS birth cohort. Int. J. Hyg. Environ. Health 2016, 219, 792–800. [Google Scholar] [CrossRef]
- Verhasselt, V.; Milcent, V.; Cazareth, J.; Kanda, A.; Fleury, S.; Dombrowicz, D.; Glaichenhaus, N.; Julia, V. Breast milk–mediated transfer of an antigen induces tolerance and protection from allergic asthma. Nat. Med. 2008, 14, 170–175. [Google Scholar] [CrossRef]
- Mosconi, E.; Rekima, A.; Seitz-Polski, B.; Kanda, A.; Fleury, S.; Tissandie, E.; Monteiro, R.; Dombrowicz, D.D.; Julia, V.; Glaichenhaus, N.; et al. Breast milk immune complexes are potent inducers of oral tolerance in neonates and prevent asthma development. Mucosal Immunol. 2010, 3, 461–474. [Google Scholar] [CrossRef] [Green Version]
- Skobeloff, E.M.; Spivey, W.H.; Clair, S.S.S.; Schoffstall, J.M. The Influence of Age and Sex on Asthma Admissions. JAMA 1992, 268, 3437–3440. [Google Scholar] [CrossRef]
- De Marco, R.; Locatelli, F.; Sunyer, J.; Burney, P. The European Community Respiratory Health Survey Study Group. Differences in Incidence of Reported Asthma Related to Age in Men and Women. Am. J. Respir. Crit. Care Med. 2000, 162, 68–74. [Google Scholar] [CrossRef]
- Melgert, B.N.; Postma, D.S.; Kuipers, I.; Geerlings, M.; Luinge, M.A.; Strate, B.W.A.; Kerstjens, H.A.M.; Timens, W.; Hylkema, M.N. Female mice are more susceptible to the development of allergic airway inflammation than male mice. Clin. Exp. Allergy 2005, 35, 1496–1503. [Google Scholar] [CrossRef]
- Chan, C.-C.; Lai, C.-W.; Wu, C.-J.; Chen, L.-C.; Tao, M.-H.; Kuo, M.-L. Liver-Specific Allergen Gene Transfer by Adeno-Associated Virus Suppresses Allergic Airway Inflammation in Mice. Hum. Gene Ther. 2016, 27, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Spandidos, A.; Wang, X.; Wang, H.; Seed, B. PrimerBank: A resource of human and mouse PCR primer pairs for gene expression detection and quantification. Nucleic Acids Res. 2009, 38, D792–D799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.A.; Peluso, J.J.; Shanley, J.D.; Puddington, L.; Thrall, R.S. Murine Cytomegalovirus Influences Foxj1 Expression, Ciliogenesis, and Mucus Plugging in Mice with Allergic Airway Disease. Am. J. Pathol. 2008, 172, 714–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, A.; Morita, S.; Iwashita, H.; Sagiya, Y.; Ashida, Y.; Shirafuji, H.; Fujisawa, Y.; Nishimura, O.; Fujino, M. Role of gob-5 in mucus overproduction and airway hyperresponsiveness in asthma. Proc. Natl. Acad. Sci. USA 2001, 98, 5175–5180. [Google Scholar] [CrossRef] [Green Version]
- Long, A.J.; Sypek, J.P.; Askew, R.; Fish, S.C.; Mason, L.E.; Williams, C.M.M.; Goldman, S.J. Gob-5 Contributes to Goblet Cell Hyperplasia and Modulates Pulmonary Tissue Inflammation. Am. J. Respir. Cell Mol. Biol. 2006, 35, 357–365. [Google Scholar] [CrossRef]
- Bonser, L.R.; Erle, D.J. Airway Mucus and Asthma: The Role of MUC5AC and MUC5B. J. Clin. Med. 2017, 6, 112. [Google Scholar] [CrossRef] [Green Version]
- Menzies-Gow, A.; Ying, S.; Sabroe, I.; Stubbs, V.L.; Soler, D.; Williams, T.J.; Kay, A.B. Eotaxin (CCL11) and Eotaxin-2 (CCL24) Induce Recruitment of Eosinophils, Basophils, Neutrophils, and Macrophages As Well As Features of Early- and Late-Phase Allergic Reactions Following Cutaneous Injection in Human Atopic and Nonatopic Volunteers. J. Immunol. 2002, 169, 2712–2718. [Google Scholar] [CrossRef]
- Garcia-Zepeda, E.A.; Rothenberg, M.E.; Ownbey, R.T.; Celestin, J.; Leder, P.; Luster, A.D. Human eotaxin is a specific chemoattractant for eosinophil cells and provides a new mechanism to explain tissue eosinophilia. Nat. Med. 1996, 2, 449–456. [Google Scholar] [CrossRef]
- White, J.R.; Imburgia, C.; Dul, E.; Appelbaum, E.; O’Donnell, K.; O’Shannessy, D.J.; Brawner, M.; Fornwald, J.; Adamou, J.; Elshourbagy, N.A.; et al. Cloning and functional characterization of a novel human CC chemokine that binds to the CCR3 receptor and activates human eosinophils. J. Leukoc. Biol. 1997, 62, 667–675. [Google Scholar] [CrossRef]
- Scheu, S.; Ali, S.; Ruland, C.; Arolt, V.; Alferink, J. The C-C Chemokines CCL17 and CCL22 and Their Receptor CCR4 in CNS Autoimmunity. Int. J. Mol. Sci. 2017, 18, 2306. [Google Scholar] [CrossRef] [Green Version]
- Tsunemi, Y.; Saeki, H.; Nakamura, K.; Nagakubo, D.; Nakayama, T.; Yoshie, O.; Kagami, S.; Shimazu, K.; Kadono, T.; Sugaya, M.; et al. CCL17 transgenic mice show an enhanced Th2-type response to both allergic and non-allergic stimuli. Eur. J. Immunol. 2006, 36, 2116–2127. [Google Scholar] [CrossRef]
- Abboud, D.; Daubeuf, F.; Do, Q.T.; Utard, V.; Villa, P.; Haiech, J.; Bonnet, D.; Hibert, M.; Bernard, P.; Galzi, J.-L.; et al. A strategy to discover decoy chemokine ligands with an anti-inflammatory activity. Sci. Rep. 2015, 5, 14746. [Google Scholar] [CrossRef] [Green Version]
- Sears, M.R. Trends in the Prevalence of Asthma. Chest 2014, 145, 219–225. [Google Scholar] [CrossRef]
- Lloyd, C.M.; Hessel, E.M. Functions of T cells in asthma: More than just TH2 cells. Nat. Rev. Immunol. 2010, 10, 838–848. [Google Scholar] [CrossRef]
- Prescott, S.L.; Macaubas, C.; Smallacombe, T.; Holt, B.J.; Sly, P.; Holt, P.G. Development of allergen-specific T-cell memory in atopic and normal children. Lancet 1999, 353, 196–200. [Google Scholar] [CrossRef]
- Grieger, J.A.; Clifton, V.L.; Tuck, A.R.; Wooldridge, A.L.; Robertson, S.A.; Gatford, K.L. In utero Programming of Allergic Susceptibility. Int. Arch. Allergy Immunol. 2016, 169, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Lockett, G.A.; Huoman, J.; Holloway, J.W. Does allergy beginin utero? Pediatr. Allergy Immunol. 2015, 26, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Vance, G.H.S.; Holloway, J. Early life exposure to dietary and inhalant allergens. Pediatr. Allergy Immunol. 2002, 13, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Edelbauer, M.; Loibichler, C.; Nentwich, I.; Gerstmayr, M.; Urbanek, R.; Szepfalusi, Z. Maternally delivered nutritive allergens in cord blood and in placental tissue of term and preterm neonates. Clin. Exp. Allergy 2004, 34, 189–193. [Google Scholar] [CrossRef]
- Holloway, J.; O Warner, J.; Vance, G.H.; Diaper, N.D.; A Warner, J.; A Jones, C. Detection of house-dust-mite allergen in amniotic fluid and umbilical-cord blood. Lancet 2000, 356, 1900–1902. [Google Scholar] [CrossRef]
- Sporik, R.; Holgate, S.T.; Platts-Mills, T.A.; Cogswell, J.J. Exposure to House-Dust Mite Allergen (Der pI) and the Development of Asthma in Childhood. N. Engl. J. Med. 1990, 323, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Gern, J.E.; Lemanske, R.F.; Busse, W.W. Early life origins of asthma. J. Clin. Investig. 1999, 104, 837–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, S.; Sherrill, D.L.; Martinez, F.D.; Barbee, R.A. Rhinitis as an independent risk factor for adult-onset asthma. J. Allergy Clin. Immunol. 2002, 109, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Hogan, S.P.; Foster, P.S.; Charlton, B.; Slattery, R.M. Prevention of Th2-mediated murine allergic airways disease by soluble antigen administration in the neonate. Proc. Natl. Acad. Sci. USA 1998, 95, 2441–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; McCusker, C. Neonatal exposure with LPS and/or allergen prevents experimental allergic airways disease: Development of tolerance using environmental antigens. J. Allergy Clin. Immunol. 2006, 118, 143–151. [Google Scholar] [CrossRef]
- Zosky, G.; Sly, P. Animal models of asthma. Clin. Exp. Allergy 2007, 37, 973–988. [Google Scholar] [CrossRef]
- Nials, A.T.; Uddin, S. Mouse models of allergic asthma: Acute and chronic allergen challenge. Dis. Model. Mech. 2008, 1, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Ostroukhova, M.; Seguin-Devaux, C.; Oriss, T.B.; Dixon-McCarthy, B.; Yang, L.; Ameredes, B.T.; Corcoran, T.E.; Ray, A. Tolerance induced by inhaled antigen involves CD4(+) T cells expressing membrane-bound TGF-beta and FOXP3. J. Clin. Investig. 2004, 114, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Brewer, J.M.; Conacher, M.; Hunter, C.A.; Mohrs, M.; Brombacher, F.; Alexander, J. Aluminium hydroxide adjuvant initiates strong antigen-specific Th2 responses in the absence of IL-4- or IL-13-mediated signaling. J. Immunol. 1999, 163, 6448–6454. [Google Scholar]
- Blyth, D.I.; Pedrick, M.S.; Savage, T.J.; Hessel, E.M.; Fattah, D. Lung inflammation and epithelial changes in a murine model of atopic asthma. Am. J. Respir. Cell Mol. Biol. 1996, 14, 425–438. [Google Scholar] [CrossRef]
- Janssen, E.M.; Van Oosterhout, A.J.M.; Nijkamp, F.P.; Van Eden, W.; Wauben, M.H.M. The efficacy of immunotherapy in an experimental murine model of allergic asthma is related to the strength and site of T cell activation during immunotherapy. J. Immunol. 2000, 165, 7207–7214. [Google Scholar] [CrossRef] [Green Version]
- Corteling, R.; Trifilieff, A. Gender comparison in a murine model of allergen-driven airway inflammation and the response to budesonide treatment. BMC Pharmacol. 2004, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Takeda, M.; Tanabe, M.; Ito, W.; Ueki, S.; Konnno, Y.; Chihara, M.; Itoga, M.; Kobayashi, Y.; Moritoki, Y.; Kayaba, H.; et al. Gender difference in allergic airway remodelling and immunoglobulin production in mouse model of asthma. Respirology 2013, 18, 797–806. [Google Scholar] [CrossRef]
- Blacquière, M.; Hylkema, M.; Postma, D.; Geerlings, M.; Timens, W.; Melgert, B. Airway Inflammation and Remodeling in Two Mouse Models of Asthma: Comparison of Males and Females. Int. Arch. Allergy Immunol. 2010, 153, 173–181. [Google Scholar] [CrossRef]
- Okuyama, K.; Wada, K.; Chihara, J.; Takayanagi, M.; Ohno, I. Sex-related splenocyte function in a murine model of allergic asthma. Clin. Exp. Allergy 2008, 38, 1212–1219. [Google Scholar] [CrossRef]
- Cai, Y.; Zhou, J.; Webb, D.C. Estrogen Stimulates Th2 Cytokine Production and Regulates the Compartmentalisation of Eosinophils during Allergen Challenge in a Mouse Model of Asthma. Int. Arch. Allergy Immunol. 2012, 158, 252–260. [Google Scholar] [CrossRef]
- Riffo-Vasquez, Y.; de Oliveira, A.P.L.; Page, C.P.; Spina, D.; Tavares-De-Lima, W. Role of sex hormones in allergic inflammation in mice. Clin. Exp. Allergy 2007, 37, 459–470. [Google Scholar] [CrossRef]
- Nunn, C.L.; Lindenfors, P.; Pursall, E.R.; Rolff, J. On sexual dimorphism in immune function. Philos. Trans. R. Soc. B Biol. Sci. 2008, 364, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Gal-Oz, S.T.; Maier, B.; Yoshida, H.; Seddu, K.; Elbaz, N.; Czysz, C.; Zuk, O.; Stranger, B.E.; Ner-Gaon, H.; Shay, T. ImmGen report: Sexual dimorphism in the immune system transcriptome. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The Cytokines of Asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef]
- Brusselle, G.; Kips, J.; Joos, G.; Bluethmann, H.; Pauwels, R. Allergen-induced airway inflammation and bronchial responsiveness in wild-type and interleukin-4-deficient mice. Am. J. Respir. Cell Mol. Biol. 1995, 12, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Busse, W.W. Leukotrienes and Inflammation. Am. J. Respir. Crit. Care Med. 1998, 157, S210–S213. [Google Scholar] [CrossRef] [PubMed]
- Grünig, G.; Warnock, M.; Wakil, A.E.; Venkayya, R.; Brombacher, F.; Rennick, D.M.; Sheppard, D.; Mohrs, M.; Donaldson, D.D.; Locksley, R.M.; et al. Requirement for IL-13 Independently of IL-4 in Experimental Asthma. Science 1998, 282, 2261–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perros, F.; Hoogsteden, H.C.; Coyle, A.J.; Lambrecht, B.N.; Hammad, H. Blockade of CCR4 in a humanized model of asthma reveals a critical role for DC-derived CCL17 and CCL22 in attracting Th2 cells and inducing airway inflammation. Allergy 2009, 64, 995–1002. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.-C.; Chan, C.-C.; Ting, N.-C.; Kuo, M.-L. Allergen Exposure in Murine Neonates Promoted the Development of Asthmatic Lungs. Biomedicines 2021, 9, 688. https://doi.org/10.3390/biomedicines9060688
Chen J-C, Chan C-C, Ting N-C, Kuo M-L. Allergen Exposure in Murine Neonates Promoted the Development of Asthmatic Lungs. Biomedicines. 2021; 9(6):688. https://doi.org/10.3390/biomedicines9060688
Chicago/Turabian StyleChen, Jeng-Chang, Cheng-Chi Chan, Nai-Chun Ting, and Ming-Ling Kuo. 2021. "Allergen Exposure in Murine Neonates Promoted the Development of Asthmatic Lungs" Biomedicines 9, no. 6: 688. https://doi.org/10.3390/biomedicines9060688