
Survival Bias and Crosstalk between Chronological and Behavioral Age: Age- and Genotype-Sensitivity Tests Define Behavioral Signatures in Middle-Aged, Old, and Long-Lived Mice with Normal and AD-Associated Aging

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Behavioral Assessments
2.3.1. Day 1. Corner and Open-Field Tests (CT and OF)
2.3.2. Day 2. Recognition Tests (OF2 and OR)
2.3.3. Day 3. Spontaneous Alternation in the T-Maze (TM-SA)
2.3.4. Day 4 to 9. Morris Water Maze Test (MWM)
2.4. Statistics
3. Results
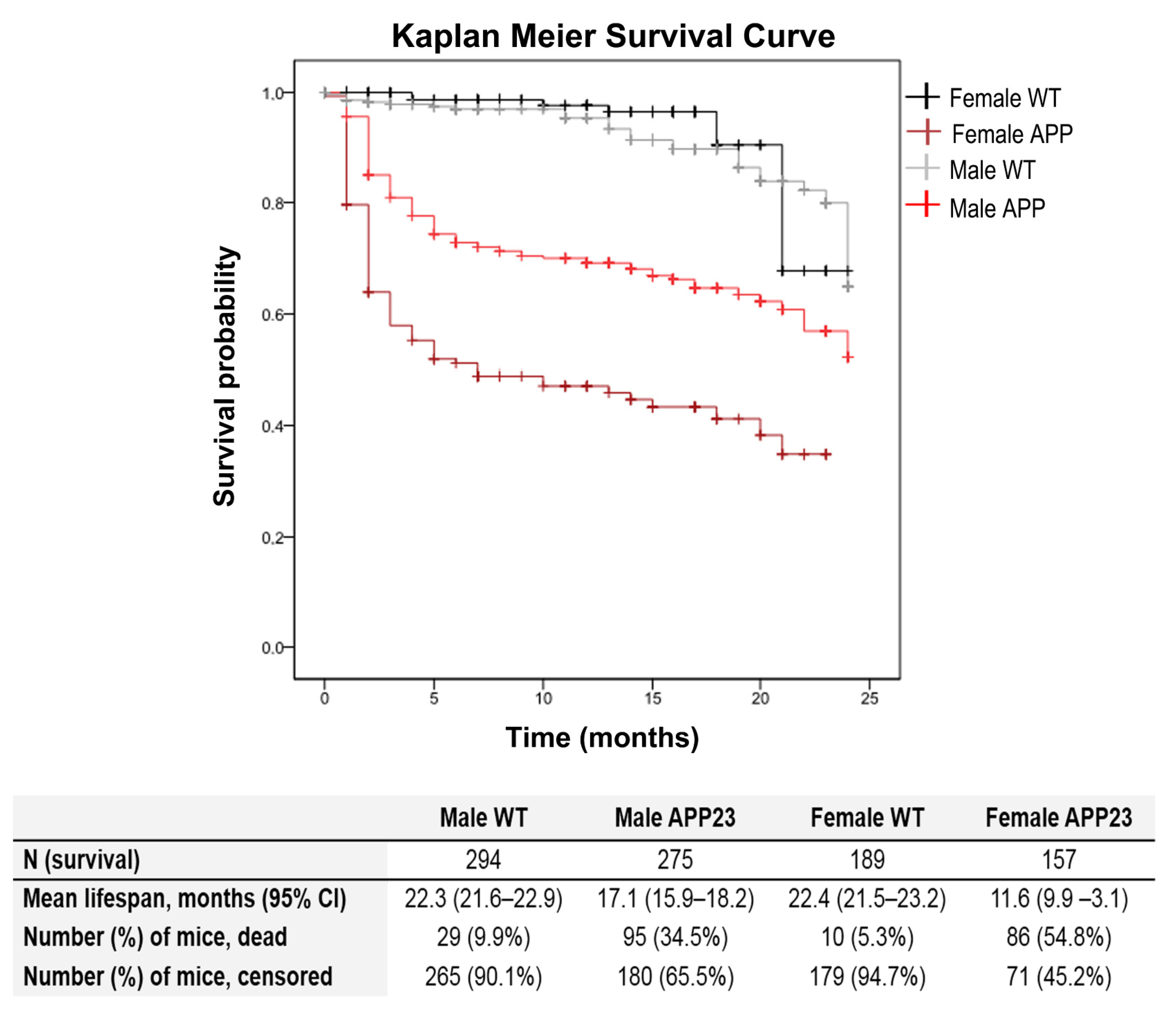
3.1. Genotype and Sex-Dependent Increased Mortality Rates
3.2. Strong Contribution of Age and Specificity of Genotype Factor on Key Variables for AD-Related Phenotype
4. Discussion
5. Conclusions and Future Directions
- (1).
- Survival curves of 920 mice of APP23 and WT C57BL/6 littermates confirmed genotype and sex-dependent increased mortality rates, and long-living males.
- (2).
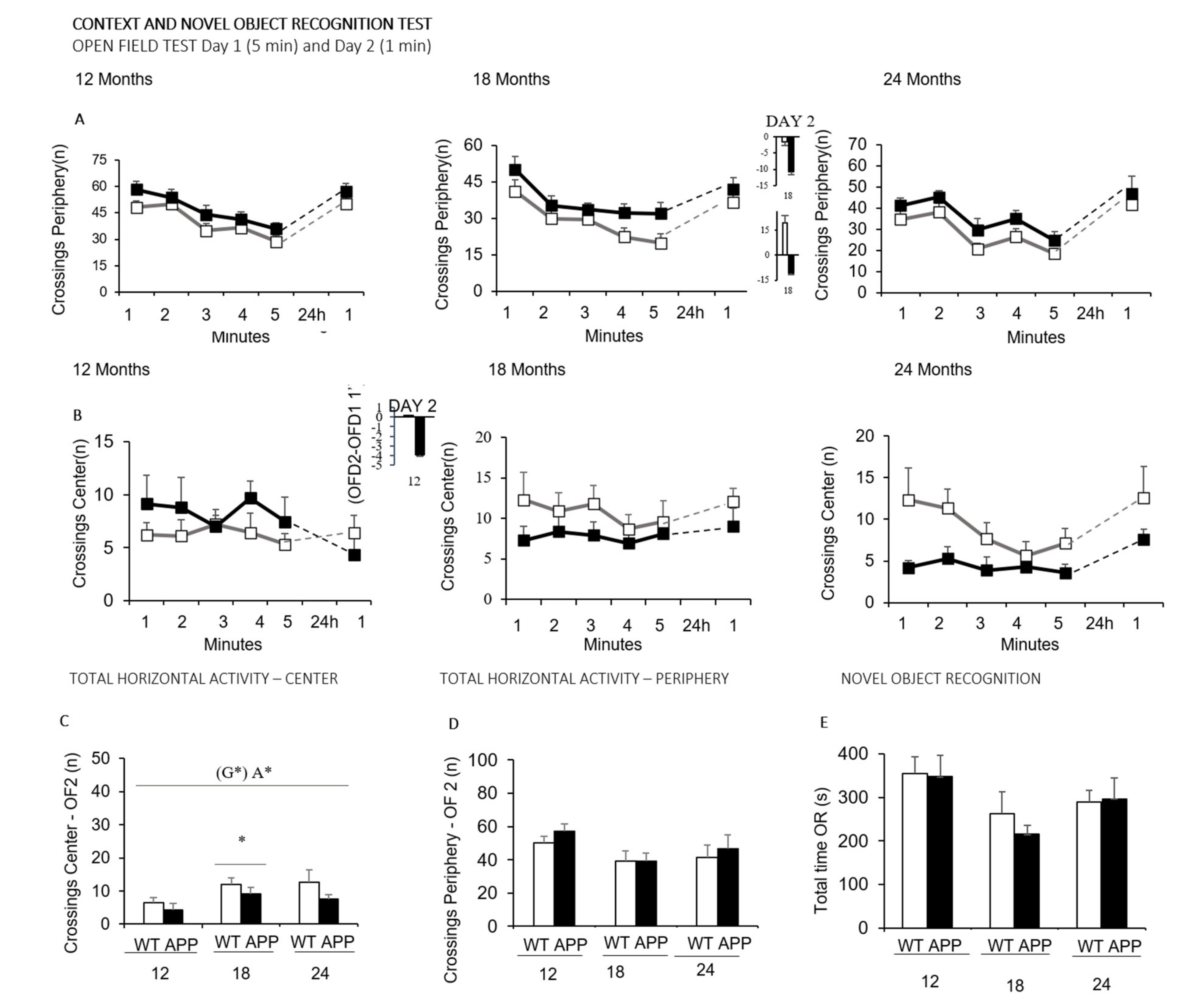
- Compared to WT littermates, the APP23 mice showed an increased number of visited corners but decreased vertical exploratory activity, evidencing increased neophobia in the corner test. Similarly, APP23 groups showed increased latencies to develop the ethogram of behaviors immediately after the direct exposure to an open and illuminated field (not shown) and increased locomotion, with increased thigmotaxis (or search for the protected peripheral area), primarily noticeable at 12 months of age.
- (3).
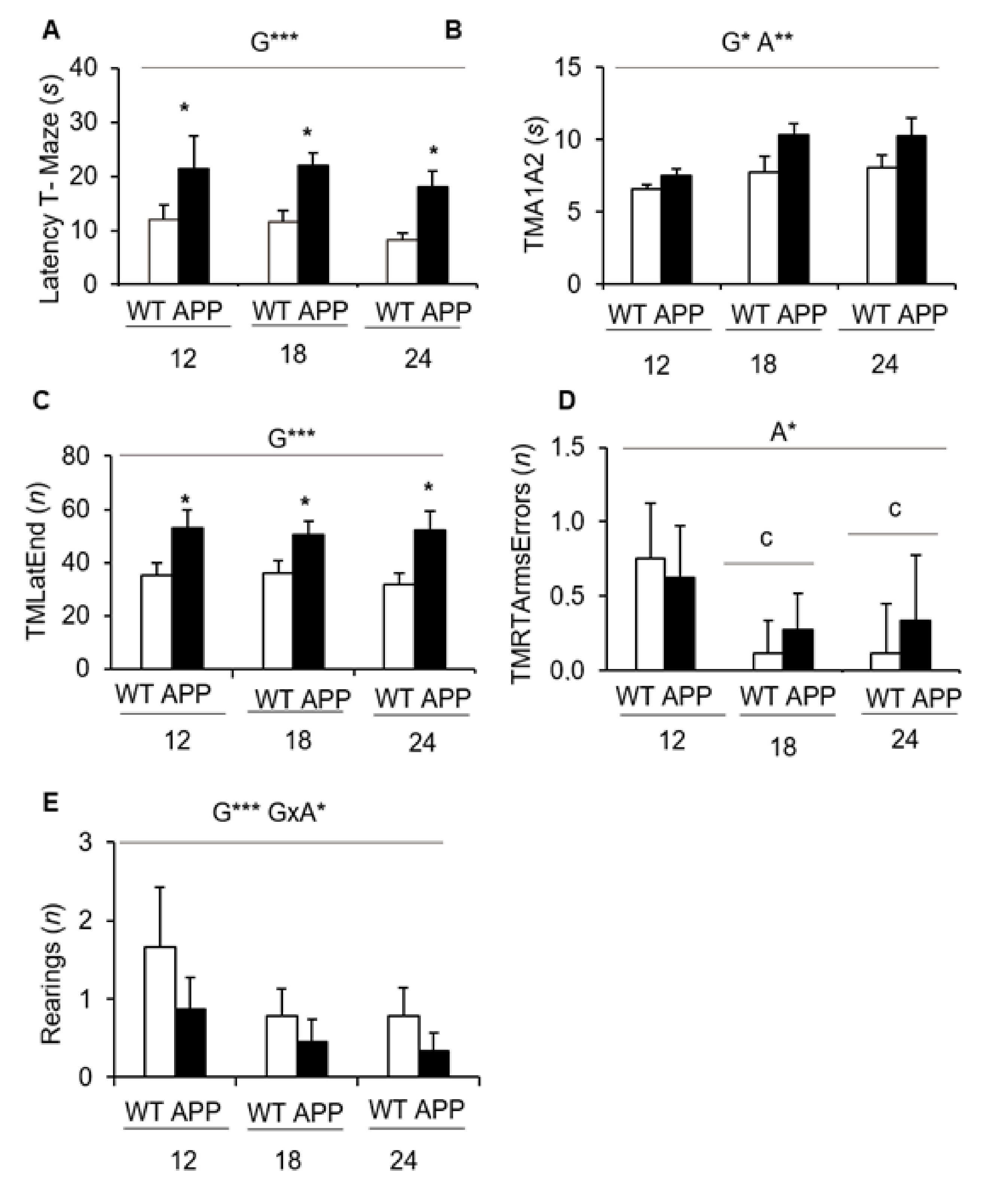
- In the T-maze, a black T-shaped corridor resembling burrows, the latency to reach the intersection was consistently shown to be increased at all ages and the longer time required for APP23 mice to complete the exploration of all the arms of the maze. As part of the differences in the status of their cognitive functions exhibited during the exploration of new environments, we assessed APP23 mice for putative cognitive deficits in discriminative tasks (novelty/familiarity) for context (remembering the anxiogenic open field scenario) and object recognition.
- (4).
- The novel object recognition test showed low sensitivity to the genotype, only noticed at the oldest age through the lowest difference in the delay in exploring the novel and familiar objects compared to WT littermates.
- (5).
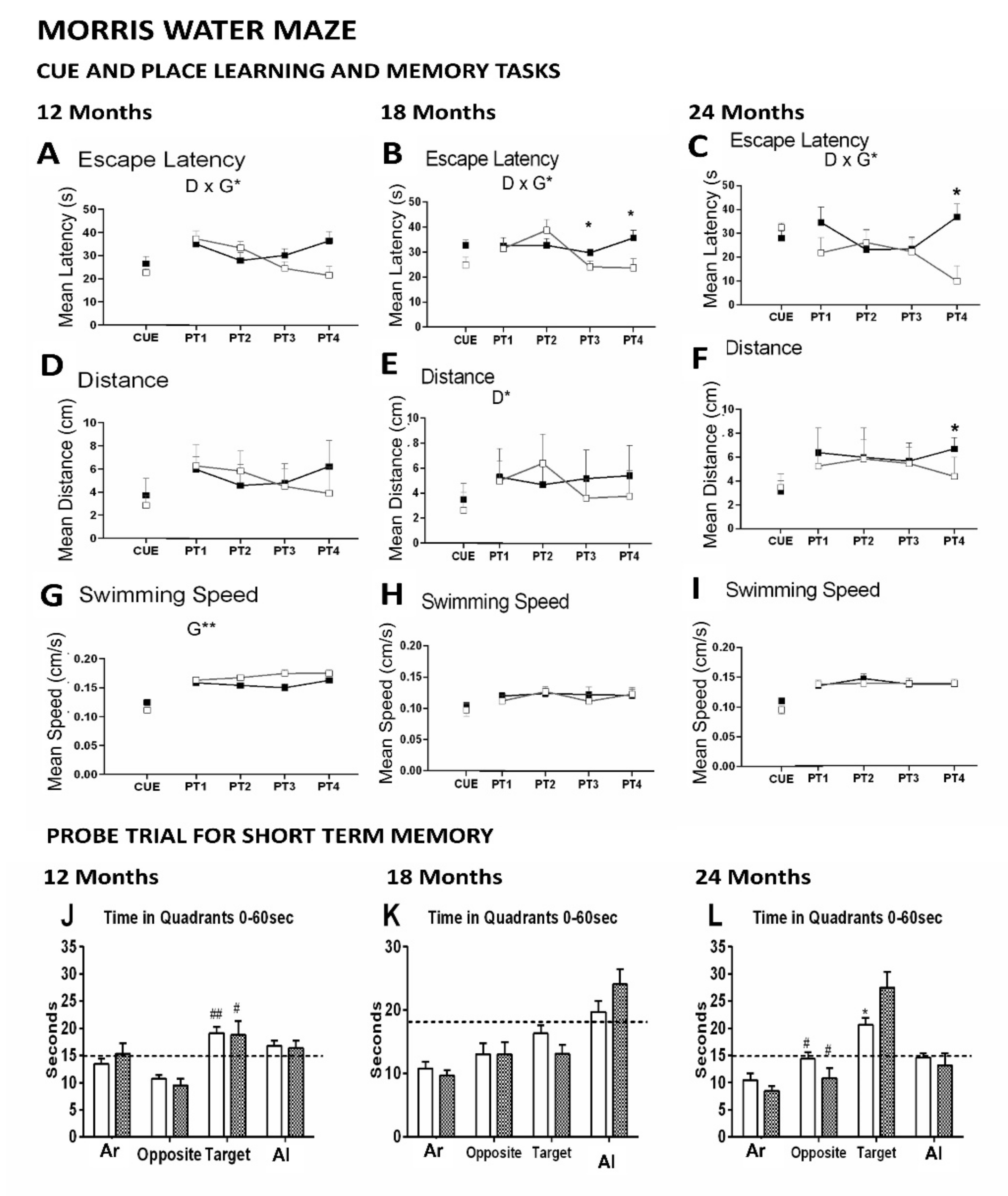
- Cognitive deficits in Spatial reference learning and memory were assessed in the Morris water maze through a 4-day place learning task using a hidden platform followed by a probe trial for long-term memory where the platform had been removed. Worse performance of 12-month-old APP23 mice was shown as compared to WT littermates during the progress of acquisition (learning and memory) in the place task, while differences were scarce in the older groups. In the probe trial, the worse spatial memory discrimination of the trained quadrant compared to the opposed quadrant was shown at 18 months of age but independently of the genotype. As we have reported in other mice models for AD, a conspicuous BPSD-like profile and age as a biological factor may result in confounding factors in an aquatic maze, considered an anxiogenic environment for mice.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brookmeyer, R.; Johnson, E.; Ziegler-Graham, K.; Arrighi, H.M. Forecasting the global burden of Alzheimer’s disease. Alzheimer’s Dement. 2007, 3, 186–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, K.; Simpkins, J.W.; Ji, X.; Leis, M.; Stambler, I. The Critical Need to Promote Research of Aging and Aging-related Diseases to Improve Health and Longevity of the Elderly Population. Aging Dis. 2015, 6, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO); United Nations (UN). Population Prospects: The 2017 Revision, Key Findings and Advance Tables; Working Paper No. ESA/P/WP/248; United Nations Department of Economic and Social Affairs/Population Division: Nairobi, Kenya, 2017. [Google Scholar]
- Arai, H.; Ouchi, Y.; Yokode, M.; Ito, H.; Uematsu, H.; Eto, F.; Oshima, S.; Ota, K.; Saito, Y.; Sasaki, H.; et al. Toward the realization of a better aged society: Messages from gerontology and geriatrics. Geriatr. Gerontol. Int. 2011, 12, 16–22. [Google Scholar] [CrossRef] [PubMed]
- WHO. Mental Health of Older Adults 2016. Available online: https://www.who.int/news-room/fact-sheets/detail/mental-health-of-older-adults (accessed on 27 December 2020).
- Fornaro, M.; Solmi, M.; Stubbs, B.; Veronese, N.; Monaco, F.; Novello, S.; Fusco, A.; Anastasia, A.; De Berardis, D.; Carvalho, A.F.; et al. Prevalence and correlates of major depressive disorder, bipolar disorder and schizophrenia among nursing home residents without dementia: Systematic review and meta-analysis. Br. J. Psychiatry 2019, 216, 6–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, J.L. Cognitive and behavioral heterogeneity in Alzheimer’s disease: Seeking the neurobiological basis. Neurobiol. Aging 2000, 21, 845–861. [Google Scholar] [CrossRef]
- Reisberg, B.; Ferris, S.H.; Franssen, E.H.; Shulman, E.; Monteiro, I.; Sclan, S.G.; Steinberg, G.; Kluger, A.; Torossian, C.; De Leon, M.J.; et al. Mortality and Temporal Course of Probable Alzheimer’s Disease: A 5-Year Prospective Study. Int. Psychogeriatr. 1996, 8, 291–311. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A. Ekavali A review on Alzheimer’s disease pathophysiology and its management: An update. Pharmacol. Rep. 2015, 67, 195–203. [Google Scholar] [CrossRef]
- Jeste, D.V.; Blazer, D.; Casey, D.; Meeks, T.; Salzman, C.; Schneider, L.; Tariot, P.; Yaffe, K. ACNP White Paper: Update on Use of Antipsychotic Drugs in Elderly Persons with Dementia. Neuropsychopharmacology 2007, 33, 957–970. [Google Scholar] [CrossRef]
- Magierski, R.; Sobow, T.; Schwertner, E.; Religa, D. Pharmacotherapy of Behavioral and Psychological Symptoms of Dementia: State of the Art and Future Progress. Front. Pharmacol. 2020, 11, 1168. [Google Scholar] [CrossRef]
- Mirakhur, A.; Craig, D.; Hart, D.J.; Mcllroy, S.P.; Passmore, A.P. Behavioural and psychological syndromes in Alzheimer’s disease. Int. J. Geriatr. Psychiatry 2004, 19, 1035–1039. [Google Scholar] [CrossRef]
- Spalletta, G.; Baldinetti, F.; Buccione, I.; Fadda, L.; Perri, R.; Scalmana, S.; Serra, L.; Caltagirone, C. Cognition and behaviour are independent and heterogeneous dimensions in Alzheimer?s disease. J. Neurol. 2004, 251, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Llort, L.; Johansson, B. Editorial: Pharmacology of BPSD (Behavioral and Psychological Symptoms of Dementia). Front. Pharmacol. Neuropharmacol. 2021. [Google Scholar] [CrossRef]
- Yunusa, I.; El Helou, M.L. The Use of Risperidone in Behavioral and Psychological Symptoms of Dementia: A Review of Pharmacology, Clinical Evidence, Regulatory Approvals, and Off-Label Use. Front. Pharmacol. 2020, 11, 596. [Google Scholar] [CrossRef] [PubMed]
- Piersanti, M.; Capannolo, M.; Turchetti, M.; Serroni, N.; De Berardis, D.; Evangelista, P.; Costantini, P.; Orsini, A.; Rossi, A.; Maggio, R. Increase in mortality rate in patients with dementia treated with atypical antipsychotics: A cohort study in outpatients in Central Italy. Riv Psichiatr. 2014, 49, 34–40. [Google Scholar] [PubMed]
- Torres-Lista, V.; López-Pousa, S.; Giménez-Llort, L. Impact of Chronic Risperidone Use on Behavior and Survival of 3xTg-AD Mice Model of Alzheimer’s Disease and Mice With Normal Aging. Front. Pharmacol. 2019, 10, 1061. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Llort, L.; Blázquez, G.; Cañete, T.; Johansson, B.; Oddo, S.; Tobeña, A.; LaFerla, F.; Fernandez-Teruel, A. Modeling behavioral and neuronal symptoms of Alzheimer’s disease in mice: A role for intraneuronal amyloid. Neurosci. Biobehav. Rev. 2007, 31, 125–147. [Google Scholar] [CrossRef]
- Gimenez-Llort, L.; Torres-Lista, V.; Fuente, M. Crosstalk between Behavior and Immune System during the Prodromal Stages of Alzheimer’s Disease. Curr. Pharm. Des. 2014, 20, 4723–4732. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Altayo, F.; Sànchez-Ventura, J.; Vila, E.; Giménez-Llort, L. Crosstalk between Peripheral Small Vessel Properties and Anxious-like Profiles: Sex, Genotype, and Interaction Effects in Mice with Normal Aging and 3×Tg-AD mice at Advanced Stages of Disease. J. Alzheimer’s Dis. 2018, 62, 1531–1538. [Google Scholar] [CrossRef]
- Komarova, N.L.; Thalhauser, C.J. High Degree of Heterogeneity in Alzheimer’s Disease Progression Patterns. PLoS Comput. Biol. 2011, 7, e1002251. [Google Scholar] [CrossRef] [Green Version]
- Torres-Lista, V.; De La Fuente, M.; Giménez-Llort, L. Survival Curves and Behavioral Profiles of Female 3xTg-AD Mice Surviving to 18-Months of Age as Compared to Mice with Normal Aging. J. Alzheimer’s Dis. Rep. 2017, 1, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Boix, P.; Giménez-Llort, L. Comorbid sensorimotor and emotional profiles in the forced swim test immobility and predictive value of a single assay in very old female mice. Exp. Gerontol. 2019, 120, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Muntsant, A.; Giménez-Llort, L. Impact of Social Isolation on the Behavioral, Functional Profiles, and Hippocampal Atrophy Asymmetry in Dementia in Times of Coronavirus Pandemic (COVID-19): A Translational Neuroscience Approach. Front. Psychiatry 2020, 11, 572583. [Google Scholar] [CrossRef]
- Spreng, R.N.; Turner, G.R. The Shifting Architecture of Cognition and Brain Function in Older Adulthood. Perspect. Psychol. Sci. 2019, 14, 523–542. [Google Scholar] [CrossRef] [PubMed]
- Amer, T.; Giovanello, K.S.; Grady, C.L.; Hasher, L. Age differences in memory for meaningful and arbitrary associations: A memory retrieval account. Psychol. Aging 2018, 33, 74–81. [Google Scholar] [CrossRef]
- Turner, R.C.; Seminerio, M.J.; Naser, Z.J.; Ford, J.N.; Martin, S.J.; Matsumoto, R.R.; Rosen, C.L.; Huber, J.D. Effects of aging on behavioral assessment performance: Implications for clinically relevant models of neurological disease. J. Neurosurg. 2012, 117, 629–637. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Scheibye-Knudsen, M.; Longo, D.L.; De Cabo, R. Animal Models of Aging Research: Implications for Human Aging and Age-Related Diseases. Annu. Rev. Anim. Biosci. 2015, 3, 283–303. [Google Scholar] [CrossRef] [Green Version]
- Puzzo, D.; Gulisano, W.; Palmeri, A.; Arancio, O. Rodent models for Alzheimer’s disease drug discovery. Expert Opin. Drug Discov. 2015, 10, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Pietropaolo, S.; Feldon, J.; Yee, B.K. Age-dependent phenotypic characteristics of a triple transgenic mouse model of Alzheimer disease. Behav. Neurosci. 2008, 122, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, M.; Bowers, W.J. Detailed immunohistochemical characterization of temporal and spatial progression of Alzheimer’s disease-related pathologies in male triple-transgenic mice. BMC Neurosci. 2008, 9, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahlström, A.; Zeberg, H.; Ulfhake, B. Changes in behaviors of male C57BL/6J mice across adult life span and effects of dietary restriction. Age 2012, 34, 1435–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giménez-Llort, L.; Ramírez-Boix, P.; De La Fuente, M. Mortality of septic old and adult male mice correlates with individual differences in premorbid behavioral phenotype and acute-phase sickness behavior. Exp. Gerontol. 2019, 127, 110717. [Google Scholar] [CrossRef]
- Torres-Lista, V.; Giménez-Llort, L. Persistence of behaviours in the Forced Swim Test in 3xTg-AD mice at advanced stages of disease. Behav. Process. 2014, 106, 118–121. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Arranz, L.; Maté, I.; De La Fuente, M. Gender-Specific Neuroimmunoendocrine Aging in a Triple-Transgenic 3×Tg-AD Mouse Model for Alzheimer’s Disease and Its Relation with Longevity. Neuroimmunomodulation 2008, 15, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Muntsant, A.; Jiménez-Altayó, F.; Puertas-Umbert, L.; Jiménez-Xarrie, E.; Vila, E.; Giménez-Llort, L. Sex-Dependent End-of-Life Mental and Vascular Scenarios for Compensatory Mechanisms in Mice with Normal and AD-Neurodegenerative Aging. Biomedicines 2021, 9, 111. [Google Scholar] [CrossRef]
- Van Dijk, P.T.; Dippel, D.W.; Habbema, J.D.F. Survival of Patients with Dementia. J. Am. Geriatr. Soc. 1991, 39, 603–610. [Google Scholar] [CrossRef]
- Zeng, A.; Song, X.; Dong, J.; Mitnitski, A.; Liu, J.; Guo, Z.; Rockwood, K. Mortality in Relation to Frailty in Patients Admitted to a Specialized Geriatric Intensive Care Unit. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2015, 70, 1586–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Brown, M.; Sampson, E.L.; Jones, L.; Barron, A.M. Prognostic indicators of 6-month mortality in elderly people with advanced dementia: A systematic review. Palliat. Med. 2012, 27, 389–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pike, C.J. Sex and the development of Alzheimer’s disease. J. Neurosci. Res. 2017, 95, 671–680. [Google Scholar] [CrossRef] [Green Version]
- Rezzani, R.; Franco, C.; Rodella, L.F. Sex differences of brain and their implications for personalized therapy. Pharmacol. Res. 2019, 141, 429–442. [Google Scholar] [CrossRef]
- Sturchler-Pierrat, C.; Staufenbiel, M. Pathogenic Mechanisms of Alzheimer’s Disease Analyzed in the APP23 Transgenic Mouse Model. Ann. N. Y. Acad. Sci. 2006, 920, 134–139. [Google Scholar] [CrossRef]
- Hirata-Fukae, C.; Li, H.-F.; Hoe, H.-S.; Gray, A.J.; Minami, S.S.; Hamada, K.; Niikura, T.; Hua, F.; Tsukagoshi-Nagai, H.; Horikoshi-Sakuraba, Y.; et al. Females exhibit more extensive amyloid, but not tau, pathology in an Alzheimer transgenic model. Brain Res. 2008, 1216, 92–103. [Google Scholar] [CrossRef]
- Rae, E.A.; Brown, R.E. The problem of genotype and sex differences in life expectancy in transgenic AD mice. Neurosci. Biobehav. Rev. 2015, 57, 238–251. [Google Scholar] [CrossRef]
- Kane, A.E.; Shin, S.; Wong, A.A.; Fertan, E.; Faustova, N.S.; Howlett, S.E.; Brown, R.E. Sex Differences in Healthspan Predict Lifespan in the 3xTg-AD Mouse Model of Alzheimer’s Disease. Front. Aging Neurosci. 2018, 10, 172. [Google Scholar] [CrossRef] [Green Version]
- Giménez-Llort, L.; Maté, I.; Manassra, R.; Vida, C.; De La Fuente, M. Peripheral immune system and neuroimmune communication impairment in a mouse model of Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2012, 1262, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Montacute, R.; Foley, K.; Forman, R.; Else, K.J.; Cruickshank, S.M.; Allan, S.M. Enhanced susceptibility of triple transgenic Alzheimer’s disease (3xTg-AD) mice to acute infection. J. Neuroinflamm. 2017, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Mesa, Y.; Giménez-Llort, L.; López, L.C.; Venegas, C.; Cristòfol, R.; Escames, G.; Acuña-Castroviejo, D.; Sanfeliu, C. Melatonin plus physical exercise are highly neuroprotective in the 3xTg-AD mouse. Neurobiol. Aging 2012, 33, 1124.e13–1124.e29. [Google Scholar] [CrossRef] [Green Version]
- Sturchler-Pierrat, C.; Abramowski, D.; Duke, M.; Wiederhold, K.-H.; Mistl, C.; Rothacher, S.; Ledermann, B.; Bürki, K.; Frey, P.; Paganetti, P.A.; et al. Two amyloid precursor protein transgenic mouse models with Alzheimer disease-like pathology. Proc. Natl. Acad. Sci. USA 1997, 94, 13287–13292. [Google Scholar] [CrossRef] [Green Version]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef] [PubMed]
- Baeta-Corral, R.; Giménez-Llort, L. Bizarre behaviors and risk assessment in 3xTg-AD mice at early stages of the disease. Behav. Brain Res. 2014, 258, 97–105. [Google Scholar] [CrossRef]
- Torres-Lista, V.; Parrado-Fernández, C.; Alvarez-Montón, I.; Frontiñán-Rubio, J.; Durán-Prado, M.; Peinado, J.R.; Johansson, B.; Alcaín, F.J.; Giménez-Llort, L. Neophobia, NQO1 and SIRT1 as premorbid and prodromal indicators of AD in 3xTg-AD mice. Behav. Brain Res. 2014, 271, 140–146. [Google Scholar] [CrossRef]
- Chen, Y.; Liang, Z.; Blanchard, J.; Dai, C.-L.; Sun, S.; Lee, M.H.; Grundke-Iqbal, I.; Iqbal, K.; Liu, F.; Gong, C.-X. A Non-transgenic Mouse Model (icv-STZ Mouse) of Alzheimer’s Disease: Similarities to and Differences from the Transgenic Model (3xTg-AD Mouse). Mol. Neurobiol. 2013, 47, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Bernier, G.; Nardini, E.; Hogan, R.; Flamier, A. Alzheimer’s disease: A tale of two diseases? Neural Regen. Res. 2021, 16, 1958–1964. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, R.; Lewis, T.; Strazielle, C.; Kim, H.; Fukuchi, K. Transgenic mice expressing the βAPP695SWE mutation: Effects on exploratory activity, anxiety, and motor coordination. Brain Res. 2003, 977, 38–45. [Google Scholar] [CrossRef]
- Sinforiani, E.; Citterio, A.; Zucchella, C.; Bono, G.; Corbetta, S.; Merlo, P.; Mauri, M. Impact of Gender Differences on the Outcome of Alzheimer’s Disease. Dement. Geriatr. Cogn. Disord. 2010, 30, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Mielke, M.; Vemuri, P.; Rocca, W. Clinical epidemiology of Alzheimer’s disease: Assessing sex and gender differences. Clin. Epidemiol. 2014, 6, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Mazure, C.M.; Swendsen, J. Sex differences in Alzheimer’s disease and other dementias. Lancet Neurol. 2016, 15, 451–452. [Google Scholar] [CrossRef] [Green Version]
- Pugh, P.L.; Richardson, J.C.; Bate, S.T.; Upton, N.; Sunter, D. Non-cognitive behaviours in an APP/PS1 transgenic model of Alzheimer’s disease. Behav. Brain Res. 2007, 178, 18–28. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Blázquez, G.; Cañete, T.; Rosa, R.; Vivó, M.; Oddo, S.; Navarro, X.; LaFerla, F.M.; Johansson, B.; Tobena, A.; et al. Modeling neuropsychiatric symptoms of Alzheimer’s disease dementia in 3xTg-AD mice. In Alzheimer’s Disease: New Advances; Iqbal, K., Winblad, B., Avila, J., Eds.; Medimond SRL: Pianoro, Italy, 2006; pp. 513–516. [Google Scholar]
- Lát, J. The analysis of habituation. Acta Neurobiol. Exp. 1973, 33, 771–789. [Google Scholar]
- King, D.L.; Arendash, G.W. Behavioral characterization of the Tg2576 transgenic model of Alzheimer’s disease through 19 months. Physiol. Behav. 2002, 75, 627–642. [Google Scholar] [CrossRef]
- Dumont, M.; Strazielle, C.; Staufenbiel, M.; Lalonde, R. Spatial learning and exploration of environmental stimuli in 24-month-old female APP23 transgenic mice with the Swedish mutation. Brain Res. 2004, 1024, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Llort, L.; Ferre, S.; Martínez, E. Effects of the systemic administration of kainic acid and NMDA on exploratory activity in rats. Pharmacol. Biochem. Behav. 1995, 51, 205–210. [Google Scholar] [CrossRef]
- Callisaya, M.L.; Beare, R.; Phan, T.G.; Chen, J.; Srikanth, V.K. Global and Regional Associations of Smaller Cerebral Gray and White Matter Volumes with Gait in Older People. PLoS ONE 2014, 9, e84909. [Google Scholar] [CrossRef] [PubMed]
- Studenski, S.; Perera, S.; Patel, K.; Rosano, C.; Faulkner, K.; Inzitari, M.; Brach, J.; Chandler, J.; Cawthon, P.; Connor, E.B.; et al. Gait speed and survival in older adults. JAMA 2011, 305, 50–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, A.; Fulk, G.D.; Beets, M.W.; Herter, T.M.; Fritz, S.L. Self-Selected Walking Speed Is Predictive of Daily Ambulatory Activity in Older Adults. J. Aging Phys. Act. 2016, 24, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artaud, F.; Singh-Manoux, A.; Dugravot, A.; Tzourio, C.; Elbaz, A. Decline in Fast Gait Speed as a Predictor of Disability in Older Adults. J. Am. Geriatr. Soc. 2015, 63, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Sabia, S.; Dumurgier, J.; Tavernier, B.; Head, J.; Tzourio, C.; Elbaz, A. Change in Fast Walking Speed Preceding Death: Results from a Prospective Longitudinal Cohort Study. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2013, 69, 354–362. [Google Scholar] [CrossRef] [Green Version]
- Callisaya, M.L.; Launay, C.P.; Srikanth, V.K.; Verghese, J.; Allali, G.; Beauchet, O. Cognitive status, fast walking speed and walking speed reserve—the Gait and Alzheimer Interactions Tracking (GAIT) study. GeroScience 2017, 39, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Allali, G.; Annweiler, C.; Blumen, H.M.; Callisaya, M.L.; De Cock, A.-M.; Kressig, R.W.; Srikanth, V.; Steinmetz, J.-P.; Verghese, J.; Beauchet, O. Gait phenotype from mild cognitive impairment to moderate dementia: Results from the GOOD initiative. Eur. J. Neurol. 2016, 23, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Nehlig, A.; Daval, J.-L.; DeBry, G. Caffeine and the central nervous system: Mechanisms of action, biochemical, metabolic and psychostimulant effects. Brain Res. Rev. 1992, 17, 139–170. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Masino, S.A.; Diao, L.; Fernández-Teruel, A.; Tobeña, A.; Halldner, L.; Fredholm, B.B. Mice lacking the adenosine A1 receptor have normal spatial learning and plasticity in the CA1 region of the hippocampus, but they habituate more slowly. Synapse 2005, 57, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, D.L.; Arendash, G.W. Maintained synaptophysin immunoreactivity in Tg2576 transgenic mice during aging: Correlations with cognitive impairment. Brain Res. 2002, 926, 58–68. [Google Scholar] [CrossRef]
- España, J.; Giménez-Llort, L.; Valero, J.; Miñano, A.; Rábano, A.; Rodriguez-Alvarez, J.; LaFerla, F.M.; Saura, C.A. Intraneuronal β-Amyloid Accumulation in the Amygdala Enhances Fear and Anxiety in Alzheimer’s Disease Transgenic Mice. Biol. Psychiatry 2010, 67, 513–521. [Google Scholar] [CrossRef]
- Thompson, R. The comparative effects of anterior and posterior cortical lesions on maze retention. J. Comp. Physiol. Psychol. 1959, 52, 506–508. [Google Scholar] [CrossRef]
- Zhang, W.-N.; Pothuizen, H.; Feldon, J.; Rawlins, J. Dissociation of function within the hippocampus: Effects of dorsal, ventral and complete excitotoxic hippocampal lesions on spatial navigation. Neuroscience 2004, 127, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M.J.; Rawlins, J.N.P. T-maze alternation in the rodent. Nat. Protoc. 2006, 1, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Llort, L.; Schiffmann, S.N.; Shmidt, T.; Canela, L.; Camón, L.; Wassholm, M.; Canals, M.; Terasmaa, A.; Fernández-Teruel, A.; Tobeña, A.; et al. Working memory deficits in transgenic rats overexpressing human adenosine A2A receptors in the brain. Neurobiol. Learn. Mem. 2007, 87, 42–56. [Google Scholar] [CrossRef] [Green Version]
- Guayerbas, N.; Puerto, M.; Ferrández, M.D.; De La Fuente, M. A diet supplemented with thiolic anti-oxidants improves leucocyte function in two strains of prematurely ageing mice. Clin. Exp. Pharmacol. Physiol. 2002, 29, 1009–1014. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; García, Y.; Buccieri, K.; Revilla, S.; Suñol, C.; Cristofol, R.; Sanfeliu, C. Gender-Specific Neuroimmunoendocrine Response to Treadmill Exercise in 3xTg-AD Mice. Int. J. Alzheimer’s Dis. 2010, 2010, 1–17. [Google Scholar] [CrossRef] [Green Version]
- García-Mesa, Y.; López-Ramos, J.C.; Giménez-Llort, L.; Revilla, S.; Guerra, R.; Gruart, A.; LaFerla, F.M.; Cristòfol, R.; Delgado-García, J.M.; Sanfeliu, C. Physical Exercise Protects Against Alzheimer’s Disease in 3xTg-AD Mice. J. Alzheimer’s Dis. 2011, 24, 421–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariza, L.; Giménez-Llort, L.; Cubizolle, A.; Pagès, G.; García-Lareu, B.; Serratrice, N.; Cots, D.; Thwaite, R.; Chillón, M.; Kremer, E.J.; et al. Central Nervous System Delivery of Helper-Dependent Canine Adenovirus Corrects Neuropathology and Behavior in Mucopolysaccharidosis Type VII Mice. Hum. Gene Ther. 2014, 25, 199–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanderson, D.J.; Bannerman, D.M. The role of habituation in hippocampus-dependent spatial working memory tasks: Evidence from GluA1 AMPA receptor subunit knockout mice. Hippocampus 2012, 22, 981–994. [Google Scholar] [CrossRef] [Green Version]
- Tempier, A.; He, J.; Zhu, S.; Zhang, R.; Kong, L.; Tan, Q.; Luo, H.; Kong, J.; Li, X.-M. Quetiapine Modulates Conditioned Anxiety and Alternation Behavior in Alzheimer’s Transgenic Mice. Curr. Alzheimer Res. 2013, 10, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, A.; Mandillo, S.; Oliverio, A.; Mele, A. D1 and D2 Receptor Antagonist Injections in the Prefrontal Cortex Selectively Impair Spatial Learning in Mice. Neuropsychopharmacology 2006, 32, 309–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delotterie, D.; Ruiz, G.; Brocard, J.; Schweitzer, A.; Roucard, C.; Roche, Y.; Suaud-Chagny, M.-F.; Bressand, K.; Andrieux, A. Chronic administration of atypical antipsychotics improves behavioral and synaptic defects of STOP null mice. Psychopharmacology 2010, 208, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Morris, R.G.M. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Rivera-Hernández, G.; Marín-Argany, M.; Sánchez-Quesada, J.L.; Villegas, S. Early intervention in the 3xTg-AD mice with an amyloid β-antibody fragment ameliorates first hallmarks of Alzheimer disease. mAbs 2013, 5, 665–864. [Google Scholar] [CrossRef] [Green Version]
- Baeta-Corral, R.; Giménez-Llort, L. Persistent hyperactivity and distinctive strategy features in the Morris water maze in 3xTg-AD mice at advanced stages of disease. Behav. Neurosci. 2015, 129, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, R.; Dumont, M.; Staufenbiel, M.; Strazielle, C. Neurobehavioral characterization of APP23 transgenic mice with the SHIRPA primary screen. Behav. Brain Res. 2005, 157, 91–98. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Tests and Variables | Statistics | Genotype | Age | G × A | Significance |
|---|---|---|---|---|---|
| Corner test | |||||
| Total visited corners | F (2,52) = 7.392 | - | A *** | - | p = 0.001 |
| F (2,52) = 4.126 | G * | - | - | p = 0.047 | |
| Total numbers of rearings | F (2,52) = 13.374 | - | A *** | - | p = 0.000 |
| Latency of rearing (s) | F (2,52) = 10.210 | - | A *** | - | p = 0.000 |
| Open field test | |||||
| Freezing—Latency of first movement (s) | F (2,52) = 13.362 | - | A *** | - | p = 0.000 |
| F (2,52) = 3.664 | - | - | G × A * | p = 0.032 | |
| Latency to exit the center (s) | F (2,52) = 6.640 | - | A ** | - | p = 0.003 |
| F (2,52) = 11.169 | G ** | - | - | p = 0.002 | |
| Latency of rearing (s) | F (2,52) = 2.905 | - | - | G × A (*) | p = 0.064 |
| F (2,52) = 2.649 | - | A (*) | - | p = 0.080 | |
| Latency of self-grooming (s) | F (2,52) = 5.917 | G * | - | - | p = 0.018 |
| Total horizontal activity (n crossings) | F (2,52) = 12.909 | - | A *** | - | p = 0.000 |
| F (2,52) = 6.043 | G* | - | - | p = 0.017 | |
| “in the center (n crossings) | F (2,52) = 3.044 | - | - | G × A (*) | p = 0.056 |
| “in the periphery (n crossings) | F (2,52) = 11.655 | - | A *** | - | p = 0.000 |
| F (2,52) = 9.404 | G ** | - | - | p = 0.003 | |
| Total vertical activity (n of rearings) | F (2,52) = 18.966 | - | A *** | - | p = 0.000 |
| Context and object recognition tests | |||||
| Latency to enter into the periphery | F(2,52) = 13.448 | - | A *** | - | p = 0.000 |
| Total horizontal activity | |||||
| “in the center (n of crossings) | F(2,52) = 3.325 | - | A * | - | p = 0.044 |
| “in the periphery (n of crossings) | F(2,52) = 3.339 | - | A * | - | p = 0.043 |
| Time exploring new object (s) | F(2,52) = 3.746 | - | A * | - | p = 0.030 |
| T-maze | |||||
| Latency to cross the intersection (s) | F(2,51) = 27.978 | G *** | - | - | p = 0.000 |
| Total time exploring both arms (s) | F(2,51) = 6.005 | - | A ** | - | p = 0.005 |
| F(2,51) = 4.107 | G * | - | - | p = 0.048 | |
| Total time to complete the test (s) | F(2,51) = 19.484 | G *** | - | - | p = 0.000 |
| Total number of errors (n) | F(2,51) = 3.139 | - | A (*) | - | p = 0.052 |
| Total vertical activity (n of rearings) | F(2,51) = 9.700 | G ** | - | - | p = 0.003 |
| F(2,51) = 3.631 | - | - | G × A * | p = 0.034 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giménez-Llort, L.; Marin-Pardo, D.; Marazuela, P.; Hernández-Guillamón, M. Survival Bias and Crosstalk between Chronological and Behavioral Age: Age- and Genotype-Sensitivity Tests Define Behavioral Signatures in Middle-Aged, Old, and Long-Lived Mice with Normal and AD-Associated Aging. Biomedicines 2021, 9, 636. https://doi.org/10.3390/biomedicines9060636
Giménez-Llort L, Marin-Pardo D, Marazuela P, Hernández-Guillamón M. Survival Bias and Crosstalk between Chronological and Behavioral Age: Age- and Genotype-Sensitivity Tests Define Behavioral Signatures in Middle-Aged, Old, and Long-Lived Mice with Normal and AD-Associated Aging. Biomedicines. 2021; 9(6):636. https://doi.org/10.3390/biomedicines9060636
Chicago/Turabian StyleGiménez-Llort, Lydia, Daniela Marin-Pardo, Paula Marazuela, and Mar Hernández-Guillamón. 2021. "Survival Bias and Crosstalk between Chronological and Behavioral Age: Age- and Genotype-Sensitivity Tests Define Behavioral Signatures in Middle-Aged, Old, and Long-Lived Mice with Normal and AD-Associated Aging" Biomedicines 9, no. 6: 636. https://doi.org/10.3390/biomedicines9060636