
Targeted SMN Exon Skipping: A Useful Control to Assess In Vitro and In Vivo Splice-Switching Studies

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
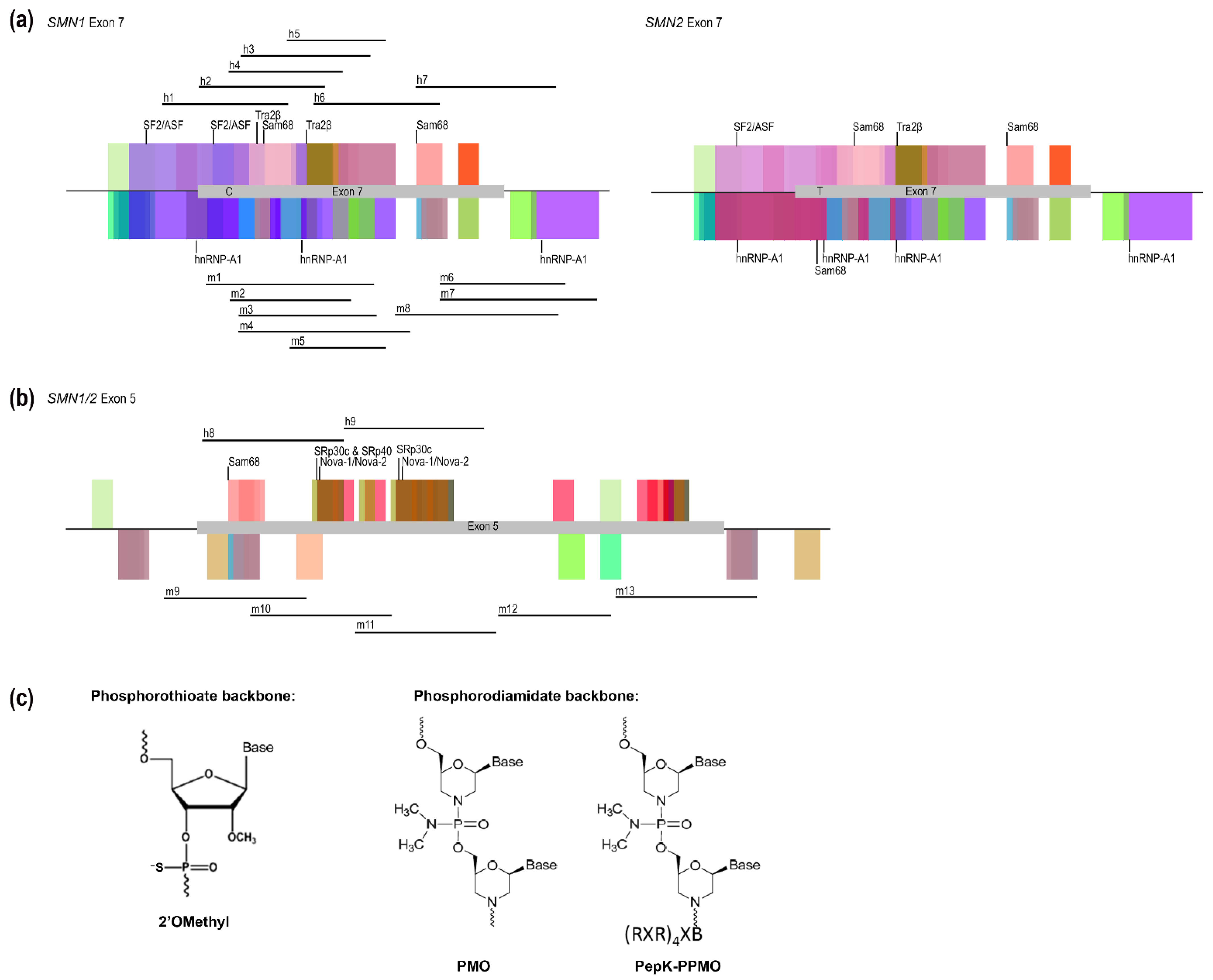
2.1. AO Design
2.2. Cell Plating and 2′-O-Methyl Phosphorothioate AO Transfection
2.3. Culture and Differentiation of SH-SY5Y Cells
2.4. Transfection of Phosphorodiamidate Morpholino Oligomers and Peptide Conjugated PMOs
2.5. RNA Extraction, Polymerase Chain Reaction, Visualisation and Analysis
2.6. In Silico Analysis of SMN and Smn Splice Site Scores
2.7. In Vivo Evaluation of a Peptide-Conjugated PMO
3. Results
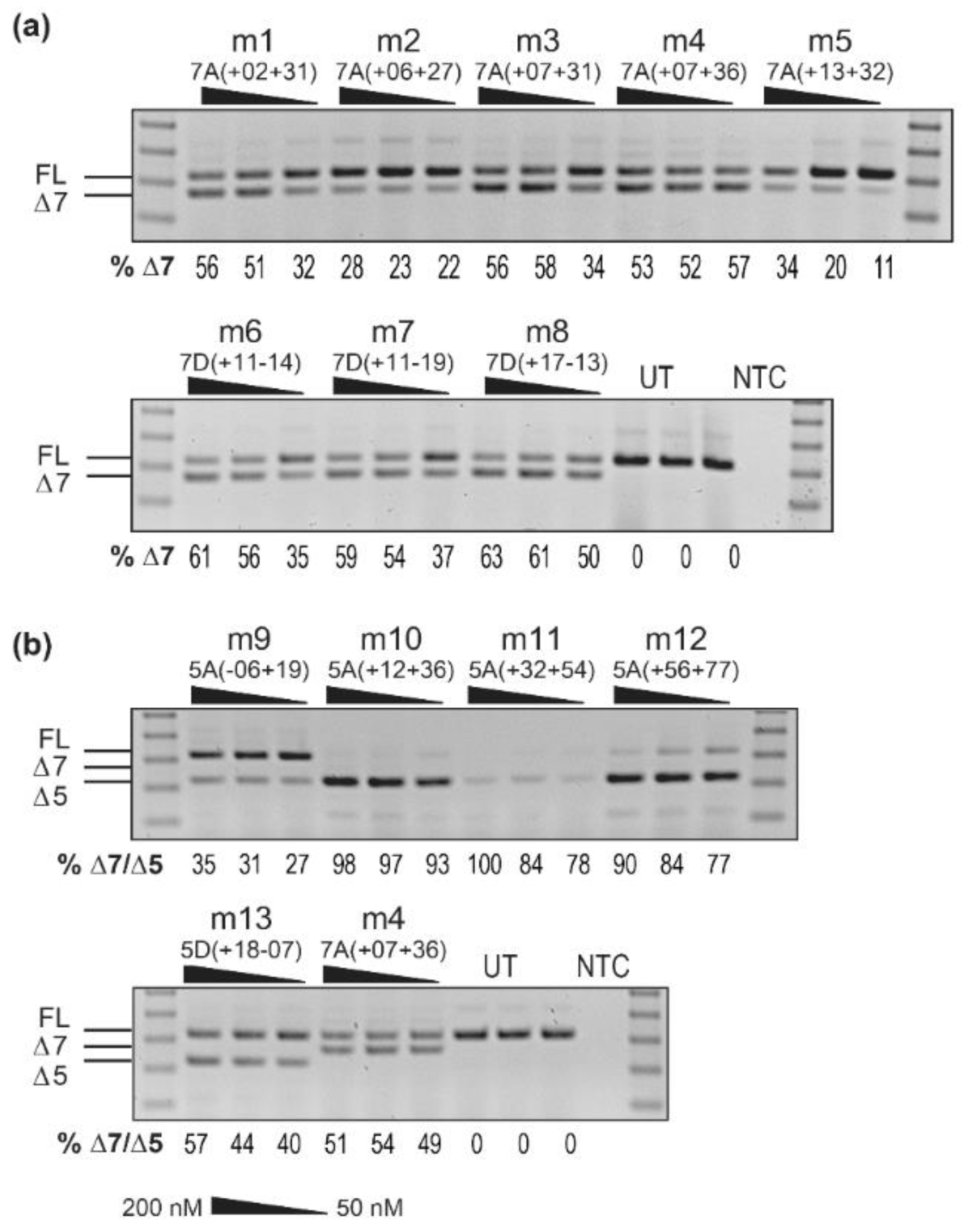
3.1. Screening of 2′-O-Methyl AOs Targeting SMN Exons 5 and 7 in Normal Human Fibroblasts
3.2. In Silico Analysis of Human and Mouse Exon Splice Site Scores
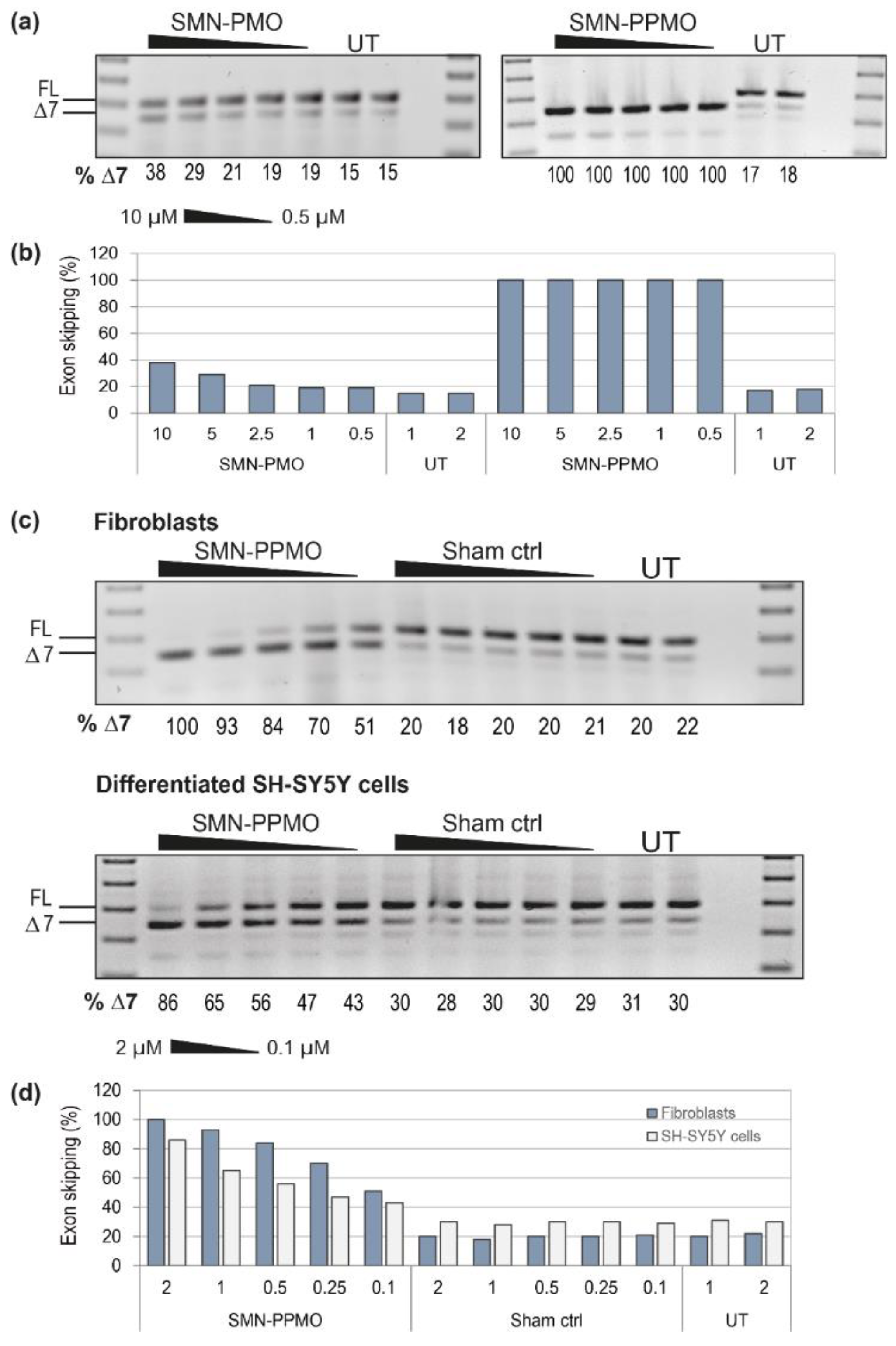
3.3. Using the SMN Exon Skipping AO as a Positive Control for Optimising AO Chemistry and Delivery Across Cell Lines
3.4. Screening of 2′-O-Methyl AOs Targeting Smn Exon 7 in Mdx Myoblasts
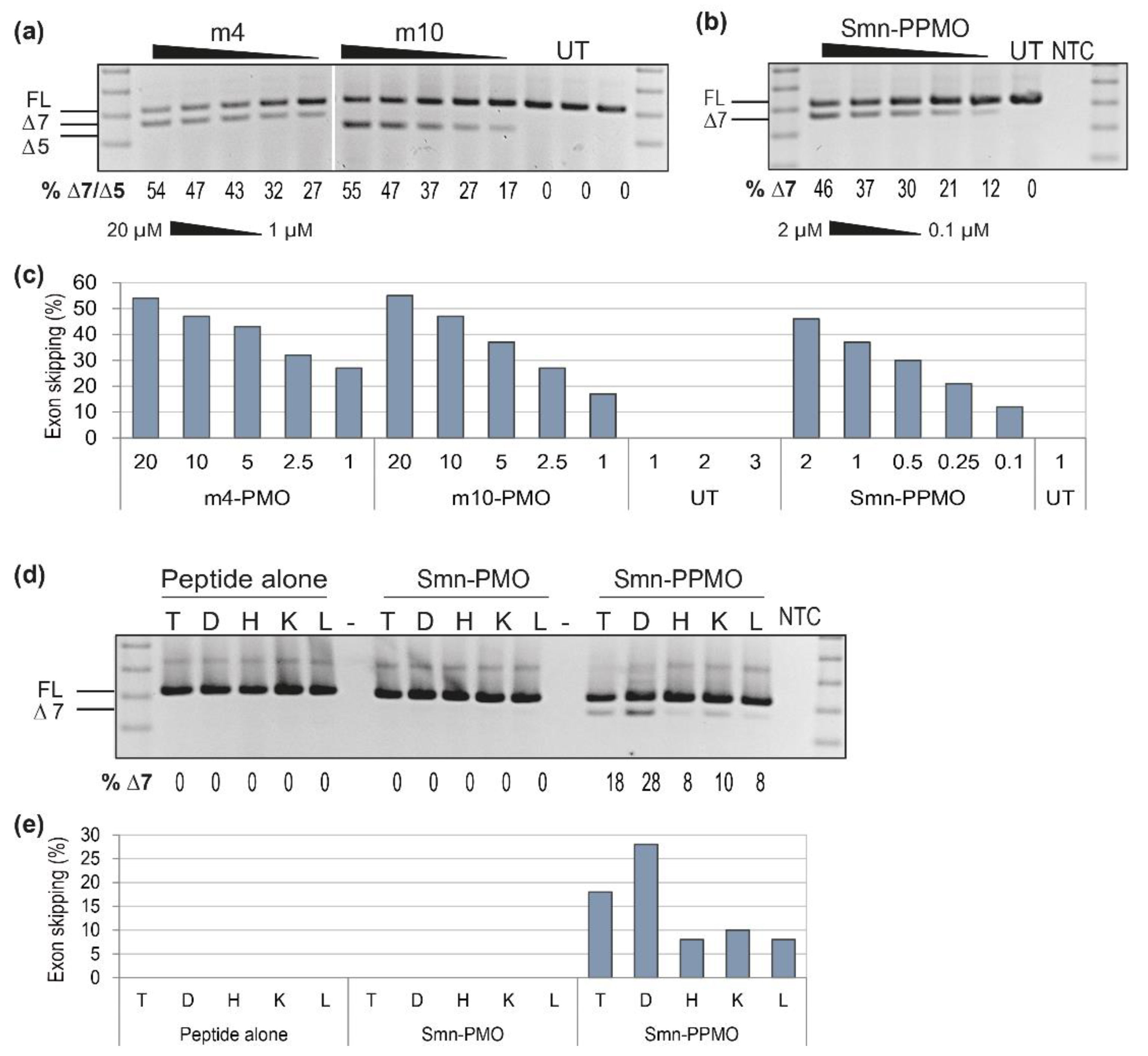
3.5. Using the Mouse Smn AO as a Positive Control for Optimizing AO Chemistry and In Vivo Delivery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AO | antisense oligonucleotide |
| SMN | survival motor neuron |
| SMA | spinal muscular atrophy |
| PMO | phosphorodiamidate morpholino oligomer |
| PPMO | peptide-conjugated phosphorodiamidate morpholino oligomer |
References
- Uehara, T.C.J.; Choong, M.; Nakamori, H.; Hayakawa, K.; Nishiyama, Y.; Kasahara, K.; Baba, T.; Nagata, T.; Yokota, H.; Tsuda, S.; et al. Amido-bridged nucleic acid (AmNA)-modified antisense oligonucleotides targeting alpha-synuclein as a novel therapy for Parkinson’s disease. Sci. Rep. 2019, 9, 7567. [Google Scholar] [CrossRef]
- Nizzardo, M.; Simone, C.; Rizzo, F.; Ulzi, G.; Ramirez, A.; Rizzuti, M.; Bordoni, A.; Bucchia, M.; Gatti, S.; Bresolin, N.; et al. Morpholino-mediated SOD1 reduction ameliorates an amyotrophic lateral sclerosis disease phenotype. Sci. Rep. 2016, 6, 21301. [Google Scholar] [CrossRef] [Green Version]
- Aung-Htut, M.T.; Comerford, I.; Johnsen, R.; Foyle, K.; Fletcher, S.; Wilton, S.D. Reduction of integrin alpha 4 activity through splice modulating antisense oligonucleotides. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Flynn, L.L.; Mitrpant, C.; Pitout, I.L.; Fletcher, S.; Wilton, S.D. Antisense Oligonucleotide-Mediated Terminal Intron Retention of the SMN2 Transcript. Mol. Ther. Nucleic Acids 2018, 11, 91–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabaglio, T.; Low, D.H.; Teo, W.K.L.; Goy, P.A.; Cywoniuk, P.; Wollmann, H.; Ho, J.; Tan, D.; Aw, J.; Pavesi, A.; et al. MBNL1 alternative splicing isoforms play opposing roles in cancer. Life Sci. Alliance 2018, 1, e201800157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilton, S.D.; Fall, A.M.; Harding, P.L.; McClorey, G.; Coleman, C.; Fletcher, S. Antisense Oligonucleotide-induced Exon Skipping Across the Human Dystrophin Gene Transcript. Mol. Ther. 2007, 15, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Pitout, I.; Flynn, L.L.; Wilton, S.D.; Fletcher, S. Antisense-mediated splice intervention to treat human disease: The odyssey continues. F1000Research 2019, 8, 710. [Google Scholar] [CrossRef] [Green Version]
- Aung-Htut, M.T.; McIntosh, C.S.; West, K.A.; Fletcher, S.; Wilton, S.D.; Ham, K. In Vitro Validation of Phosphorodiamidate Morpholino Oligomers. Molecules 2019, 24, 2922. [Google Scholar] [CrossRef] [Green Version]
- Aung-Htut, M.T.; McIntosh, C.S.; Ham, K.A.; Pitout, I.L.; Flynn, L.L.; Greer, K.; Fletcher, S.; Wilton, S.D. Systematic Approach to Developing Splice Modulating Antisense Oligonucleotides. Int. J. Mol. Sci. 2019, 20, 5030. [Google Scholar] [CrossRef] [Green Version]
- Errington, S.J.; Mann, C.J.; Fletcher, S.; Wilton, S.D. Target selection for antisense oligonucleotide induced exon skipping in the dystrophin gene. J. Gene Med. 2003, 5, 518–527. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Larcher, L.M.; Ma, L.; Veedu, R.N. Systematic Screening of Commonly Used Commercial Transfection Reagents towards Efficient Transfection of Single-Stranded Oligonucleotides. Molecules 2018, 23, 2564. [Google Scholar] [CrossRef] [Green Version]
- Linnane, E.; Davey, P.; Zhang, P.; Puri, S.; Edbrooke, M.; Chiarparin, E.; Revenko, A.S.; MacLeod, A.R.; Norman, J.C.; Ross, S.J. Differential uptake, kinetics and mechanisms of intracellular trafficking of next-generation antisense oligonucleotides across human cancer cell lines. Nucleic Acids Res. 2019, 47, 4375–4392. [Google Scholar] [CrossRef]
- Wang, S.; Sun, H.; Tanowitz, M.; Liang, X.-H.; Crooke, S.T. Intra-endosomal trafficking mediated by lysobisphosphatidic acid contributes to intracellular release of phosphorothioate-modified antisense oligonucleotides. Nucleic Acids Res. 2017, 45, 5309–5322. [Google Scholar] [CrossRef]
- Flynn, L.L.; Ruohan, L.; Aung-Htut, M.T.; Pitout, I.L.; Cooper, J.; Hubbard, A.; Griffiths, L. Interaction of modified oligonucleotides with nuclear proteins, formation of novel nuclear structures and sequence-independent effects on RNA processing. bioRxiv 2019. [Google Scholar] [CrossRef]
- Shen, W.; De Hoyos, C.L.; Sun, H.; Vickers, T.A.; Liang, X.-H.; Crooke, S.T. Acute hepatotoxicity of 2′ fluoro-modified 5–10–5 gapmer phosphorothioate oligonucleotides in mice correlates with intracellular protein binding and the loss of DBHS proteins. Nucleic Acids Res. 2018, 46, 2204–2217. [Google Scholar] [CrossRef] [Green Version]
- Crooke, S.T.; Baker, B.F.; Witztum, J.L.; Kwoh, T.J.; Pham, N.C.; Salgado, N.; McEvoy, B.W.; Cheng, W.; Hughes, S.G.; Bhanot, S.; et al. The Effects of 2′-O-Methoxyethyl Containing Antisense Oligonucleotides on Platelets in Human Clinical Trials. Nucleic Acid Ther. 2017, 27, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Adams, A.M.; Harding, P.L.; Iversen, P.L.; Coleman, C.; Fletcher, S.; Wilton, S.D. Antisense oligonucleotide induced exon skipping and the dystrophin gene transcript: Cocktails and chemistries. BMC Mol. Biol. 2007, 8, 57–58. [Google Scholar] [CrossRef] [Green Version]
- Tsoumpra, M.K.; Fukumoto, S.; Matsumoto, T.; Takeda, S.; Wood, M.J.; Aoki, Y. Peptide-conjugate antisense based splice-correction for Duchenne muscular dystrophy and other neuromuscular diseases. EBioMedicine 2019, 45, 630–645. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, K.; Milech, N.; Juraja, S.M.; Cunningham, P.T.; Stone, S.R.; Francis, R.W.; Anastasas, M.; Hall, C.M.; Heinrich, T.; Bogdawa, H.M.; et al. A platform for discovery of functional cell-penetrating peptides for efficient multi-cargo intracellular delivery. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M.; Hazell, G.; Shabanpoor, F.; Saleh, A.F.; Bowerman, M.; Sleigh, J.N.; Meijboom, K.E.; Zhou, H.; Muntoni, F.; Talbot, K.; et al. Systemic peptide-mediated oligonucleotide therapy improves long-term survival in spinal muscular atrophy. Proc. Natl. Acad. Sci. USA 2016, 113, 10962–10967. [Google Scholar] [CrossRef] [Green Version]
- Guidotti, G.; Brambilla, L.; Rossi, D. Cell-Penetrating Peptides: From Basic Research to Clinics. Trends Pharmacol. Sci. 2017, 38, 406–424. [Google Scholar] [CrossRef]
- Singh, N.K.; Singh, N.N.; Androphy, E.J.; Singh, R.N. Splicing of a Critical Exon of Human Survival Motor Neuron Is Regulated by a Unique Silencer Element Located in the Last Intron. Mol. Cell. Biol. 2006, 26, 1333–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, E.Y.; Miller, M.R.; Robbins, K.L.; Lombardi, A.M.; Atkinson, A.K.; Brehm, A.J.; Lorson, C.L. Morpholino antisense oligonucleotides targeting intronic repressor Element1 improve phenotype in SMA mouse models. Hum. Mol. Genet. 2014, 23, 4832–4845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkel, R.S.; Chiriboga, C.A.; Vajsar, J.; Day, J.W.; Montes, J.; De Vivo, D.C.; Yamashita, M.; Rigo, F.; Hung, G.; Schneider, E.; et al. Treatment of infantile-onset spinal muscular atrophy with nusinersen: A phase 2, open-label, dose-escalation study. Lancet 2016, 388, 3017–3026. [Google Scholar] [CrossRef]
- Sahashi, K.; Hua, Y.; Ling, K.K.; Hung, G.; Rigo, F.; Horev, G.; Katsuno, M.; Sobue, G.; Ko, C.-P.; Bennett, C.F.; et al. TSUNAMI: An antisense method to phenocopy splicing-associated diseases in animals. Genes Dev. 2012, 26, 1874–1884. [Google Scholar] [CrossRef] [Green Version]
- Mitrpant, C.; Adams, A.M.; Meloni, P.L.; Muntoni, F.; Fletcher, S.; Wilton, S.D. Rational Design of Antisense Oligomers to Induce Dystrophin Exon Skipping. Mol. Ther. 2009, 17, 1418–1426. [Google Scholar] [CrossRef]
- Bramsen, J.B.; Ekjems, J. Development of Therapeutic-Grade Small Interfering RNAs by Chemical Engineering. Front. Genet. 2012, 3, 154. [Google Scholar] [CrossRef] [Green Version]
- Warren, T.K.; Shurtleff, A.C.; Bavari, S. Advanced morpholino oligomers: A novel approach to antiviral therapy. Antivir. Res. 2012, 94, 80–88. [Google Scholar] [CrossRef]
- Piva, F.; Giulietti, M.; Burini, A.B.; Principato, G. SpliceAid 2: A database of human splicing factors expression data and RNA target motifs. Hum. Mutat. 2011, 33, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Yeo, E.; Hoon, S.; Venkatesh, B.; Burge, C.B. Variation in sequence and organization of splicing regulatory elements in vertebrate genes. Proc. Natl. Acad. Sci. USA 2004, 101, 15700–15705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, C.J.; Honeyman, K.; McClorey, G.; Fletcher, S.; Wilton, S.D. Improved antisense oligonucleotide induced exon skipping in themdx mouse model of muscular dystrophy. J. Gene Med. 2002, 4, 644–654. [Google Scholar] [CrossRef]
- Jearawiriyapaisarn, N.; Moulton, H.M.; Buckley, B.; Roberts, J.; Sazani, P.; Fucharoen, S.; Iversen, P.L.; Kole, R. Sustained Dystrophin Expression Induced by Peptide-conjugated Morpholino Oligomers in the Muscles of mdx Mice. Mol. Ther. 2008, 16, 1624–1629. [Google Scholar] [CrossRef] [PubMed]
- Yeo, G.; Burge, C.B. Maximum entropy modeling of short sequence motifs with applications to RNA splicing signals. In Proceedings of the Seventh Annual International Conference on Computational Molecular Biology RECOMB ’03, Berlin, Germany, 10–13 April 2003; Volume 11, pp. 322–331. [Google Scholar] [CrossRef]
- Mohaghegh, P.; Rodrigues, N.R.; Owen, N.; Ponting, C.P.; Le, T.T.; Burghes, A.H.; Davies, K.E. Analysis of mutations in the tudor domain of the survival motor neuron protein SMN. Eur. J. Hum. Genet. 1999, 7, 519–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moles-Fernández, A.; Duran-Lozano, L.; Montalban, G.; Bonache, S.; López-Perolio, I.; Menéndez, M.; Santamariña, M.; Behar, R.; Blanco, A.; Carrasco, E.; et al. Computational Tools for Splicing Defect Prediction in Breast/Ovarian Cancer Genes: How Efficient Are They at Predicting RNA Alterations? Front. Genet. 2018, 9, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betts, C.; Saleh, A.F.; Arzumanov, A.A.; Hammond, S.M.; Godfrey, C.; Coursindel, T.; Gait, M.J.; Wood, M.J. Pip6-PMO, A New Generation of Peptide-oligonucleotide Conjugates With Improved Cardiac Exon Skipping Activity for DMD Treatment. Mol. Ther. Nucleic Acids 2012, 1, e38. [Google Scholar] [CrossRef] [PubMed]
- Boisguérin, P.; Deshayes, S.; Gait, M.J.; O’Donovan, L.; Godfrey, C.; Betts, C.A.; Wood, M.J.; Lebleu, B. Delivery of therapeutic oligonucleotides with cell penetrating peptides. Adv. Drug Deliv. Rev. 2015, 87, 52–67. [Google Scholar] [CrossRef]
- Jing, N.; Li, Y.; Xiong, W.; Sha, W.; Jing, L.; Tweardy, D.J. G-Quartet Oligonucleotides. Cancer Res. 2004, 64, 6603–6609. [Google Scholar] [CrossRef] [Green Version]
- Moulton, J.D.; Yan, Y. Using Morpholinos to Control Gene Expression. Curr. Protoc. Mol. Biol. 2008, 83, 26.8.1–26.8.29. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species and Exon | AO Name/Number | Coordinates | Sequence 5′-3′ | Length (bp) | Tm (ᵒC) |
|---|---|---|---|---|---|
| Human Exon 7 | h1 h2 h3 h4 h5 h6 h7 | SMN H7A(− 08 + 17) SMN H7A(+ 01 + 25) 1 SMN H7A(+ 07 + 31) 1 SMN H7A(+ 06 + 27) 1 SMN H7A(+ 13 + 32) 1 SMN H7A(+ 22 + 46) 1 SMN H7D(+ 17 − 13) | UGA UUU UGU CUA AAA CCC UGU AAG G CUU CUU UUU GAU UUU GUC UAA AAC C ACC UUC CUU CUU UUU GAU UUU GUC U UCC UUC UUU UUG AUU UUG UCU G CAC CUU CCU UCU UUU UGA UU UUA AGG AAU GUG AGC ACC UUC CUU C CUG GCA GAC UUA CUC CUU AAU UUA AGG AAU | 25 25 25 22 20 25 30 | 66.4 58.6 64.7 59.5 59.2 70.5 70.2 |
| Human Exon 5 | h8 h9 | SMN H5A(+ 04 + 28) SMN H5A(+ 29 + 53) | GUG GUG GGC CAU UGA AUU UUA GAC C UGG GGU GGU GGU GGU GGC GGU GGC G | 25 25 | 71.1 87.0 |
| Mouse Exon 7 | m1 m2 m3 m4 m5 m6 m7 m8 | Smn M7A(+ 02 + 31) Smn M7A(+ 06 + 27) Smn M7A(+ 07 + 31) Smn M7A(+ 07 + 36) 2 Smn M7A(+ 13 + 32) Smn M7D(+ 11 − 14) Smn M7D(+ 11 − 19) Smn M7D(+ 17 − 13) | ACU UUC CUU CUU UUU UAU UUU GUC UGA AAC UCC UUC UUU UUU AUU UUG UCU G ACU UUC CUU CUU UUU UAU UUU GUC U UGA GCA CUU UCC UUC UUU UUU AUU UUG UCU CAC UUU CCU UCU UUU UUA UU AAU GAC AGA CUU ACU UCU UAA UUU G UUU AAA AUG ACA GAC UUA CUU CUU AAU UUG AUG ACA GAC UUA CUU CUU AAU UUG UAU GUG | 30 22 25 30 20 25 30 30 | 63.5 56.9 59.8 67.2 52.6 60.7 61.9 65.9 |
| Mouse Exon 5 | m9 m10 m11 m12 m13 | Smn M5A(− 06 + 19) Smn M5A(+ 12 + 36) Smn M5A(+ 32 + 54) Smn M5A(+ 56 + 77) Smn M5A(+ 18 − 07) | CGU UGA AUU UUA GAC CUG GCU AUA A AGG CGG CGG CGG CGG GCC GUU GAA U AAG GGG GGA GGG GGU AGU GGA GGC G AAC GGG GGC AUC CAG CAC GGC AG UAC UUA CUG GUG GUC CUG AAG GGA A | 25 25 25 23 25 | 65.8 86.8 85.9 81.6 75.7 |
| Transcript Sizes | Primer | Primer Sequence | Temperature Profile |
|---|---|---|---|
| SMN FL 404 bp Δ5 308 bp Δ7 350 bp | Exon 4 Fwd Exon 8 Rev | AGGTCTCCTGGAAATAAATCAG TGGTGTCATTTAGTGCTGCTCT | 55 °C 30 min 94 °C 2 min 25 cycles: 94 °C 40 s 56 °C 1 min 68 °C 1 min |
| Smn FL 430 bp Δ5 334 bp Δ7 380 bp | Exon 4 Fwd Exon 8 Rev | GAAAGTCAAGTTTCCACAGACG CACCCCATCTCCTGAGACAGAGC | 55 °C 30 min 94 °C 2 min 28 cycles: 94 °C 40 s 60 °C 1 min 68 °C 1 min |
| Species | Exon | Splice Site | Splice Site Sequence | MES Score |
|---|---|---|---|---|
| Human | 5 | Acceptor Donor | ctttgaaatattccttatag CCA CCAgtaagt | 5.44 9.09 |
| 7 | Acceptor Donor | ttcctttattttccttacagGGT GGAgtaagt | 10.92 8.57 | |
| Mouse | 5 | Acceptor Donor | cttggaaatattctttatagCCA CCAgtaagt | 3.29 9.09 |
| 7 | Acceptor Donor | tttatatgctctctttacagGGT GAAgtaagt | 11.13 9.82 |
| Species | Coordinates | Sequence 5′-3′ |
|---|---|---|
| Human | SMN H7A(+ 07 + 31) SMN H7A(+ 13 + 32) | ACC UUC CUU CUU UUU GAU UUU GUC U CAC CUU CCU UCU UUU UGA UU |
| Mouse | Smn M7A(+ 07 + 31) Smn M5A(+ 12 + 36) | ACU UUC CUU CUU UUU UAU UUU GUC U AGG CGG CGG CGG CGG GCC GUU GAA U |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flynn, L.L.; Mitrpant, C.; Adams, A.; Pitout, I.L.; Stirnweiss, A.; Fletcher, S.; Wilton, S.D. Targeted SMN Exon Skipping: A Useful Control to Assess In Vitro and In Vivo Splice-Switching Studies. Biomedicines 2021, 9, 552. https://doi.org/10.3390/biomedicines9050552
Flynn LL, Mitrpant C, Adams A, Pitout IL, Stirnweiss A, Fletcher S, Wilton SD. Targeted SMN Exon Skipping: A Useful Control to Assess In Vitro and In Vivo Splice-Switching Studies. Biomedicines. 2021; 9(5):552. https://doi.org/10.3390/biomedicines9050552
Chicago/Turabian StyleFlynn, Loren L., Chalermchai Mitrpant, Abbie Adams, Ianthe L. Pitout, Anja Stirnweiss, Sue Fletcher, and Steve D. Wilton. 2021. "Targeted SMN Exon Skipping: A Useful Control to Assess In Vitro and In Vivo Splice-Switching Studies" Biomedicines 9, no. 5: 552. https://doi.org/10.3390/biomedicines9050552