
Evidence of Neurovascular Un-Coupling in Mild Alzheimer’s Disease through Multimodal EEG-fNIRS and Multivariate Analysis of Resting-State Data

 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Design
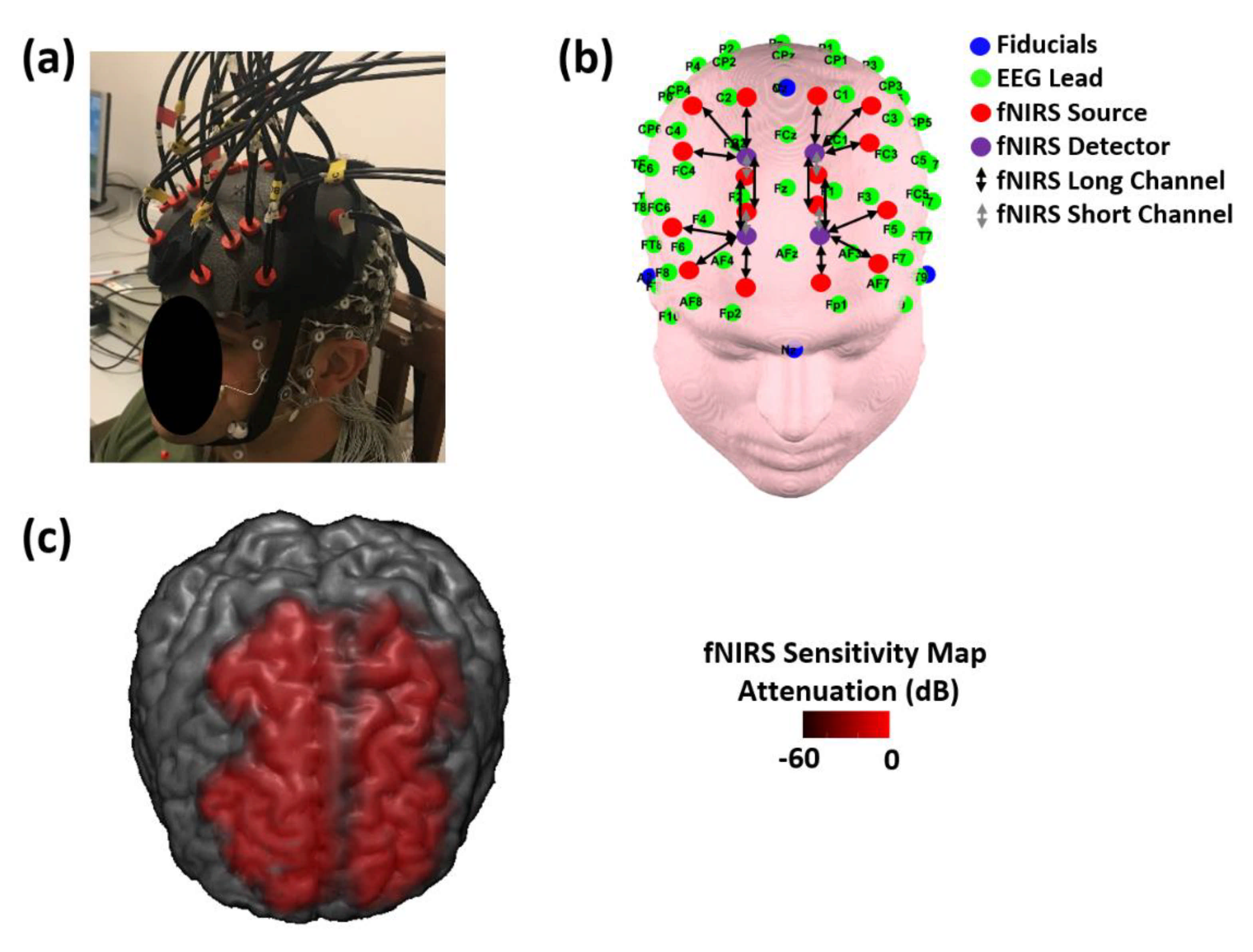
2.3. EEG Instrumentation and Measurements
2.4. FNIRS Instrumentation and Measurements

2.5. EEG Pre-Processing
2.6. FNIRS Pre-Processing
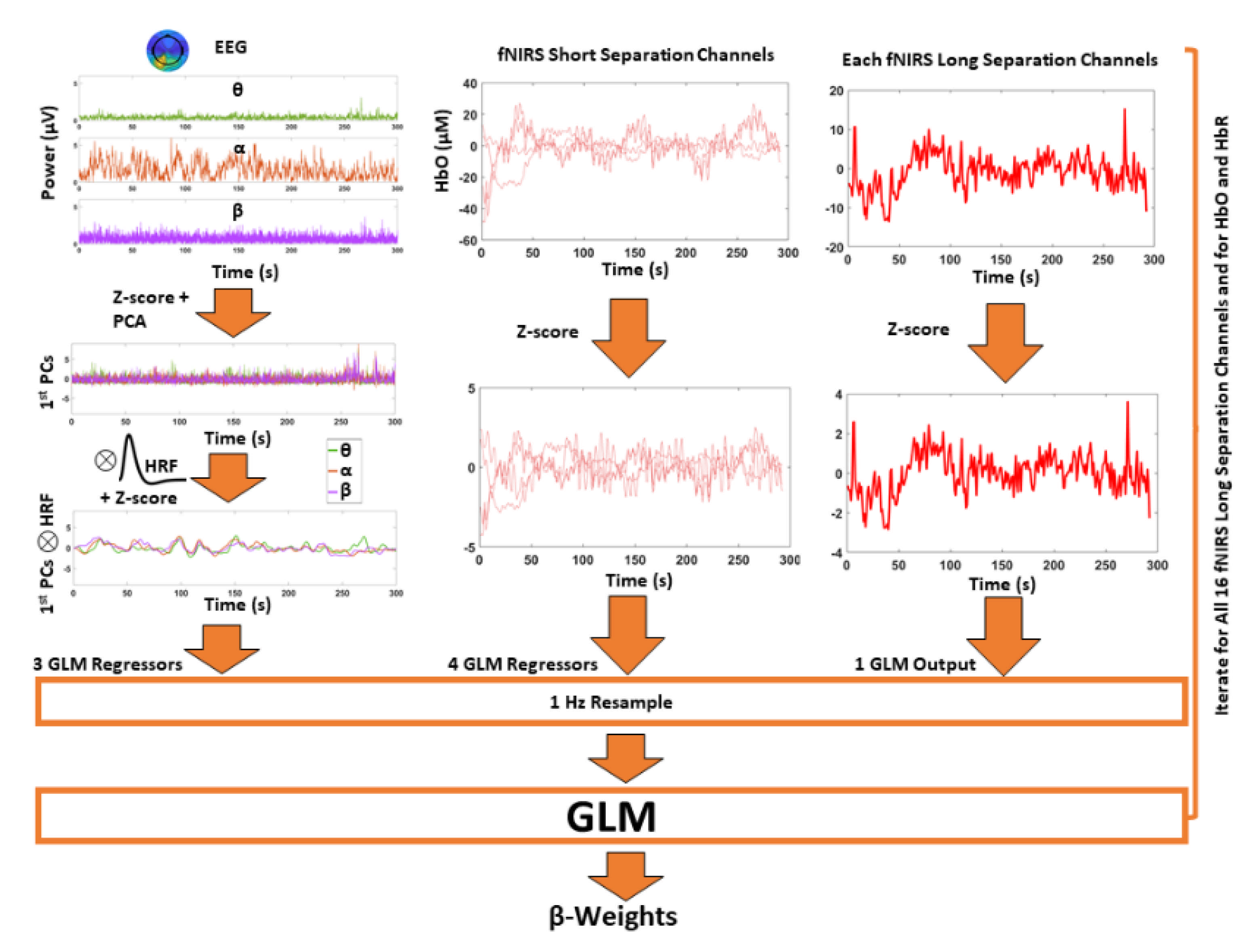
2.7. Neurovascular Coupling Estimation
2.8. Statistical Inference and Multivariate Data-Driven Analysis
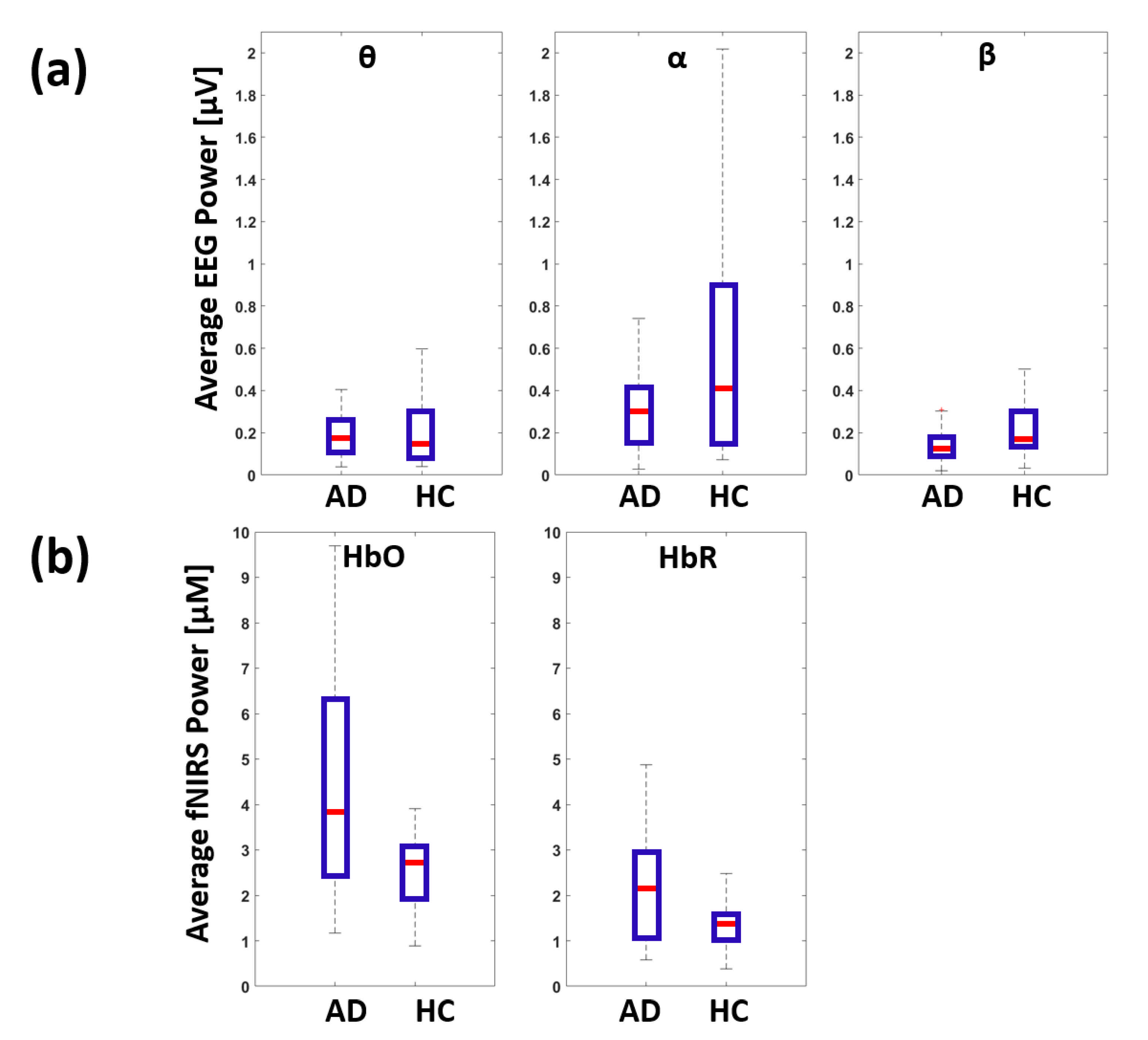
3. Results
4. Discussion
4.1. Identification of EEG-fNIRS De-Coupling in AD
4.2. Multivariate Exploitation of EEG-fNIRS De-Coupling in AD
4.3. Study Limitations
4.4. Study Significance
4.5. Future Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Abbreviation | Meaning |
| AD | Alzheimer’s Disease |
| AUC | Area Under the Curve |
| BOLD | Blood Oxygen Level-Dependent |
| CBF | Cerebral Blood Flow |
| CMRO2 | Cerebral Metabolic Rate of Oxygen |
| CV | Cross-Validation |
| CVR | Cerebrovascular Reactivity |
| DPF | Differential Pathlength Factor |
| EEG | Electroencephalography |
| fMRI | functional Magnetic Resonance Imaging |
| fNIRS | functional Near-Infrared Spectroscopy |
| GLM | General Linear Model |
| HbO | Oxy-hemoglobin |
| HbR | Deoxy-hemoglobin |
| HC | Healthy Controls |
| HRF | Hemodynamic Response Function |
| ICA | Independent Component Analysis |
| MMSE | Mini Mental State Examination |
| NC | Neurovascular Coupling |
| OEF | Oxygen Extraction Fraction |
| PCA | Principal Component Analysis |
| PMT | Photomultiplier Tube |
| ROC | Receiver Operating Characteristic |
References
- Dubois, B.; Feldman, H.H.; Jacova, C.; DeKosky, S.T.; Barberger-Gateau, P.; Cummings, J.; Delacourte, A.; Galasko, D.; Gauthier, S.; Jicha, G.; et al. Research criteria for the diagnosis of alzheimer’s disease: Revising the NINCDS–ADRDA criteria. Lancet Neurol. 2007, 6, 734–746. [Google Scholar] [CrossRef]
- Perpetuini, D.; Cardone, D.; Bucco, R.; Zito, M.; Merla, A. Assessment of the autonomic response in alzehimer’s patients during the execution of memory tasks: A functional thermal imaging study. Curr. Alzheimer Res. 2018, 15, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Filippini, C.; Perpetuini, D.; Cardone, D.; Chiarelli, A.M.; Merla, A. Thermal Infrared Imaging and Artificial Intelligence Techniques Can Support Mild Alzheimer Disease Diagnosis; CEUR Workshop Proceedings: Aachen, Germany, 2020; Volume 2804, pp. 31–39. [Google Scholar]
- Reitz, C.; Brayne, C.; Mayeux, R. Epidemiology of alzheimer disease. Nat. Rev. Neurol. 2011, 7, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Finch, C.E.; Cohen, D.M. Aging, metabolism, and alzheimer disease: Review and hypotheses. Exp. Neurol. 1997, 143, 82–102. [Google Scholar] [CrossRef] [PubMed]
- Alsunusi, S.; Kumosani, T.A.; Glabe, C.G.; Huwait, E.A.; Moselhy, S.S. In Vitro study of the mechanism of intraneuronal β-amyloid aggregation in alzheimer’s disease. Arch. Physiol. Biochem. 2020, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Mamun, A.A.; Uddin, M.S.; Mathew, B.; Ashraf, G.M. Toxic tau: Structural origins of tau aggregation in alzheimer’s disease. Neural Regen Res 2020, 15, 1417–1420. [Google Scholar] [CrossRef]
- Mandelkow, E.-M.; Mandelkow, E. Tau in alzheimer’s disease. Trends Cell Biol. 1998, 8, 425–427. [Google Scholar] [CrossRef]
- Gómez-Isla, T.; Hollister, R.; West, H.; Mui, S.; Growdon, J.H.; Petersen, R.C.; Parisi, J.E.; Hyman, B.T. Neuronal loss correlates with but exceeds neurofibrillary tangles in alzheimer’s disease. Ann. Neurol. 1997, 41, 17–24. [Google Scholar] [CrossRef]
- Koper, M.J.; Van Schoor, E.; Ospitalieri, S.; Vandenberghe, R.; Vandenbulcke, M.; von Arnim, C.A.F.; Tousseyn, T.; Balusu, S.; De Strooper, B.; Thal, D.R. Necrosome complex detected in granulovacuolar degeneration is associated with neuronal loss in alzheimer’s disease. Acta Neuropathol. 2020, 139, 463–484. [Google Scholar] [CrossRef]
- Grammas, P. Neurovascular dysfunction, inflammation and endothelial activation: Implications for the pathogenesis of alzheimer’s disease. J. Neuroinflamm. 2011, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Nelson, A.R.; Sweeney, M.D.; Sagare, A.P.; Zlokovic, B.V. Neurovascular dysfunction and neurodegeneration in dementia and alzheimer’s disease. Biochim. Et Biophys. Acta (Bba) Mol. Basis Dis. 2016, 1862, 887–900. [Google Scholar] [CrossRef]
- Zlokovic, B.V. Neurovascular mechanisms of alzheimer’s neurodegeneration. Trends Neurosci. 2005, 28, 202–208. [Google Scholar] [CrossRef]
- Kisler, K.; Nelson, A.R.; Montagne, A.; Zlokovic, B.V. Cerebral blood flow regulation and neurovascular dysfunction in alzheimer disease. Nat. Rev. Neurosci. 2017, 18, 419–434. [Google Scholar] [CrossRef] [Green Version]
- Kotliar, K.; Hauser, C.; Ortner, M.; Muggenthaler, C.; Diehl-Schmid, J.; Angermann, S.; Hapfelmeier, A.; Schmaderer, C.; Grimmer, T. Altered neurovascular coupling as measured by optical imaging: A biomarker for alzheimer’s disease. Sci. Rep. 2017, 7, 12906. [Google Scholar] [CrossRef]
- Perpetuini, D.; Chiarelli, A.M.; Cardone, D.; Filippini, C.; Bucco, R.; Zito, M.; Merla, A. Complexity of frontal cortex FNIRS Can support alzheimer disease diagnosis in memory and visuo-spatial tests. Entropy 2019, 21, 26. [Google Scholar] [CrossRef] [Green Version]
- Chow, B.W.; Nuñez, V.; Kaplan, L.; Granger, A.J.; Bistrong, K.; Zucker, H.L.; Kumar, P.; Sabatini, B.L.; Gu, C. Caveolae in CNS arterioles mediate neurovascular coupling. Nature 2020, 579, 106–110. [Google Scholar] [CrossRef]
- Croce, P.; Zappasodi, F.; Merla, A.; Chiarelli, A.M. Exploiting neurovascular coupling: A bayesian sequential monte carlo approach applied to simulated EEG FNIRS data. J. Neural Eng. 2017, 14, 046029. [Google Scholar] [CrossRef]
- Croce, P.; Basti, A.; Marzetti, L.; Zappasodi, F.; Del Gratta, C. EEG? FMRI bayesian framework for neural activity estimation: A simulation study. J. Neural Eng. 2016, 13, 066017. [Google Scholar] [CrossRef]
- Haydon, P.G.; Carmignoto, G. Astrocyte control of synaptic transmission and neurovascular coupling. Physiol. Rev. 2006, 86, 1009–1031. [Google Scholar] [CrossRef] [Green Version]
- Uludağ, K.; Dubowitz, D.J.; Yoder, E.J.; Restom, K.; Liu, T.T.; Buxton, R.B. Coupling of cerebral blood flow and oxygen consumption during physiological activation and deactivation measured with FMRI. NeuroImage 2004, 23, 148–155. [Google Scholar] [CrossRef]
- Buxton, R.B. Introduction to Functional Magnetic Resonance Imaging: Principles and Techniques; Cambridge University Press: Cambridge, UK, 2009; ISBN 978-1-139-48130-4. [Google Scholar]
- Penny, W.D.; Friston, K.J.; Ashburner, J.T.; Kiebel, S.J.; Nichols, T.E. Statistical Parametric Mapping: The Analysis of Functional Brain Images; Academic Press: Cambridge, MA, USA, 2011; ISBN 978-0-08-046650-7. [Google Scholar]
- Parker, A.; Derrington, A.; Blakemore, C.; Logothetis, N.K. The neural basis of the blood–oxygen–level–dependent functional magnetic resonance imaging signal. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2002, 357, 1003–1037. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, M.; Quaresima, V. A brief review on the history of human functional near-infrared spectroscopy (FNIRS) development and fields of application. Neuroimage 2012, 63, 921–935. [Google Scholar] [CrossRef]
- Quaresima, V.; Ferrari, M. Functional Near-infrared spectroscopy (FNIRS) for assessing cerebral cortex function during human behavior in natural/social situations: A concise review. Organ. Res. Methods 2019, 22, 46–68. [Google Scholar] [CrossRef]
- Arenth, P.M.; Ricker, J.H.; Schultheis, M.T. Applications of functional near-infrared spectroscopy (FNIRS) to Neurorehabilitation of cognitive disabilities. Clin. Neuropsychol. 2007, 21, 38–57. [Google Scholar] [CrossRef]
- Costantini, M.; Vacri, A.D.; Chiarelli, A.M.; Ferri, F.; Romani, G.L.; Merla, A. Studying social cognition using near-infrared spectroscopy: The case of social simon effect. JBO JBOPFO 2013, 18, 025005. [Google Scholar] [CrossRef]
- Irani, F.; Platek, S.M.; Bunce, S.; Ruocco, A.C.; Chute, D. Functional near infrared spectroscopy (FNIRS): An emerging neuroimaging technology with important applications for the study of brain disorders. Clin. Neuropsychol. 2007, 21, 9–37. [Google Scholar] [CrossRef]
- Naseer, N.; Hong, K.-S. FNIRS-based Brain-computer interfaces: A review. Front. Hum. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perpetuini, D.; Bucco, R.; Zito, M.; Merla, A. Study of memory deficit in alzheimer’s disease by means of complexity analysis of FNIRS signal. Neurophotonics 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Pinti, P.; Aichelburg, C.; Lind, F.; Power, S.; Swingler, E.; Merla, A.; Hamilton, A.; Gilbert, S.; Burgess, P.; Tachtsidis, I. Using fiberless, wearable FNIRS to monitor brain activity in real-world cognitive tasks. J. Vis. Exp. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, H.; Shitara, Y.; Aoki, Y.; Inoue, T.; Tsuchida, S.; Takahashi, N.; Taga, G. Hemoglobin phase of oxygenation and deoxygenation in early brain development measured using FNIRS. Proc. Natl. Acad. Sci. USA 2017, 114, E1737–E1744. [Google Scholar] [CrossRef] [Green Version]
- Bennett, C.M.; Miller, M.B. FMRI reliability: Influences of Task and experimental design. Cogn. Affect. Behav. Neurosci. 2013, 13, 690–702. [Google Scholar] [CrossRef]
- Lindquist, M.A.; Meng Loh, J.; Atlas, L.Y.; Wager, T.D. Modeling the hemodynamic response function in FMRI: Efficiency, bias and mis-modeling. NeuroImage 2009, 45, S187–S198. [Google Scholar] [CrossRef] [Green Version]
- Friston, K.J.; Mechelli, A.; Turner, R.; Price, C.J. Nonlinear responses in FMRI: The balloon model, volterra kernels, and other hemodynamics. Neuroimage 2000, 12, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Jasdzewski, G.; Strangman, G.; Wagner, J.; Kwong, K.K.; Poldrack, R.A.; Boas, D.A. Differences in the hemodynamic response to event-related motor and visual paradigms as measured by near-infrared spectroscopy. NeuroImage 2003, 20, 479–488. [Google Scholar] [CrossRef]
- Steffener, J.; Tabert, M.; Reuben, A.; Stern, Y. Investigating hemodynamic response variability at the group level using basis functions. NeuroImage 2010, 49, 2113–2122. [Google Scholar] [CrossRef]
- Niedermeyer, E.; da Silva, F.H.L. Electroencephalography: Basic Principles, Clinical Applications, and Related Fields; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2005; ISBN 978-0-7817-5126-1. [Google Scholar]
- Hämäläinen, M.; Hari, R.; Ilmoniemi, R.J.; Knuutila, J.; Lounasmaa, O.V. Magnetoencephalography—Theory, instrumentation, and applications to noninvasive studies of the working human brain. Rev. Mod. Phys. 1993, 65, 413–497. [Google Scholar] [CrossRef] [Green Version]
- Ritter, P.; Villringer, A. Simultaneous EEG–FMRI. Neurosci. Biobehav. Rev. 2006, 30, 823–838. [Google Scholar] [CrossRef]
- Schulz, M.; Chau, W.; Graham, S.J.; McIntosh, A.R.; Ross, B.; Ishii, R.; Pantev, C. An integrative MEG–FMRI study of the primary somatosensory cortex using cross-modal correspondence analysis. NeuroImage 2004, 22, 120–133. [Google Scholar] [CrossRef]
- Stickland, R.; Allen, M.; Magazzini, L.; Singh, K.D.; Wise, R.G.; Tomassini, V. Neurovascular coupling during visual stimulation in multiple sclerosis: A MEG-FMRI STUDY. Neuroscience 2019, 403, 54–69. [Google Scholar] [CrossRef]
- Brueggen, K.; Fiala, C.; Berger, C.; Ochmann, S.; Babiloni, C.; Teipel, S.J. Early changes in alpha band power and dmn bold activity in alzheimer’s disease: A Simultaneous resting state EEG-FMRI study. Front. Aging Neurosci. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Chiarelli, A.M.; Zappasodi, F.; Pompeo, F.D.; Merla, A. Simultaneous functional near-infrared spectroscopy and electroencephalography for monitoring of human brain activity and oxygenation: A review. NphNeurow 2017, 4, 041411. [Google Scholar] [CrossRef]
- Li, R.; Potter, T.; Huang, W.; Zhang, Y. Enhancing Performance of a hybrid EEG-FNIRS system using channel selection and early temporal features. Front. Hum. Neurosci. 2017, 11, 462. [Google Scholar] [CrossRef] [Green Version]
- Perpetuini, D.; Chiarelli, A.M.; Filippini, C.; Cardone, D.; Croce, P.; Rotunno, L.; Anzoletti, N.; Zito, M.; Zappasodi, F.; Merla, A. Working memory decline in alzheimer’s disease is detected by complexity analysis of multimodal EEG-FNIRS. Entropy 2020, 22, 1380. [Google Scholar] [CrossRef]
- Davidson, R.J.; Jackson, D.C.; Larson, C.L. Human electroencephalography. In Handbook of Psychophysiology, 2nd ed.; Cambridge University Press: New York, NY, USA, 2000; pp. 27–52. ISBN 978-0-521-62634-7. [Google Scholar]
- Kiloh, L.G.; McComas, A.J.; Osselton, J.W. Clinical Electroencephalography; Butterworth-Heinemann: Oxford, UK, 2013; ISBN 978-1-4831-9215-4. [Google Scholar]
- Li, R.; Nguyen, T.; Potter, T.; Zhang, Y. Dynamic cortical connectivity alterations associated with alzheimer’s disease: An EEG and FNIRS integration study. Neuroimage Clin. 2019, 21, 101622. [Google Scholar] [CrossRef]
- Cicalese, P.A.; Li, R.; Ahmadi, M.B.; Wang, C.; Francis, J.T.; Selvaraj, S.; Schulz, P.E.; Zhang, Y. An EEG-FNIRS hybridization technique in the four-class classification of alzheimer’s disease. J. Neurosci. Methods 2020, 336, 108618. [Google Scholar] [CrossRef]
- Dutta, A.; Jacob, A.; Chowdhury, S.R.; Das, A.; Nitsche, M.A. EEG-NIRS based assessment of neurovascular coupling during anodal transcranial direct current stimulation—A stroke case series. J. Med. Syst. 2015, 39, 36. [Google Scholar] [CrossRef] [PubMed]
- Hendrikx, D.; Smits, A.; Lavanga, M.; De Wel, O.; Thewissen, L.; Jansen, K.; Caicedo, A.; Van Huffel, S.; Naulaers, G. Measurement of neurovascular coupling in neonates. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Othman, M.H.; Bhattacharya, M.; Møller, K.; Kjeldsen, S.; Grand, J.; Kjaergaard, J.; Dutta, A.; Kondziella, D. Resting-state NIRS–EEG in unresponsive patients with acute brain injury: A proof-of-concept study. Neurocrit Care 2020. [Google Scholar] [CrossRef] [PubMed]
- Csipo, T.; Mukli, P.; Lipecz, A.; Tarantini, S.; Bahadli, D.; Abdulhussein, O.; Owens, C.; Kiss, T.; Balasubramanian, P.; Nyúl-Tóth, Á.; et al. Assessment of Age-related decline of neurovascular coupling responses by functional near-infrared spectroscopy (FNIRS) in humans. GeroScience 2019, 41, 495–509. [Google Scholar] [CrossRef]
- Chiarelli, A.M.; Romani, G.L.; Merla, A. Fast optical signals in the sensorimotor cortex: General linear convolution model applied to multiple source–detector distance-based data. NeuroImage 2014, 85, 245–254. [Google Scholar] [CrossRef]
- Monti, M.M. Statistical analysis of FMRI time-series: A critical review of the GLM approach. Front. Hum. Neurosci. 2011, 5. [Google Scholar] [CrossRef] [Green Version]
- Perpetuini, D.; Cardone, D.; Filippini, C.; Chiarelli, A.M.; Merla, A. Modelling impulse response function of functional infrared imaging for general linear model analysis of autonomic activity. Sensors 2019, 19. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Zhao, C.; Wang, C.; Wang, J.; Zhang, Y. Enhancing FNIRS analysis using EEG rhythmic signatures: An EEG-informed FNIRS analysis study. IEEE Trans. Biomed. Eng. 2020, 67, 2789–2797. [Google Scholar] [CrossRef] [PubMed]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-Mental State”: A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Tucker, D.M. Spatial sampling of head electrical fields: The geodesic sensor net. Electroencephalogr. Clin. Neurophysiol. 1993, 87, 154–163. [Google Scholar] [CrossRef]
- Homan, R.W.; Herman, J.; Purdy, P. Cerebral location of international 10–20 system electrode placement. Electroencephalogr. Clin. Neurophysiol. 1987, 66, 376–382. [Google Scholar] [CrossRef]
- Brigadoi, S.; Cooper, R.J. How short is short? Optimum source-detector distance for short-separation channels in functional near-infrared spectroscopy. Neurophotonics 2015, 2, 025005. [Google Scholar] [CrossRef] [Green Version]
- Tachtsidis, I.; Scholkmann, F. False positives and false negatives in functional near-infrared spectroscopy: Issues, challenges, and the way forward. NPh 2016, 3, 031405. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, L.; Cooper, R.J.; Yücel, M.A.; Perdue, K.L.; Greve, D.N.; Boas, D.A. Short separation channel location impacts the performance of short channel regression in NIRS. NeuroImage 2012, 59, 2518–2528. [Google Scholar] [CrossRef] [Green Version]
- Dehghani, H.; Eames Matthew, E.; Yalavarthy Phaneendra, K.; Davis Scott, C.; Srinivasan, S.; Carpenter Colin, M.; Pogue Brian, W.; Paulsen Keith, D. Near infrared optical tomography using NIRFAST: Algorithm for numerical model and image reconstruction. Commun. Numer. Methods Eng. 2008, 25, 711–732. [Google Scholar] [CrossRef]
- Barbati, G.; Porcaro, C.; Zappasodi, F.; Rossini, P.M.; Tecchio, F. Optimization of an independent component analysis approach for artifact identification and removal in magnetoencephalographic signals. Clin. Neurophysiol. 2004, 115, 1220–1232. [Google Scholar] [CrossRef] [PubMed]
- Croce, P.; Zappasodi, F.; Marzetti, L.; Merla, A.; Pizzela, V.; Chiarelli, A.M. Deep convolutional neural networks for feature-less automatic classification of independent components in multi-channel electrophysiological brain recordings. IEEE Trans. Biomed. Eng. 2018, 1. [Google Scholar] [CrossRef] [PubMed]
- Chiarelli, A.M.; Maclin, E.L.; Fabiani, M.; Gratton, G. A Kurtosis-based wavelet algorithm for motion artifact correction of FNIRS data. NeuroImage 2015, 112, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Kocsis, L.; Herman, P.; Eke, A. The modified beer–lambert law revisited. Phys. Med. Biol. 2006, 51, N91–N98. [Google Scholar] [CrossRef]
- Zijlstra, W.G.; Buursma, A.; Meeuwsen-van der Roest, W.P. Absorption spectra of human fetal and adult oxyhemoglobin, de-oxyhemoglobin, carboxyhemoglobin, and methemoglobin. Clin. Chem. 1991, 37, 1633–1638. [Google Scholar] [CrossRef]
- Chiarelli, A.M.; Perpetuini, D.; Filippini, C.; Cardone, D.; Merla, A. Differential pathlength factor in continuous wave functional near-infrared spectroscopy: Reducing hemoglobin’s cross talk in high-density recordings. NPh 2019, 6, 035005. [Google Scholar] [CrossRef] [Green Version]
- Scholkmann, F.; Wolf, M. General equation for the differential pathlength factor of the frontal human head depending on wavelength and age. J. Biomed. Opt. 2013, 18, 105004. [Google Scholar] [CrossRef] [Green Version]
- Musso, F.; Brinkmeyer, J.; Mobascher, A.; Warbrick, T.; Winterer, G. Spontaneous brain activity and EEG microstates. A novel EEG/FMRI analysis approach to explore resting-state networks. Neuroimage 2010, 52, 1149–1161. [Google Scholar] [CrossRef]
- Yuan, H.; Zotev, V.; Phillips, R.; Drevets, W.C.; Bodurka, J. Spatiotemporal dynamics of the brain at rest—Exploring EEG microstates as electrophysiological signatures of BOLD resting state networks. NeuroImage 2012, 60, 2062–2072. [Google Scholar] [CrossRef]
- Hogg, R.; McKean, J.; Craig, A. Introduction to Mathematical Statistics, 6th ed.; Pearson Education, Inc.: London, UK, 2005. [Google Scholar]
- Johnson, R.A.; Wichern, D.W. Multivariate analysis. In Encyclopedia of Statistical Sciences; American Cancer Society: Atlanta, GA, USA, 2006; ISBN 978-0-471-66719-3. [Google Scholar]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning: Data Mining, Inference, and Prediction, 2nd ed.; Springer Series in Statistics; 2nd ed. Springer: New York, NY, USA, 2009; ISBN 978-0-387-84857-0. [Google Scholar]
- Liu, R.; Gillies, D.F. Overfitting in linear feature extraction for classification of high-dimensional image data. Pattern Recognit. 2016, 53, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Kohavi, R. A study of cross-validation and bootstrap for accuracy estimation and model selection. In Proceedings of the 14th International Joint Conference on Artificial Intelligence—Volume 2; Morgan Kaufmann Publishers Inc.: San Francisco, CA, USA, 1995; pp. 1137–1143. [Google Scholar]
- Esterman, M.; Tamber-Rosenau, B.J.; Chiu, Y.-C.; Yantis, S. Avoiding non-independence in FMRI data analysis: Leave one subject out. NeuroImage 2010, 50, 572–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, R.C. Mild cognitive impairment: Transition between aging and alzheimer’s disease. NEUROLOGIA 2000, 15, 93–101. [Google Scholar] [CrossRef]
- Serrano-Pozo, A.; Frosch, M.P.; Masliah, E.; Hyman, B.T. Neuropathological alterations in alzheimer disease. Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef]
- Logothetis, N.K.; Pauls, J.; Augath, M.; Trinath, T.; Oeltermann, A. Neurophysiological investigation of the basis of the FMRI signal. Nature 2001, 412, 150–157. [Google Scholar] [CrossRef]
- Laufs, H.; Kleinschmidt, A.; Beyerle, A.; Eger, E.; Salek-Haddadi, A.; Preibisch, C.; Krakow, K. EEG-correlated FMRI of human alpha activity. NeuroImage 2003, 19, 1463–1476. [Google Scholar] [CrossRef]
- Ekstrom, A.; Suthana, N.; Millett, D.; Fried, I.; Bookheimer, S. Correlation between BOLD FMRI and theta-band local field potentials in the human hippocampal area. J. Neurophysiol. 2009, 101, 2668–2678. [Google Scholar] [CrossRef] [Green Version]
- Salat, D.H.; Kaye, J.A.; Janowsky, J.S. Selective preservation and degeneration within the prefrontal cortex in aging and alzheimer disease. Arch. Neurol. 2001, 58, 1403–1408. [Google Scholar] [CrossRef]
- Brun, A.; Gustafson, L. Distribution of cerebral degeneration in alzheimer’s disease. Arch. Für Psychiatr. Und Nervenkrankh. 1976, 223, 15–33. [Google Scholar] [CrossRef]
- Mahady, L.; Nadeem, M.; Malek-Ahmadi, M.; Chen, K.; Perez, S.E.; Mufson, E.J. Frontal cortex epigenetic dysregulation during the progression of alzheimer’s disease. J. Alzheimer’s Dis. 2018, 62, 115–131. [Google Scholar] [CrossRef]
- Buzsáki, G.; Anastassiou, C.A.; Koch, C. The origin of extracellular fields and currents—EEG, ECoG, LFP and spikes. Nat. Rev. Neurosci. 2012, 13, 407–420. [Google Scholar] [CrossRef]
- Chiarelli, A.M.; Maclin, E.L.; Low, K.A.; Mathewson, K.E.; Fabiani, M.; Gratton, G. Combining energy and laplacian regularization to accurately retrieve the depth of brain activity of diffuse optical tomographic data. J. Biomed. Opt. 2016, 21, 36008. [Google Scholar] [CrossRef] [Green Version]
- Chiarelli, A.M.; Maclin, E.L.; Low, K.A.; Fabiani, M.; Gratton, G. Comparison of procedures for co-registering scalp-recording locations to anatomical magnetic resonance images. J. Biomed. Opt. 2015, 20, 016009. [Google Scholar] [CrossRef] [Green Version]
- Swainson, R.; Hodges, J.R.; Galton, C.J.; Semple, J.; Michael, A.; Dunn, B.D.; Iddon, J.L.; Robbins, T.W.; Sahakian, B.J. Early detection and differential diagnosis of alzheimer’s disease and depression with neuropsychological tasks. DEM 2001, 12, 265–280. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AD | HC | p-Value | |
|---|---|---|---|
| Gender (M–F) | 9–8 | 9–9 | / |
| Age (years) (mean ± SD) | 75.1 ± 7.1 | 71.4 ± 7.8 | 0.1354 |
| Education (years) (mean ± SD) | 9.6 ± 4.5 | 10.9 ± 4.3 | 0.3719 |
| Mini Mental State Examination (MMSE) score (mean ± SD) | 22.9 ± 3.2 | 27.5 ± 2.2 | 4.05 × 10−6 |
| Label | HC | AD | TOT | |
|---|---|---|---|---|
| Counts | HC | 17 | 1 | 18 |
| AD | 2 | 15 | 17 | |
| % | HC | 94.5% | 5.5% | 100% |
| AD | 11.8% | 88.2% | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiarelli, A.M.; Perpetuini, D.; Croce, P.; Filippini, C.; Cardone, D.; Rotunno, L.; Anzoletti, N.; Zito, M.; Zappasodi, F.; Merla, A. Evidence of Neurovascular Un-Coupling in Mild Alzheimer’s Disease through Multimodal EEG-fNIRS and Multivariate Analysis of Resting-State Data. Biomedicines 2021, 9, 337. https://doi.org/10.3390/biomedicines9040337
Chiarelli AM, Perpetuini D, Croce P, Filippini C, Cardone D, Rotunno L, Anzoletti N, Zito M, Zappasodi F, Merla A. Evidence of Neurovascular Un-Coupling in Mild Alzheimer’s Disease through Multimodal EEG-fNIRS and Multivariate Analysis of Resting-State Data. Biomedicines. 2021; 9(4):337. https://doi.org/10.3390/biomedicines9040337
Chicago/Turabian StyleChiarelli, Antonio M., David Perpetuini, Pierpaolo Croce, Chiara Filippini, Daniela Cardone, Ludovica Rotunno, Nelson Anzoletti, Michele Zito, Filippo Zappasodi, and Arcangelo Merla. 2021. "Evidence of Neurovascular Un-Coupling in Mild Alzheimer’s Disease through Multimodal EEG-fNIRS and Multivariate Analysis of Resting-State Data" Biomedicines 9, no. 4: 337. https://doi.org/10.3390/biomedicines9040337