
Inhibition of Prolyl Oligopeptidase Prevents Consequences of Reperfusion following Intestinal Ischemia

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Surgical Procedure for Intestinal IRI
2.3. Experimental Groups
- Sham group: mice were operated on with surgical steps; however, they were not subjected to IR and were treated with either a vehicle (saline).
- IRI + vehicle group: mice (n = 10) were subjected to intestinal ischemia by SAO (30 min), followed by reperfusion (1 h);
- IRI + KYP-2047 1 mg/kg group: mice (n = 10) were subjected to surgical procedures, described as above, and KYP-2047 (1 mg/kg, i.p.) was administered 5 min prior to reperfusion);
- IRI + KYP-2047 2.5 mg/kg group: mice (n = 10) were subjected to surgical procedures described as above, and KYP-2047 (2.5 mg/kg i.p.) was administered 5 min prior to reperfusion);
- IRI+ KYP-2047 5 mg/kg group: mice (n = 10) were subjected to surgical procedures described as above, and KYP-2047 (5 mg/kg i.p.) was administered 5 min prior to reperfusion).
2.4. Histological Examination
2.5. Myeloperoxidase Activity (MPO)
2.6. Western Blot Analysis for PREP, NF-κB Pathway, Angiogenesis and Apoptosis Markers
2.7. Immunofluorescence (IF) for Zonula Occludens (ZO)-1 and Filaggrin (FLG)
2.8. Immunohistochemical Localization of Vascular Endothelial Growth Factor (VEGF) and CD34
2.9. TUNEL Assay
3. Results
3.1. Inhibition of PREP by KYP-2047 Reduces Intestinal IR-Induced Histological Damage and Neutrophil Accumulation in the Intestine
3.2. KYP-2047 Administration Led to Limit Progressive Intestinal Inflammation and Fibrosis following IRI
3.3. Inhibition of PREP Attenuated the NF-κB Pathway Activation following Intestinal IRI
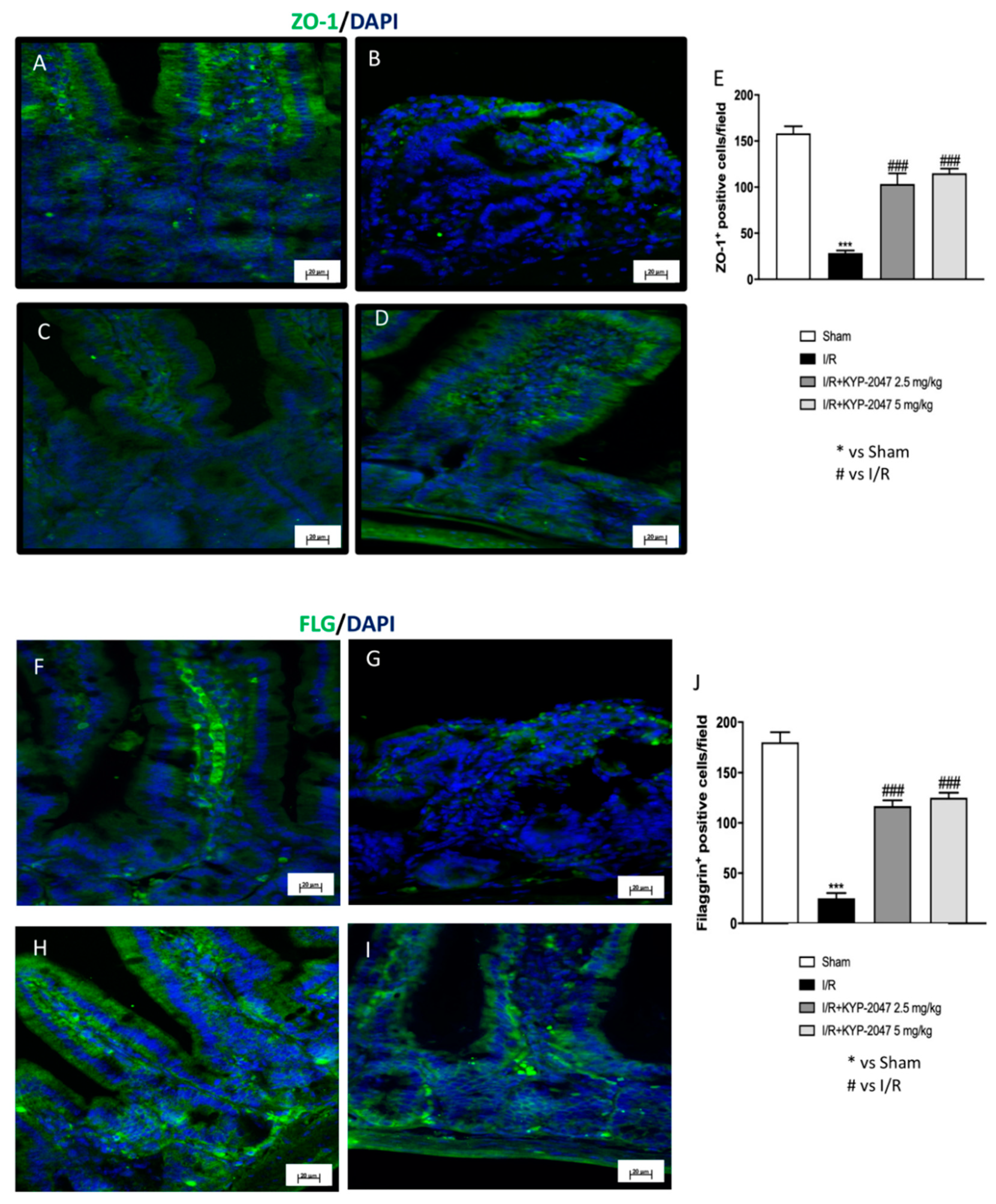
3.4. Treatment with KYP-2047 Stabilized the Expression of Tight Junction Zona Occludens-1 (ZO-1) and Restored Structural and Functional Role of Filaggrin (FLG) in the Intestine
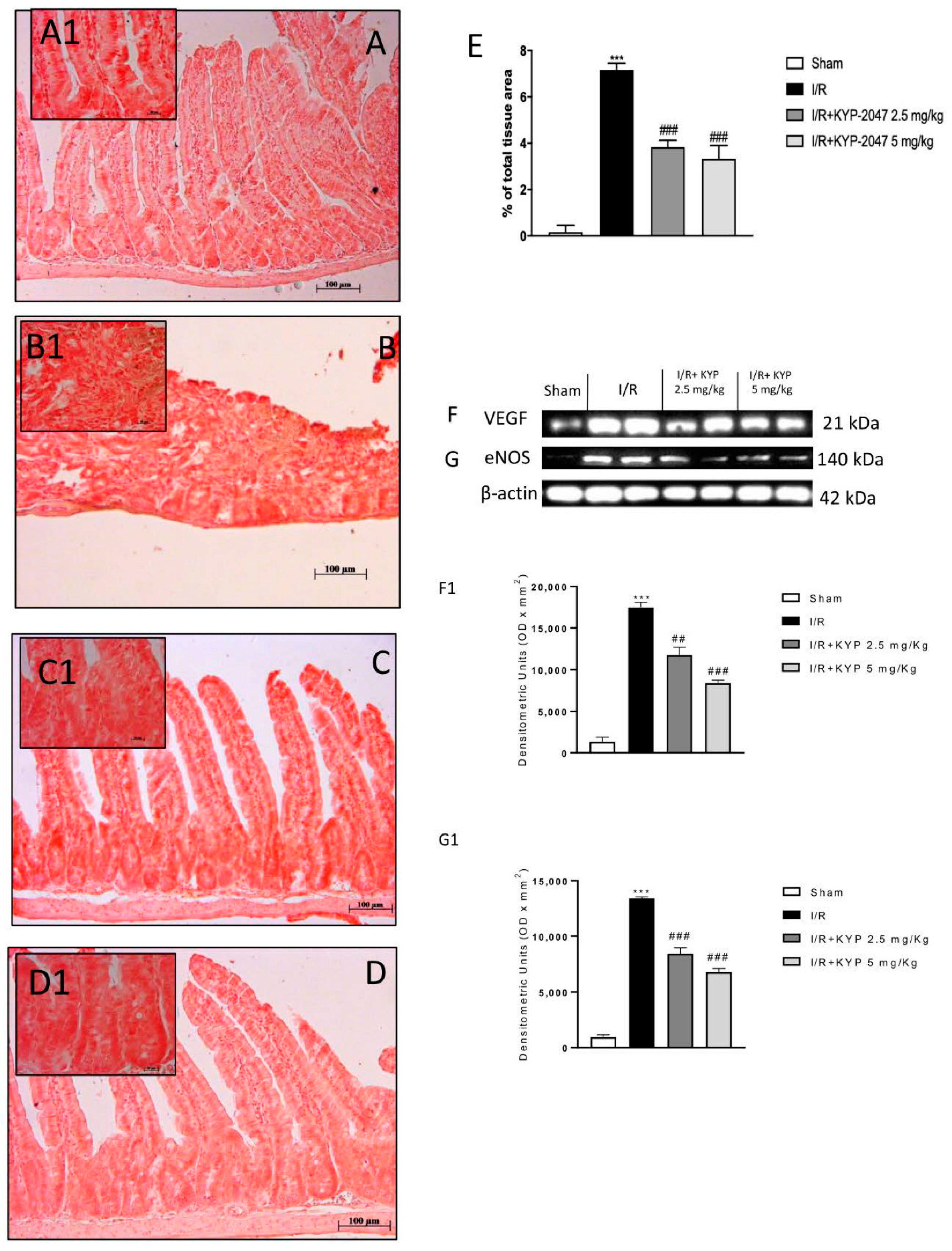
3.5. VEGF-Mediated Vascular Patterning and the Neovascular Process Are Regulated by KYP-2047 during IRI
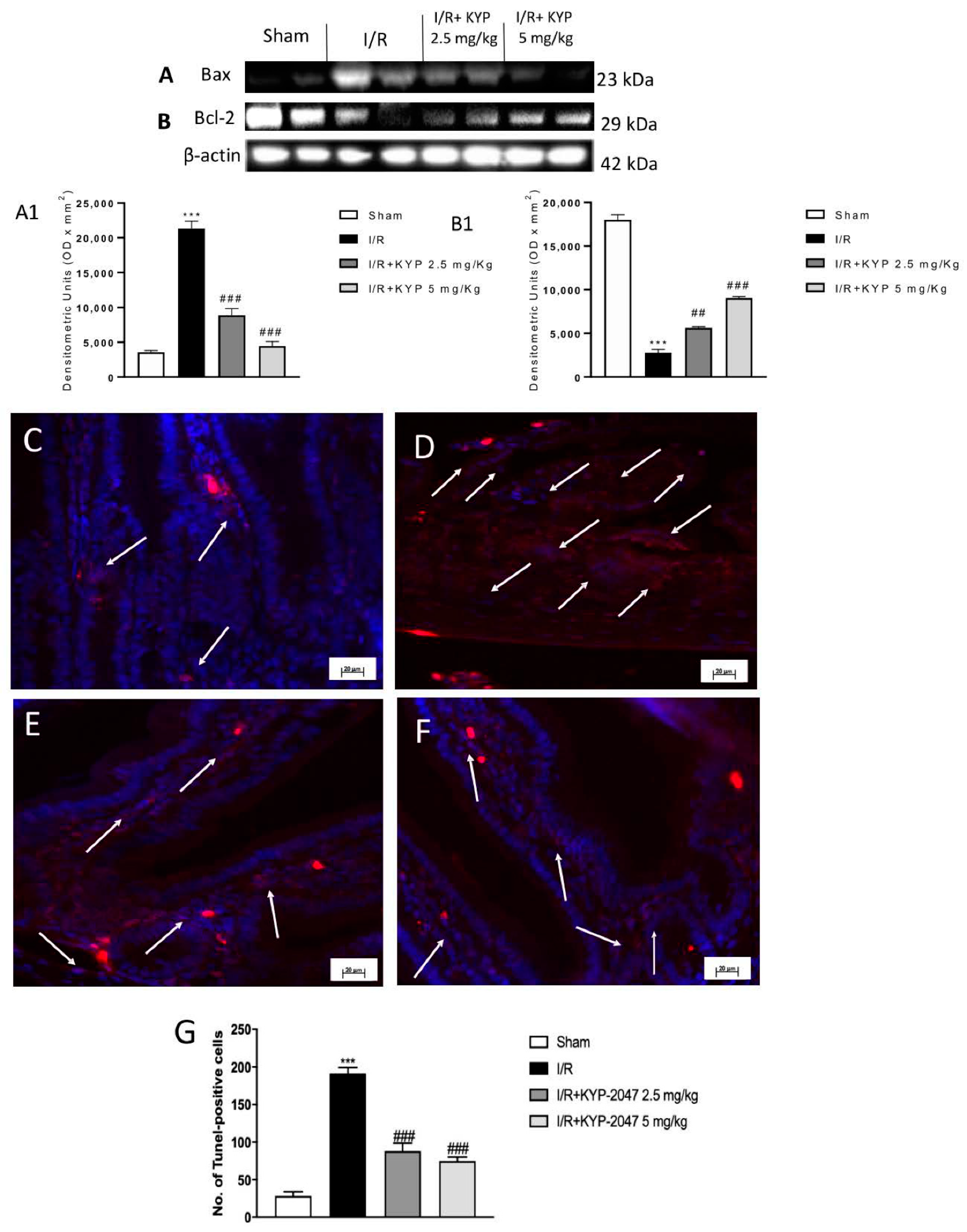
3.6. Apoptosis Effects of KYP-2047 on the Bcl-2/Bax Pathway and DNA Fragmentation following Intestinal IRI
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cerqueira, N.F.; Hussni, C.A.; Yoshida, W.B. Pathophysiology of mesenteric ischemia/reperfusion: A review. Acta Cir. Bras. 2005, 20, 336–343. [Google Scholar] [CrossRef]
- Zhang, Z.; Pan, C.; Wang, H.; Li, Y. Protective effects of osthole on intestinal ischemia-reperfusion injury in mice. Exp. Clin. Transplant. 2014, 12, 246–252. [Google Scholar] [CrossRef]
- Tsukamoto, T.; Chanthaphavong, R.S.; Pape, H.C. Current theories on the pathophysiology of multiple organ failure after trauma. Injury 2010, 41, 21–26. [Google Scholar] [CrossRef]
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight Junction in the Intestinal Epithelium: Its Association with Diseases and Regulation by Phytochemicals. J. Immunol. Res. 2018, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, F.G.; Sundback, C.; Kent, L.; Leachc, D.; Stanley, M.; Shleya, W.A.; Acanti, J.V.; Whanga, E. The effect of sustained delivery of vascular endothelial growth factor on angiogenesis in tissue-engineered intestine. Biomaterials 2019. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, G.; Feng, D.; Zu, G.; Li, Y.; Shi, X.; Zhao, Y.; Jing, H.; Ning, S.; Le, W.; et al. Targeting the miR-665-3p-ATG4B-autophagy axis relieves inflammation and apoptosis in intestinal ischemia/reperfusion. Cell Death Dis. 2018, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Svarcbahs, R.; Jäntti, M.; Kilpeläinen, T.; Julku, U.H.; Urvas, L.; Kivioja, S.; Norrbacka, S.; Myöhänen, T.T. Prolyl oligopeptidase inhibition activates autophagy via protein phosphatase 2A. Pharmacol. Res. 2020, 151, 104558. [Google Scholar] [CrossRef] [PubMed]
- Campolo, M.; Di Paola, R.; Impellizzeri, D.; Crupi, R.; Morittu, V.M.; Procopio, A.; Perri, E.; Britti, D.; Peli, A.; Esposito, E.; et al. Effects of a polyphenol present in olive oil, oleuropein aglycone, in a murine model of intestinal ischemia/reperfusion injury. J. Leukoc. Biol. 2012, 93, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Cordaro, M.; Campolo, M.; Gugliandolo, E.; Esposito, E.; Benedetto, F.; Cuzzocrea, S.; Navarra, M. Anti-inflammatory and Antioxidant Effects of Flavonoid-Rich Fraction of Bergamot Juice (BJe) in a Mouse Model of Intestinal Ische mia/Reperfusion Injury. Front. Pharmacol. 2016, 7, 203. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wu, J.; Liu, Q.; Li, X.; Li, S.; Chen, J.; Hong, Z.; Wu, X.; Zhao, Y.; Ren, J. mtDNA-STING pathway promotes necroptosis-dependent enterocyte injury in intestinal ischemia reperfusion. Cell Death Dis. 2020, 11, 1050. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.M.; Kwak, S.C.; Kim, J.Y.; Ahn, S.J.; Jun, H.Y.; Yoon, K.H.; Lee, M.S.; Oh, J. Evaluation of a novel technique for intraperitoneal injections in mice. Lab. Anim. 2015, 44, 440–444. [Google Scholar] [CrossRef]
- Filippone, A.; Lanza, M.; Campolo, M.; Casili, G.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. The Anti-Inflammatory and Antioxidant Effects of Sodium Propionate. Int. J. Mol. Sci. 2020, 21, 3026. [Google Scholar] [CrossRef] [PubMed]
- Mullane, K.M.; Kraemer, R.; Smith, B. Myeloperoxidase activity as a quantitative assessment of neutrophil infiltration into ischemic myocardium. J. Pharmacol. Methods 1985, 14, 157–167. [Google Scholar] [CrossRef]
- Filippone, A.; Lanza, M.; Campolo, M.; Casili, G.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Protective effect of sodium propionate in Aβ1-42 induced neurotoxicity and spinal cord trauma. Neuropharmacology 2020, 166, 107977. [Google Scholar] [CrossRef] [PubMed]
- Casili, G.; Lanza, M.; Scuderi, S.A.; Messina, S.; Paterniti, I.; Campolo, M.; Esposito, E. The Inhibition of Prolyl Oligopeptidase as New Target to Counteract Chronic Venous Insufficiency: Findings in a Mouse Model. Biomedicines 2020, 8, 604. [Google Scholar] [CrossRef] [PubMed]
- Bass, J.J.; Wilkinson, D.J.; Rankin, D.; Phillips, B.E.; Szewczyk, N.J.; Smith, K.; Atherton, P.J. An overview of technical considerations for Western blotting applications to physi ological research. Scand. J. Med. Sci. Sports 2017, 27, 4–25. [Google Scholar] [CrossRef] [PubMed]
- Campolo, M.; Siracusa, R.; Cordaro, M.; Filippone, A.; Gugliandolo, E.; Peritore, A.F.; Impellizzeri, D.; Crupi, R.; Paterniti, I.; Cuzzocrea, S. The association of adelmidrol with sodium hyaluronate displays beneficial properties against bladder changes following spinal cord injury in mice. PLoS ONE 2019, 14, e0208730. [Google Scholar] [CrossRef]
- Casili, G.; Campolo, M.; Paterniti, I.; Lanza, M.; Filippone, A.; Cuzzocrea, S.; Esposito, E. Dimethyl fumarate attenuates neuroinflammation and neurobehavioral deficits induced by experimental traumatic brain injury. J. Neurotrauma 2018, 35, 1437–1451. [Google Scholar] [CrossRef]
- Tas, U.; Ayan, M.; Sogut, E.; Kuloglu, T.; Uysal, M.; Tanriverdi, H.I.; Senel, U.; Ozyurt, B.; Sarsilmaz, M. Protective effects of thymoquinone and melatonin on intestinal ischemia–reperfusion injury. Saudi J. Gastroenterol. Off. J. Saudi Gastroenterol. Assoc. 2015, 21, 284–289. [Google Scholar] [CrossRef]
- Ban, K.; Kozar, R. Protective Role of p70S6K in Intestinal Ischemia/Reperfusion Injury in Mice. PLoS ONE 2012, 7, e41584. [Google Scholar] [CrossRef]
- Latella, G.; Rieder, F. Intestinal fibrosis: Ready to be Reversed. Curr. Opin. Gastroenterol. 2017, 33, 239–245. [Google Scholar] [CrossRef]
- Larrinaga, G.; Perez, I.; Blanco, L.; Sanz, B.; Errarte, P.; Beitia, M.; Etxezarraga, M.C.; Loizate, A.; Gil, J.; Irazusta, J.; et al. Prolyl Endopeptidase Activity Is Correlated with Colorectal Cancer Prognosis. Int. J. Med. Sci. 2014, 11, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Seetharaman, R.; Park, S.Y.; Yoon, M.K.; Kwak, J.O.; Son, B.G.; Park, Y.H.; Kag, J.M.; Lee, Y.J.; Choi, Y.C.; Bae, Y.S.; et al. Heme Oxygenase-1-mediated Anti-inflammatory Effect of a Novel Garlic Compound in RAW264.7 Murine Macrophages. Hort. Environ. Biotechnol. 2014, 55, 148–157. [Google Scholar] [CrossRef]
- Warners, M.J.; Vlieg-Boerstra, B.J.; Verheij, J.; Van Hamersveld, P.H.; Van Rhijn, B.D.; Van Ampting, M.T.; Harthoorn, L.F.; De Jonge, W.J.; Smout, A.J.; Bredenoord, A.J. Esophageal and Small Intestinal Mucosal Integrity in Eosinophilic Esophagitis and Response to an Elemental Diet. Am. J. Gastroenterol. 2017, 112, 1061–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrante, M.; Pierik, M.; Henckaerts, L.; Joossens, M.; Claes, K.; Van Schuerbeek, N.; Vlietinck, R.; Rutgeerts, P.; Van Assche, G.; Vermeire, S. The role of vascular endothelial growth factor (VEGF) in inflammatory bowel disease. Inflamm. Bowel Dis. 2006, 12, 870–878. [Google Scholar] [CrossRef]
- Papapetropoulos, A.; Garcia-Cardena, G.; Madri, J.A.; Sessa, W.C. Nitric oxide production contributes to the angiogenic properties of vascular endothelial growth factor in human endothelial cells. J. Clin. Investig. 1997, 100, 3131–3139. [Google Scholar] [CrossRef] [PubMed]
- Maltby, S.; Wohlfarth, C.; Gold, M.; Zbytnuik, L.; Hughes, M.R.; McNagny, K.M. CD34 Is Required for Infiltration of Eosinophils into the Colon and Pathology Associated with DSS-Induced Ulcerative Colitis. Am. J. Pathology 2010, 177, 1244–1254. [Google Scholar] [CrossRef]
- Stzepourginski, I.; Nigro, G.; Jacob, J.M.; Dulauroy, S.; Sansonetti, P.J.; Eberl, G.; Peduto, L. CD34+ mesenchymal cells are a major component of the intestinal stem cells niche at homeostasis and after injury. Proc. Natl. Acad. Sci. USA 2017, 114, E506–E513. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, A.; Madesh, M.; Balasubramanian, K. Apoptosis in the intestinal epithelium: Its relevance in normal and pathophysiological conditions. J. Gastroenterol. Hepatol. 2001, 15, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Block, T.A.; Acosta, S.; Bjorck, M. Endovascular and open surgery for acute occlusion of the superior mesenteric artery. J. Vasc. Surg. 2010, 52, 959–966. [Google Scholar] [CrossRef] [Green Version]
- Szabo’, A.; Vollmar, B.; Boros, M.; Menger, M.D. In vivo fluorescence microscopic imaging for dynamic quantitative assessment of intestinal mucosa permeability in mice. J. Surg. Res. 2008, 145, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Casili, G.; Lanza, M.; Campolo, M.; Messina, S.; Scuderi, S.; Ardizzone, A.; Filippone, A.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Therapeutic potential of flavonoids in the treatment of chronic venous insufficiency. Vasc. Pharmacol. 2021, 137, 106825. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Rimola, J.; Chen, M.H.; Rieder, F. Intestinal fibrosis: The Achilles heel of inflammatory bowel diseases? Editor. J. Dig. Dis. 2020, 21, 306–307. [Google Scholar] [CrossRef] [PubMed]
- Speca, S.; Giusti, I.; Rieder, F.; Latella, G. Cellular and molecular mechanisms of intestinal fibrosis. World J. Gastroenterol. 2012, 28, 3635–3661. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.M.; Moeser, A.J.; Blikslager, A.T. Animal models of ischemia-reperfusion-induced intestinal injury: Progress and promise for translational research. Physiol. Med. 2015, 308, G63–G75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.Y.; Hsiao, J.K.; Lu, Y.Z.; Lee, T.C.; Linda, C.H. Anti-apoptotic PI3K/Akt signaling by sodium/glucose transporter 1 reduces epithelial barrier damage and bacterial translocation in intestinal ischemia. Lab. Investig. 2011, 91, 294–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Qu, L.; Li, Y.; Gu, L.; Shi, M.; Zhang, J.; Zhu, W.; Li, J. Bone Marrow Mesenchymal Stem Cells Reduce Intestinal Ischemia/Reperfusion Injuries in Rats. J. Surg. Res. 2011, 168, 127–134. [Google Scholar] [CrossRef]
- Sandor, Z.; Deng, X.M.; Khomenko, T.; Tarnawski, A.S.; Szabo, S. Altered angiogenic balance in ulcerative colitis: A key to impaired healing? Biochem. Biophys. Res. Commun. 2006, 350, 147–150. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippone, A.; Casili, G.; Ardizzone, A.; Lanza, M.; Mannino, D.; Paterniti, I.; Esposito, E.; Campolo, M. Inhibition of Prolyl Oligopeptidase Prevents Consequences of Reperfusion following Intestinal Ischemia. Biomedicines 2021, 9, 1354. https://doi.org/10.3390/biomedicines9101354
Filippone A, Casili G, Ardizzone A, Lanza M, Mannino D, Paterniti I, Esposito E, Campolo M. Inhibition of Prolyl Oligopeptidase Prevents Consequences of Reperfusion following Intestinal Ischemia. Biomedicines. 2021; 9(10):1354. https://doi.org/10.3390/biomedicines9101354
Chicago/Turabian StyleFilippone, Alessia, Giovanna Casili, Alessio Ardizzone, Marika Lanza, Deborah Mannino, Irene Paterniti, Emanuela Esposito, and Michela Campolo. 2021. "Inhibition of Prolyl Oligopeptidase Prevents Consequences of Reperfusion following Intestinal Ischemia" Biomedicines 9, no. 10: 1354. https://doi.org/10.3390/biomedicines9101354