
Chronic Restraint Stress-Induced Muscle Atrophy Leads to Fatigue in Mice by Inhibiting the AMPK Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and CRS Treatments
2.2. Behavioral Tests
2.3. Biological Sample Collection
2.4. Determination of Biochemical Parameters
2.5. Transcriptomic Analysis
2.6. Western Blotting Analysis
2.7. Histological and Immunohistochemistry Analyses of Muscle
2.8. Transmission Electron Microscopy
2.9. Statistical Analysis
3. Results
3.1. CRS Induced Fatigue and Its Specific Dynamic Process in Mice
3.2. CRS Induced Muscle Atrophy and Disorder in Gastrocnemius Structure and Function
3.3. CRS Induced Changes in the Gene Expression Profile in the Gastrocnemius of Mice
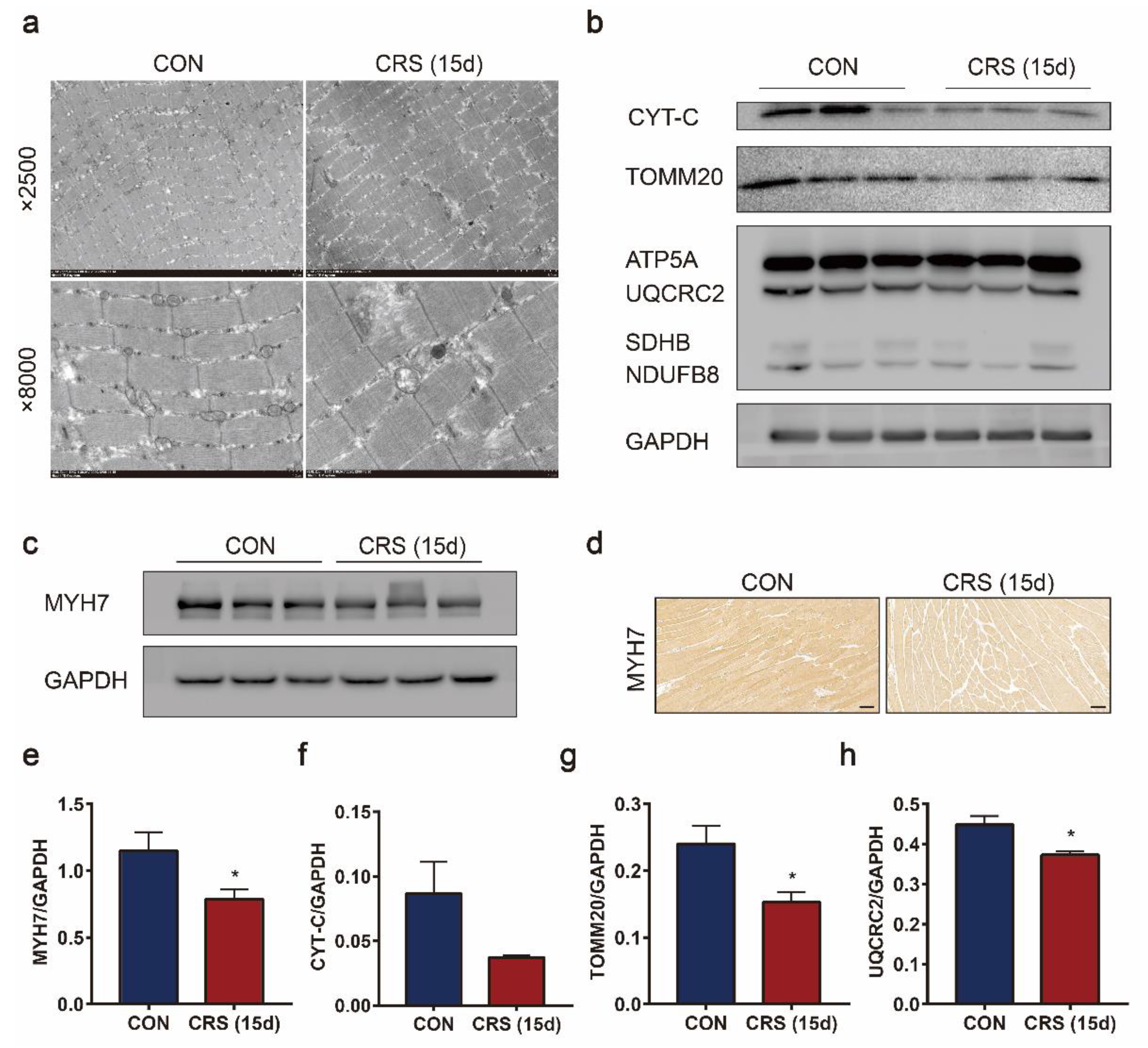
3.4. CRS Induced Mitochondrial Loss and Dysfunction in Muscle Cells
3.5. CRS Induced Muscle Dysfunction and Mitochondrial Biogenesis Decrease via AMPK/PGC-1α
3.6. CRS Blocked Mitophagy in Muscle Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dittner, A.J.; Wessely, S.C.; Brown, R.G. The assessment of fatigue: A practical guide for clinicians and researchers. J. Psychosom Res. 2004, 56, 157–170. [Google Scholar] [CrossRef]
- Azizoddin, D.R.; Jolly, M.; Arora, S.; Yelin, E.; Katz, P. Longitudinal Study of Fatigue, Stress, and Depression: Role of Reduction in Stress Toward Improvement in Fatigue. Arthritis Rheum. 2020, 72, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Ament, W.; Verkerke, G.J. Exercise and fatigue. Sports Med. 2009, 39, 389–422. [Google Scholar] [CrossRef] [PubMed]
- Bower, J.E. Cancer-related fatigue—Mechanisms, risk factors, and treatments. Nat. Rev. Clin. Oncol. 2014, 11, 597–609. [Google Scholar] [CrossRef]
- Kluger, B.M.; Friedman, J.H. Fatigue in Parkinson’s disease. Non-Mot. Symptoms Parkinson’s Dis. 2009, 133, 135–146. [Google Scholar] [CrossRef]
- Cropley, M.; Rydstedt, L.W.; Andersen, D. Recovery from work: Testing the effects of chronic internal and external workload on health and well-being. J. Epidemiol. Community Health 2020, 74, 919–924. [Google Scholar] [CrossRef]
- Maghout-Juratli, S.; Janisse, J.; Schwartz, K.; Arnetz, B.B. The Causal Role of Fatigue in the Stress-Perceived Health Relationship: A MetroNet Study. J. Am. Board Fam. Med. 2010, 23, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Krogstad, H.; Loge, J.H.; Grotmol, K.S.; Kaasa, S.; Kiserud, C.E.; Salvesen, Ø.; Hjermstad, M.J. Symptoms in the general Norwegian adult population-prevalence and associated factors. BMC Public Health 2020, 20, 988. [Google Scholar] [CrossRef]
- Roberts, E.; Wessely, S.; Chalder, T.; Chang, C.-K.; Hotopf, M. Mortality of people with chronic fatigue syndrome: A retrospective cohort study in England and Wales from the South London and Maudsley NHS Foundation Trust Biomedical Research Centre (SLaM BRC) Clinical Record Interactive Search (CRIS) Register. Lancet 2016, 387, 1638–1643. [Google Scholar] [CrossRef] [Green Version]
- Rashid, A.A.; Devaraj, N.K.; Yusof, H.M.; Mustapha, F.; Wong, S.V.; Ismail, A.F.; Ismail, K.I.; Qureshi, A.M.; Bin Nordin, R. Prevalence and predictors of road crash involvement among medical doctors in Malaysia: A cross-sectional study protocol. BMJ Open 2020, 10, e037653. [Google Scholar] [CrossRef]
- Li, M.; Shu, Q.; Huang, H.; Bo, W.; Wang, L.; Wu, H. Associations of occupational stress, workplace violence, and organizational support on chronic fatigue syndrome among nurses. J. Adv. Nurs. 2020, 76, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Parikh, S.; Galioto, R.; Lapin, B.; Haas, R.; Hirano, M.; Koenig, M.K.; Saneto, R.P.; Zolkipli-Cunningham, Z.; Goldstein, A.; Karaa, A. Fatigue in primary genetic mitochondrial disease: No rest for the weary. Neuromuscul. Disord. 2019, 29, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Gorman, G.S.; Elson, J.L.; Newman, J.; Payne, B.; McFarland, R.; Newton, J.L.; Turnbull, D. Perceived fatigue is highly prevalent and debilitating in patients with mitochondrial disease. Neuromuscul. Disord. 2015, 25, 563–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, D.C. Mitochondrial Dynamics and Its Involvement in Disease. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 235–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Marcos, P.J.; Auwerx, J. Regulation of PGC-1α, a nodal regulator of mitochondrial biogenesis. Am. J. Clin. Nutr. 2011, 93, 884S–890S. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yan, M.; Pan, R.; Wang, Z.; Tao, X.; Li, C.; Xia, T.; Liu, X.; Chang, Q. Radix Polygalae extract exerts antidepressant effects in behavioral despair mice and chronic restraint stress-induced rats probably by promoting autophagy and inhibiting neuroinflammation. J. Ethnopharmacol. 2021, 265, 113317. [Google Scholar] [CrossRef]
- Sahoo, S.; Kharkar, P.S.; Sahu, N.U. Anxiolytic activity of Psidium guajava in mice subjected to chronic restraint stress and effect on neurotransmitters in brain. Phytotherapy Res. 2021, 35, 1399–1415. [Google Scholar] [CrossRef]
- Oh, D.R.; Kim, Y.; Im, S.; Oh, K.N.; Shin, J.; Jeong, C.; Kim, Y.; Choi, E.J.; Choi, C. Vaccinium bracteatum Improves Spatial Learning and Memory by Regulating N-methyl-D-aspartate Receptors and Tau Phosphorylation in Chronic Restraint Stress-Induced Memory Impaired Mice. Am. J. Chin. Med. 2021, 49, 69–94. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Jang, S.; Son, E.; Lee, S.W.; Sung, Y.-Y.; Kim, H.K. Polygonum aviculare L. extract reduces fatigue by inhibiting neuroinflammation in restraint-stressed mice. Phytomedicine 2018, 42, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Jang, S.; Lee, S.W.; Sung, Y.-Y.; Kim, H.K. Akebia quinata Decaisne aqueous extract acts as a novel anti-fatigue agent in mice exposed to chronic restraint stress. J. Ethnopharmacol. 2018, 222, 270–279. [Google Scholar] [CrossRef]
- Picard, M.; McEwen, B.S. Psychological Stress and Mitochondria: A Systematic Review. Psychosom. Med. 2018, 80, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stults-Kolehmainen, M.A.; Sinha, R. The Effects of Stress on Physical Activity and Exercise. Sports Med. 2014, 44, 81–121. [Google Scholar] [CrossRef] [PubMed]
- Hogan, P.S.; Chen, S.X.; Teh, W.W.; Chib, V.S. Neural mechanisms underlying the effects of physical fatigue on effort-based choice. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Theilen, N.T.; Kunkel, G.H.; Tyagi, S.C. The Role of Exercise and TFAM in Preventing Skeletal Muscle Atrophy. J. Cell. Physiol. 2017, 232, 2348–2358. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Wei, Y.J.; Dong, X.X.; Liu, J.Y.; Chen, Q.Y.; Zhang, G.X. The effect of candesartan on chronic stress induced imbalance of glucose homeostasis. Biomed. Pharmacother. 2020, 128, 110300. [Google Scholar] [CrossRef]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef]
- Nascimbeni, A.C.; Fanin, M.; Masiero, E.; Angelini, C.; Sandri, M. Impaired autophagy contributes to muscle atrophy in glycogen storage disease type II patients. Autophagy 2012, 8, 1697–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Meo, S.; Napolitano, G.; Venditti, P. Mediators of Physical Activity Protection against ROS-Linked Skeletal Muscle Damage. Int. J. Mol. Sci. 2019, 20, 3024. [Google Scholar] [CrossRef] [Green Version]
- Fan, K.-Q.; Li, Y.-Y.; Wang, H.-L.; Mao, X.-T.; Guo, J.-X.; Wang, F.; Huang, L.-J.; Ma, X.-Y.; Gao, Z.-J.; Chen, W.; et al. Stress-Induced Metabolic Disorder in Peripheral CD4+ T Cells Leads to Anxiety-like Behavior. Cell 2019, 179, 864–879. [Google Scholar] [CrossRef]
- Hardie, D.G.; Iwadate, Y.; Yumura, S. The AMP-activated protein kinase pathway—New players upstream and downstream. J. Cell Sci. 2004, 117, 5479–5487. [Google Scholar] [CrossRef] [Green Version]
- Houten, S.; Auwerx, J. PGC-1α: Turbocharging mitochondria. Cell 2004, 119, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Reyes, N.L.; Banks, G.B.; Tsang, M.; Margineantu, D.; Gu, H.; Djukovic, D.; Chan, J.; Torres, M.; Liggitt, H.D.; Hirenallur-S, D.K.; et al. Fnip1 regulates skeletal muscle fiber type specification, fatigue resistance, and susceptibility to muscular dystrophy. Proc. Natl. Acad. Sci. USA 2015, 112, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kou, X.; Li, J.; Liu, X.; Yang, X.; Fan, J.; Chen, N. Ampelopsin attenuates the atrophy of skeletal muscle from d -gal-induced aging rats through activating AMPK/SIRT1/PGC-1α signaling cascade. Biomed. Pharmacother. 2017, 90, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Liao, Q.; Liu, J.; Pan, R.; Lee, S.M.; Lin, L. Myricanol rescues dexamethasone-induced muscle dysfunction via a sirtuin 1-dependent mechanism. J. Cachex- Sarcopenia Muscle 2019, 10, 429–444. [Google Scholar] [CrossRef]
- Lin, J.; Wu, P.-H.; Tarr, P.T.; Lindenberg, K.S.; St-Pierre, J.; Zhang, C.-Y.; Mootha, V.K.; Jager, S.; Vianna, C.R.; Reznick, R.M.; et al. Defects in Adaptive Energy Metabolism with CNS-Linked Hyperactivity in PGC-1α Null Mice. Cell 2004, 119, 121–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Wu, H.; Tarr, P.T.; Zhang, C.-Y.; Wu, Z.; Boss, O.; Michael, L.F.; Puigserver, P.; Isotani, E.; Olson, E.N.; et al. Transcriptional co-activator PGC-1α drives the formation of slow-twitch muscle fibres. Nat. Cell Biol. 2002, 418, 797–801. [Google Scholar] [CrossRef]
- Greenhaff, P.; Nevill, M.; Soderlund, K.; Bodin, K.; Boobis, L.H.; Williams, C.; Hultman, E. The metabolic responses of human type I and II muscle fibres during maximal treadmill sprinting. J. Physiol. 1994, 478, 149–155. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C. Fiber Types in Mammalian Skeletal Muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [Green Version]
- Masiero, E.; Agatea, L.; Mammucari, C.; Blaauw, B.; Loro, E.; Komatsu, M.; Metzger, D.; Reggiani, C.; Schiaffino, S.; Sandri, M. Autophagy Is Required to Maintain Muscle Mass. Cell Metab. 2009, 10, 507–515. [Google Scholar] [CrossRef]
- Quinn, P.M.J.; Moreira, P.I.; Ambrósio, A.F.; Alves, C.H. PINK1/PARKIN signalling in neurodegeneration and neuroinflammation. Acta Neuropathol. Commun. 2020, 8, 1–20. [Google Scholar] [CrossRef]
- Yoo, S.-M.; Jung, Y.-K. A Molecular Approach to Mitophagy and Mitochondrial Dynamics. Mol. Cells 2018, 41, 18–26. [Google Scholar]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in Mammalian Autophagy Research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, D.F.; Shackelford, D.B.; Mihaylova, M.M.; Gelino, S.; Kohnz, R.A.; Mair, W.; Vasquez, D.S.; Joshi, A.; Gwinn, D.M.; Taylor, R.; et al. Phosphorylation of ULK1 (hATG1) by AMP-Activated Protein Kinase Connects Energy Sensing to Mitophagy. Science 2011, 331, 456–461. [Google Scholar] [CrossRef] [Green Version]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.-L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Egan, D.F.; Kim, J.; Shaw, R.J.; Guan, K.-L. The autophagy initiating kinase ULK1 is regulated via opposing phosphorylation by AMPK and mTOR. Autophagy 2011, 7, 643–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quy, P.N.; Kuma, A.; Pierre, P.; Mizushima, N. Proteasome-dependent activation of mammalian target of rapamycin complex 1 (mTORC1) is essential for autophagy suppression and muscle remodeling following denervation. J. Biol. Chem. 2013, 288, 1125–1134. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Ren, J.; Chen, X.; Liu, J.; Xu, X.; Li, X.; Zhao, D.; Sun, L. 20(S)-ginsenoside Rg3 promotes myoblast differentiation and protects against myotube atrophy via regulation of the Akt/mTOR/FoxO3 pathway. Biochem. Pharmacol. 2020, 180, 114145. [Google Scholar] [CrossRef]
- Ham, D.J.; Börsch, A.; Lin, S.; Thürkauf, M.; Weihrauch, M.; Reinhard, J.R.; Delezie, J.; Battilana, F.; Wang, X.; Kaiser, M.S.; et al. The neuromuscular junction is a focal point of mTORC1 signaling in sarcopenia. Nat. Commun. 2020, 11, 1–21. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Xia, T.; Jin, S.; Liu, X.; Pan, R.; Yan, M.; Chang, Q. Chronic Restraint Stress-Induced Muscle Atrophy Leads to Fatigue in Mice by Inhibiting the AMPK Signaling Pathway. Biomedicines 2021, 9, 1321. https://doi.org/10.3390/biomedicines9101321
Wang Z, Xia T, Jin S, Liu X, Pan R, Yan M, Chang Q. Chronic Restraint Stress-Induced Muscle Atrophy Leads to Fatigue in Mice by Inhibiting the AMPK Signaling Pathway. Biomedicines. 2021; 9(10):1321. https://doi.org/10.3390/biomedicines9101321
Chicago/Turabian StyleWang, Zhi, Tianji Xia, Suwei Jin, Xinmin Liu, Ruile Pan, Mingzhu Yan, and Qi Chang. 2021. "Chronic Restraint Stress-Induced Muscle Atrophy Leads to Fatigue in Mice by Inhibiting the AMPK Signaling Pathway" Biomedicines 9, no. 10: 1321. https://doi.org/10.3390/biomedicines9101321