Low-Grade Inflammation Is Not Present in Former Obese Males but Adipose Tissue Macrophage Infiltration Persists

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Experimental Protocol
2.3. Analytical Procedures
2.4. Statistics
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bastard, J.P.; Maach, M.; Lagathu, C.; Kim, M.J.; Caron, M.; Vidal, H.; Capeau, J.; Feve, B. Recent advances in the relationship between obesity, inflammation, and insulin resistance. Eur. Cytokine Netw. 2006, 17, 4–12. [Google Scholar] [PubMed]
- Hill, A.A.; Reid, B.W.; Hasty, A.H. A decade of progress in adipose tissue macrophage biology. Immunol. Rev. 2014, 262, 134–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, E.W.; Carey, A.L.; Sacchetti, M.; Steinberg, G.R.; Macaulay, S.L.; Febbraio, M.A.; Pedersen, B.K. Acute IL-6 treatment increases fatty acid turnover in elderly humans in vivo and in tissue culture in vitro. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E155–E162. [Google Scholar] [CrossRef]
- Nicklas, B.J.; You, T.; Pahor, M. Behavioural treatments for chronic systemic inflammation: Effects of dietary weight loss and exercise training. CMAJ 2005, 172, 1199–1209. [Google Scholar] [CrossRef] [Green Version]
- Bruun, J.M.; Lihn, A.S.; Pedersen, S.B.; Richelsen, B. Monocyte chemoattractant protein-1 release is higher in visceral than subcutaneous human adipose tissue (AT): Implication of macrophages resident in the AT. J. Clin. Endocrinol. Metab. 2005, 90, 2282–2289. [Google Scholar] [CrossRef]
- Fain, J.N. Release of interleukins and other inflammatory cytokines by human adipose tissue is enhanced in obesity and primarily due to the nonfat cells. Vitam. Horm. 2006, 74, 443–477. [Google Scholar] [CrossRef]
- Miller, G.D.; Isom, S.; Morgan, T.M.; Vitolins, M.Z.; Blackwell, C.; Brosnihan, K.B.; Diz, D.I.; Katula, J.; Goff, D. Effects of a community-based weight loss intervention on adipose tissue circulating factors. Diabetes Metab. Syndr. 2014, 8, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Castoldi, A.; Naffah de, S.C.; Camara, N.O.; Moraes-Vieira, P.M. The Macrophage Switch in Obesity Development. Front. Immunol. 2015, 6, 637. [Google Scholar] [CrossRef] [Green Version]
- Roszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Goh, J.; Goh, K.P.; Abbasi, A. Exercise and Adipose Tissue Macrophages: New Frontiers in Obesity Research? Front. Endocrinol. (Lausanne) 2016, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Cancello, R.; Henegar, C.; Viguerie, N.; Taleb, S.; Poitou, C.; Rouault, C.; Coupaye, M.; Pelloux, V.; Hugol, D.; Bouillot, J.L.; et al. Reduction of macrophage infiltration and chemoattractant gene expression changes in white adipose tissue of morbidly obese subjects after surgery-induced weight loss. Diabetes 2005, 54, 2277–2286. [Google Scholar] [CrossRef] [Green Version]
- Auerbach, P.; Nordby, P.; Bendtsen, L.Q.; Mehlsen, J.L.; Basnet, S.K.; Vestergaard, H.; Ploug, T.; Stallknecht, B. Differential effects of endurance training and weight loss on plasma adiponectin multimers and adipose tissue macrophages in younger, moderately overweight men. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R490–R498. [Google Scholar] [CrossRef]
- Kovacikova, M.; Sengenes, C.; Kovacova, Z.; Siklova-Vitkova, M.; Klimcakova, E.; Polak, J.; Rossmeislova, L.; Bajzova, M.; Hejnova, J.; Hnevkovska, Z.; et al. Dietary intervention-induced weight loss decreases macrophage content in adipose tissue of obese women. Int. J. Obes. (Lond.) 2011, 35, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Aron-Wisnewsky, J.; Tordjman, J.; Poitou, C.; Darakhshan, F.; Hugol, D.; Basdevant, A.; Aissat, A.; Guerre-Millo, M.; Clement, K. Human adipose tissue macrophages: m1 and m2 cell surface markers in subcutaneous and omental depots and after weight loss. J. Clin. Endocrinol. Metab. 2009, 94, 4619–4623. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, M.D.; Lund, M.T.; Hansen, M.; Poulsen, S.S.; Ploug, T.; Dela, F.; Helge, J.W.; Prats, C. Macrophage Area Content and Phenotype in Hepatic and Adipose Tissue in Patients with Obesity Undergoing Roux-en-Y Gastric Bypass. Obesity. (Silver Spring) 2017, 25, 1921–1931. [Google Scholar] [CrossRef] [Green Version]
- Bruun, J.M.; Helge, J.W.; Richelsen, B.; Stallknecht, B. Diet and exercise reduce low-grade inflammation and macrophage infiltration in adipose tissue but not in skeletal muscle in severely obese subjects. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E961–E967. [Google Scholar] [CrossRef]
- You, T.; Ryan, A.S.; Nicklas, B.J. The metabolic syndrome in obese postmenopausal women: Relationship to body composition, visceral fat, and inflammation. J. Clin. Endocrinol. Metab. 2004, 89, 5517–5522. [Google Scholar] [CrossRef] [Green Version]
- Kovacova, Z.; Tencerova, M.; Roussel, B.; Wedellova, Z.; Rossmeislova, L.; Langin, D.; Polak, J.; Stich, V. The impact of obesity on secretion of adiponectin multimeric isoforms differs in visceral and subcutaneous adipose tissue. Int. J. Obes. (Lond.) 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.; Goedecke, J.H.; Soderstrom, I.; Buren, J.; Alvehus, M.; Blomquist, C.; Jonsson, F.; Hayes, P.M.; Adams, K.; Dave, J.A.; et al. Depot- and ethnic-specific differences in the relationship between adipose tissue inflammation and insulin sensitivity. Clin. Endocrinol. (Oxf.) 2011, 74, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Ara, I.; Larsen, S.; Stallknecht, B.; Guerra, B.; Morales-Alamo, D.; Andersen, J.L.; Ponce-Gonzalez, J.G.; Guadalupe-Grau, A.; Galbo, H.; Calbet, J.A.; et al. Normal mitochondrial function and increased fat oxidation capacity in leg and arm muscles in obese humans. Int. J. Obes. (Lond.) 2011, 35, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergström, J. Muscle electrolytes in man: Determined by neutron activation analysis on needle biopsy specimens. A study on normal subjects, kidney patients and patients with chronic diarrhea. Scand. J. Clin. Lab. Invest. Supplementum. 1962, 68, 11–13. [Google Scholar]
- Katz, A.; Nambi, S.S.; Mather, K.; Baron, A.D.; Follmann, D.A.; Sullivan, G.; Quon, M.J. Quantitative insulin sensitivity check index: A simple, accurate method for assessing insulin sensitivity in humans. J. Clin. Endocrinol. Metab. 2000, 85, 2402–2410. [Google Scholar] [CrossRef]
- Arvidsson, E.; Viguerie, N.; Andersson, I.; Verdich, C.; Langin, D.; Arner, P. Effects of different hypocaloric diets on protein secretion from adipose tissue of obese women. Diabetes 2004, 53, 1966–1971. [Google Scholar] [CrossRef] [Green Version]
- Tam, C.S.; Covington, J.D.; Ravussin, E.; Redman, L.M. Little evidence of systemic and adipose tissue inflammation in overweight individuals(dagger). Front. Genet. 2012, 3, 58. [Google Scholar] [CrossRef] [Green Version]
- Klimcakova, E.; Polak, J.; Moro, C.; Hejnova, J.; Majercik, M.; Viguerie, N.; Berlan, M.; Langin, D.; Stich, V. Dynamic strength training improves insulin sensitivity without altering plasma levels and gene expression of adipokines in subcutaneous adipose tissue in obese men. J. Clin. Endocrinol. Metab. 2006, 91, 5107–5112. [Google Scholar] [CrossRef] [Green Version]
- Polak, J.; Klimcakova, E.; Moro, C.; Viguerie, N.; Berlan, M.; Hejnova, J.; Richterova, B.; Kraus, I.; Langin, D.; Stich, V. Effect of aerobic training on plasma levels and subcutaneous abdominal adipose tissue gene expression of adiponectin, leptin, interleukin 6, and tumor necrosis factor alpha in obese women. Metabolism 2006, 55, 1375–1381. [Google Scholar] [CrossRef]
- Forsythe, L.K.; Wallace, J.M.; Livingstone, M.B. Obesity and inflammation: The effects of weight loss. Nutr. Res. Rev. 2008, 21, 117–133. [Google Scholar] [CrossRef]
- Magkos, F.; Fraterrigo, G.; Yoshino, J.; Luecking, C.; Kirbach, K.; Kelly, S.C.; de Las, F.L.; He, S.; Okunade, A.L.; Patterson, B.W.; et al. Effects of Moderate and Subsequent Progressive Weight Loss on Metabolic Function and Adipose Tissue Biology in Humans with Obesity. Cell Metab. 2016, 23, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holness, C.L.; Simmons, D.L. Molecular cloning of CD68, a human macrophage marker related to lysosomal glycoproteins. Blood 1993, 81, 1607–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odegaard, J.I.; Ricardo-Gonzalez, R.R.; Goforth, M.H.; Morel, C.R.; Subramanian, V.; Mukundan, L.; Red, E.A.; Vats, D.; Brombacher, F.; Ferrante, A.W.; et al. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature 2007, 447, 1116–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumeng, C.N.; Delproposto, J.B.; Westcott, D.J.; Saltiel, A.R. Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes 2008, 57, 3239–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, K.; Parker, J.L.; Ouchi, N.; Higuchi, A.; Vita, J.A.; Gokce, N.; Pedersen, A.A.; Kalthoff, C.; Tullin, S.; Sams, A.; et al. Adiponectin promotes macrophage polarization toward an anti-inflammatory phenotype. J. Biol. Chem. 2010, 285, 6153–6160. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Liu, M. Adiponectin: A versatile player of innate immunity. J. Mol. Cell Biol. 2016, 8, 120–128. [Google Scholar] [CrossRef]
- You, T.; Wang, X.; Murphy, K.M.; Lyles, M.F.; Demons, J.L.; Yang, R.; Gong, D.W.; Nicklas, B.J. Regional adipose tissue hormone/cytokine production before and after weight loss in abdominally obese women. Obesity (Silver Spring) 2014, 22, 1679–1684. [Google Scholar] [CrossRef] [Green Version]
- Fisher, F.M.; McTernan, P.G.; Valsamakis, G.; Chetty, R.; Harte, A.L.; Anwar, A.J.; Starcynski, J.; Crocker, J.; Barnett, A.H.; McTernan, C.L.; et al. Differences in adiponectin protein expression: Effect of fat depots and type 2 diabetic status. Horm. Metab. Res. 2002, 34, 650–654. [Google Scholar] [CrossRef]
- Lihn, A.S.; Bruun, J.M.; He, G.; Pedersen, S.B.; Jensen, P.F.; Richelsen, B. Lower expression of adiponectin mRNA in visceral adipose tissue in lean and obese subjects. Mol. Cell Endocrinol. 2004, 219, 9–15. [Google Scholar] [CrossRef]



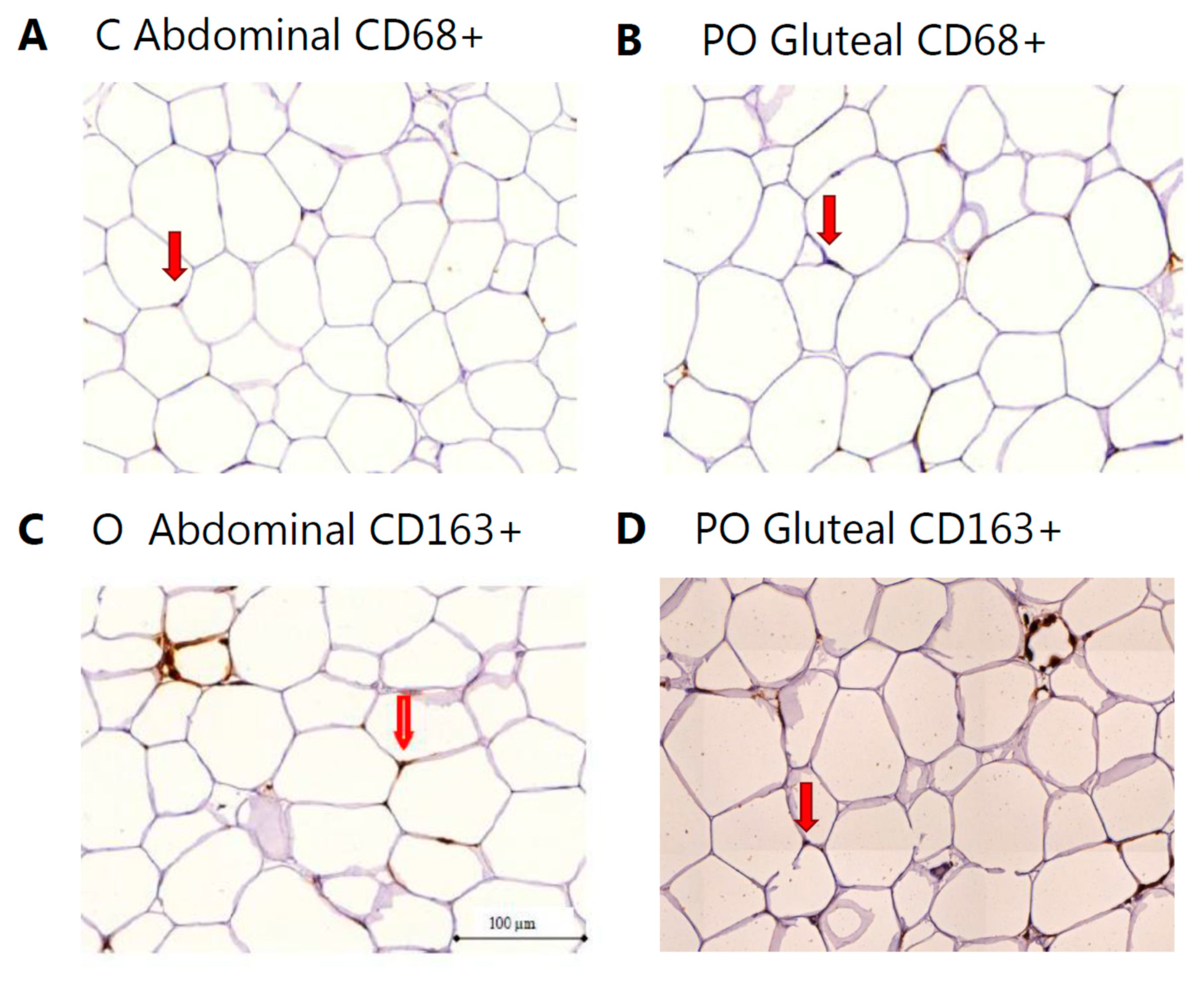{kind=link}
{kind=link}
| POST OBESE (n = 10) | OBESE (n = 10) | CONTROL (n = 10) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Age (years) | 31.5 | ± | 1.6 | 30.4 | ± | 2.3 | 31.2 | ± | 1.5 |
| Height (cm) | 184.0 | ± | 1.7 | 183.9 | ± | 2.6 | 184.3 | ± | 3.0 |
| Weight (kg) | 90.2 | ± | 3.1 * | 115.0 | ± | 5.4 | 90.9 | ± | 4.2 * |
| BMI (kg/m2) | 26.6 | ± | 0.7 * | 33.8 | ± | 1.0 | 26.6 | ± | 0.6 * |
| Body fat (%) | 22.8 | ± | 1.8 * | 34.9 | ± | 1.6 | 24.7 | ± | 1.8 * |
| LBM (kg) | 66.8 | ± | 1.5 | 69.2 | ± | 3.2 | 65.5 | ± | 1.9 |
| VO2peak (L/min) | 3.7 | ± | 0.1 | 3.4 | ± | 0.2 | 3.6 | ± | 0.1 |
| QUICKI | 0.38 | ± | 0.01 * | 0.33 | ± | 0.01 | 0.37 | ± | 0.01 * |
| POST OBESE (n = 9) | OBESE (n = 10) | CONTROL (n = 10) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Plasma | |||||||||
| TNF-α (pg/mL) | 1.28 | ± | 0.16 * | 1.79 | ± | 0.08 | 1.39 | ± | 0.13 |
| IL-18 (pg/mL) | 327 | ± | 34 | 469 | ± | 50 | 389 | ± | 40 |
| IL-6 (pg/mL) | 0.98 | ± | 0.32 * | 1.66 | ± | 0.24 | 0.92 | ± | 0.22 * |
| CRP (mg/L) | 0.76 | ± | 0.17 * | 3.84 | ± | 1.04 | 1.92 | ± | 1.01 * |
| Adipocyte Area | |||||||||
| Abdominal depot (103 µm2) | 10.2 | ± | 0.7 | 12.8 | ± | 0.7 | 11.2 | ± | 0.7 |
| Gluteal depot (103 µm2) | 10.0 | ± | 0.9 | 12.1 | ± | 0.8 | 10.5 | ± | 0.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ara, I.; Auerbach, P.; Larsen, S.; Mata, E.; Stallknecht, B.; Ploug, T.; Prats, C.; Helge, J.W. Low-Grade Inflammation Is Not Present in Former Obese Males but Adipose Tissue Macrophage Infiltration Persists. Biomedicines 2020, 8, 123. https://doi.org/10.3390/biomedicines8050123
Ara I, Auerbach P, Larsen S, Mata E, Stallknecht B, Ploug T, Prats C, Helge JW. Low-Grade Inflammation Is Not Present in Former Obese Males but Adipose Tissue Macrophage Infiltration Persists. Biomedicines. 2020; 8(5):123. https://doi.org/10.3390/biomedicines8050123
Chicago/Turabian StyleAra, Ignacio, Pernille Auerbach, Steen Larsen, Esmeralda Mata, Bente Stallknecht, Thorkil Ploug, Clara Prats, and Jørn W. Helge. 2020. "Low-Grade Inflammation Is Not Present in Former Obese Males but Adipose Tissue Macrophage Infiltration Persists" Biomedicines 8, no. 5: 123. https://doi.org/10.3390/biomedicines8050123