Atropine Differentially Modulates ECM Production by Ocular Fibroblasts, and Its Ocular Surface Toxicity Is Blunted by Colostrum

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. ECM Proteins Analysis by Western Blotting
2.3. Cell Survival with Atropine
3. Results
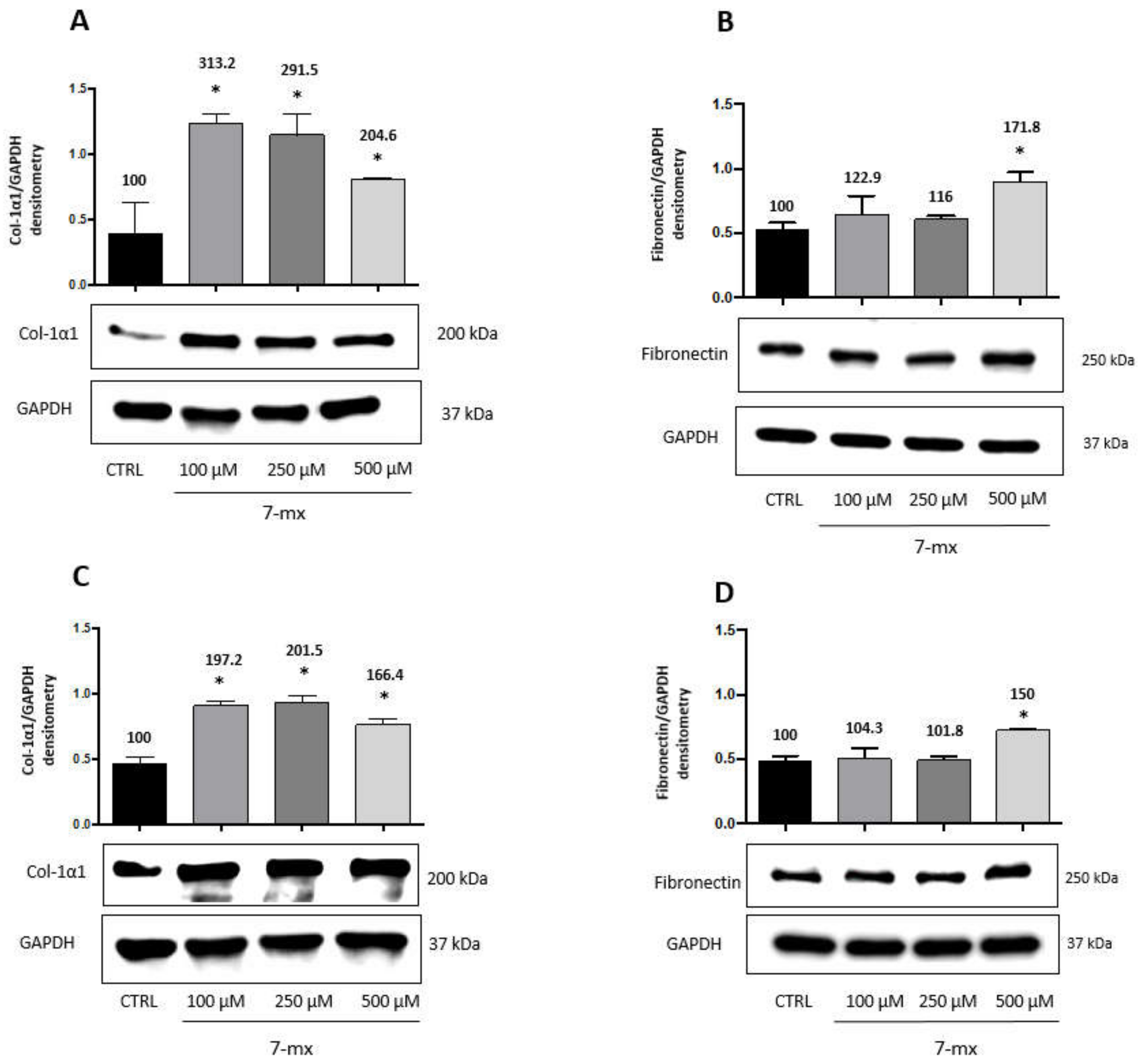
3.1. ECM Production in Scleral Fibroblasts
3.2. ECM Production in Choroidal Fibroblasts
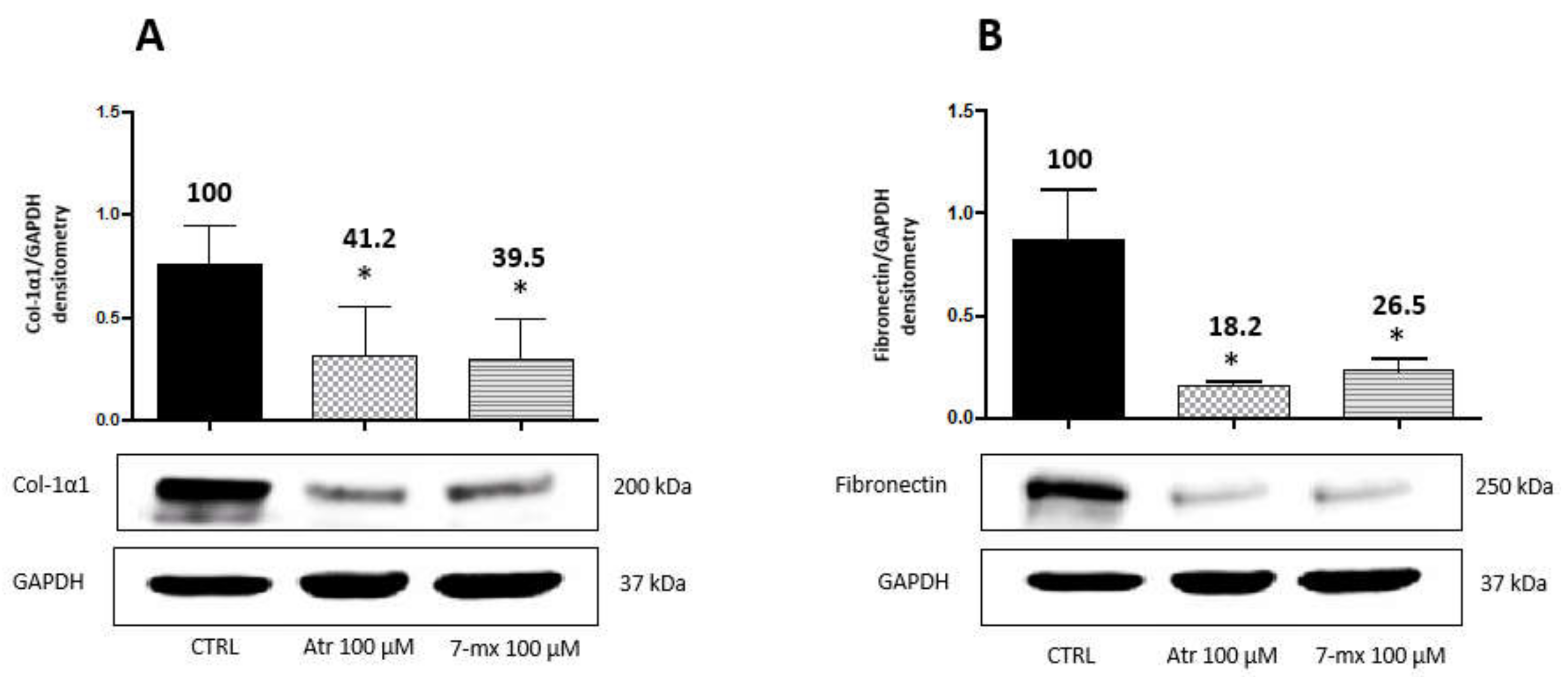
3.3. Atropine Toxicity Is Blunted by Colostrum and Fucosyl-Lactose
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moriyama, M.; Ohno-Matsui, K.; Hayashi, K.; Shimada, N.; Yoshida, T.; Tokoro, T.; Morita, I. Topographic Analyses of Shape of Eyes with Pathologic Myopia by High-Resolution Three-Dimensional Magnetic Resonance Imaging. Ophthalmology 2011, 118, 1626–1637. [Google Scholar] [CrossRef] [PubMed]
- Pineles, S.L.; Kraker, R.T.; VanderVeen, D.K.; Hutchinson, A.K.; Galvin, J.A.; Wilson, L.B.; Lambert, S.R. Atropine for the Prevention of Myopia Progression in Children. Ophthalmology 2017, 124, 1857–1866. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Janowski, M.; Luo, M.; Wei, H.; Chen, B.; Yang, G.; Liu, L. Efficacy and Adverse Effects of Atropine in Childhood Myopia: A Meta-analysis. JAMA Ophthalmol. 2017, 1, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Zhou, X.; Xie, R.; Zhang, L.; Hu, D.; Li, H.; Lu, F. The Presence of m1 to m5 Receptors in Human Sclera: Evidence of the Sclera as a Potential Site of Action for Muscarinic Receptor Antagonists. Curr. Eye Res. 2006, 31, 587–597. [Google Scholar] [CrossRef] [PubMed]
- McBrien, N.A.; Jobling, A.I.; Gentle, A. Biomechanics of the Sclera in Myopia: Extracellular and Cellular Factors. Optom. Vis. Sci. 2009, 86, E23–E30. [Google Scholar] [CrossRef] [PubMed]
- Metlapally, R.; Wildsoet, C.F. Scleral Mechanisms Underlying Ocular Growth and Myopia. Progress Mol. Biol. Transl. Sci. 2015, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Rabinowitz, Y.S. Keratoconus. Surv. Ophthalmol. 1998, 42, 297–319. [Google Scholar] [CrossRef]
- Yokoi, T.; Jonas, J.B.; Shimada, N.; Nagaoka, N.; Moriyama, M.; Yoshida, T.; Ohno-Matsui, K. Peripapillary Diffuse Chorioretinal Atrophy in Children as a Sign of Eventual Pathologic Myopia in Adults. Ophthalmology 2016, 123, 1783–1787. [Google Scholar] [CrossRef]
- Wu, H.; Chen, W.; Zhao, F.; Zhou, Q.; Reinach, P.S.; Deng, L.; Ma, L.; Luo, S.; Srinivasalu, N.; Pan, M.; et al. Scleral hypoxia is a target for myopia control. Proc. Natl. Acad. Sci. USA 2018, 115, E7091–E7100. [Google Scholar] [CrossRef] [Green Version]
- Trier, K.; Munk Ribel-Madsen, S.; Cui, D.; Brøgger Christensen, S. Systemic 7-methylxanthine in retarding axial eye growth and myopia progression: A 36-month pilot study. J. Ocul. Biol. Dis. Inf. 2008, 1, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Schaeffel, F.; Trier, K.; Feldkaemper, M. Effects of 7-Methylxanthine on Deprivation Myopia and Retinal Dopamine Release in Chickens. Ophthalmic Res. 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Ma, J. Will the long-term use of atropine eye drops in children increase the risk of dry eye? Med. Hypotheses 2019, 132, 109331. [Google Scholar] [CrossRef] [PubMed]
- Anfuso, C.D.; Olivieri, M.; Fidilio, A.; Lupo, G.; Rusciano, D.; Pezzino, S.; Gagliano, C.; Drago, F.; Bucolo, C. Gabapentin Attenuates Ocular Inflammation: In vitro and In vivo Studies. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef]
- Hung, L.-F.; Arumugam, B.; Ostrin, L.; Patel, N.; Trier, K.; Jong, M.; Ehlers, L.S. The Adenosine Receptor Antagonist, 7-Methylxanthine, Alters Emmetropizing Responses in Infant Macaques. Investig. Opthalmol. Vis. Sci. 2018, 59, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagge, A.; Ferro Desideri, L.; Nucci, P.; Serafino, M.; Giannaccare, G.; Traverso, C. Prevention of Progression in Myopia: A Systematic Review. Diseases 2018, 6, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katona, M.; Vizvári, E.; Németh, L.; Facskó, A.; Venglovecz, V.; Rakonczay, Z.; Hegyi, P.; Tóth-Molnár, E. Experimental Evidence of Fluid Secretion of Rabbit Lacrimal Gland Duct Epithelium. Investig. Opthalmol. Vis. Sci. 2014, 55, 4360. [Google Scholar] [CrossRef] [Green Version]
- Bucolo, C.; Musumeci, M.; Salomone, S.; Romano, G.L.; Leggio, G.M.; Gagliano, C.; Reibaldi, M.; Avitabile, T.; Uva, M.G.; Musumeci, S.; et al. Effects of Topical Fucosyl-Lactose, a Milk Oligosaccharide, on Dry Eye Model: An Example of Nutraceutical Candidate. Front. Pharmacol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Edwards, N. Myopia: Prevalence and Progression; National Academy Press: Washington, DC, USA, 1989; p. 113. [Google Scholar]
- Tkatchenko, T.V.; Troilo, D.; Benavente-Perez, A.; Tkatchenko, A.V. Gene expression in response to optical defocus of opposite signs reveals bidirectional mechanism of visually guided eye growth. PLOS Biol. 2018, 16, e2006021. [Google Scholar] [CrossRef] [Green Version]
- Read, S.A.; Fuss, J.A.; Vincent, S.J.; Collins, M.J.; Alonso-Caneiro, D. Choroidal changes in human myopia: Insights from optical coherence tomography imaging. Clin. Exp. Optom. 2018, 102, 270–285. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Pardue, M.T.; Iuvone, P.M.; Qu, J. Dopamine signaling and myopia development: What are the key challenges. Progress Retin. Eye Res. 2017, 61, 60–71. [Google Scholar] [CrossRef]
- Meng, B.; Li, S.-M.; Yang, Y.; Yang, Z.-R.; Sun, F.; Kang, M.-T.; Sun, Y.-Y.; Ran, A.-R.; Wang, J.-N.; Yan, R.; et al. The association of TGFB1 genetic polymorphisms with high myopia: A systematic review and meta-analysis. Int. J. Clin. Exp. Med. 2015, 8, 20355–20367. [Google Scholar] [PubMed]
- Kearney, S.; O’Donoghue, L.; Pourshahidi, L.K.; Cobice, D.; Saunders, K.J. Myopes have significantly higher serum melatonin concentrations than non-myopes. Ophthalmic Physiol. Opt. 2017, 37, 557–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Wu, S.; Wang, Y.; Pan, S.; Wang, P.; Cheng, L. Inflammatory cytokines in highly myopic eyes. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, T.T.; Miller, E.J. Collagen and protein levels in sclera during normal development, induced myopia, and recovery in tree shrews. Invest. Ophthalmol. Vis. Sci. 1995, 36, S760. [Google Scholar]
- Gentle, A.; Liu, Y.; Martin, J.E.; Conti, G.L.; McBrien, N.A. Collagen Gene Expression and the Altered Accumulation of Scleral Collagen during the Development of High Myopia. J. Biol. Chem. 2003, 278, 16587–16594. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.; Liu, R.; Zhang, X.; Chu, R.; Dai, J.; Zhou, H.; Liu, H. Upregulation of regulator of G-protein signaling 2 in the sclera of a form deprivation myopic animal model. Mol. Vis. 2014, 20, 977–987. [Google Scholar]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Miller, C.G.; Budoff, G.; Prenner, J.L.; Schwarzbauer, J.E. Minireview: Fibronectin in retinal disease. Exp. Biol. Med. 2016, 242, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jonas, J.B.; Ohno-Matsui, K.; Jiang, W.J.; Panda-Jonas, S. Bruch membrane and the mechanism of myopization. Retina 2017, 37, 1428–1440. [Google Scholar] [CrossRef]
- Beach, K.M.; Hung, L.-F.; Arumugam, B.; Smith, E.L.; Ostrin, L.A. Adenosine receptor distribution in Rhesus monkey ocular tissue. Exp. Eye Res. 2018, 174, 40–50. [Google Scholar] [CrossRef]
- McBrien, N.A.; Jobling, A.I.; Truong, H.T.; Cottriall, C.L.; Gentle, A. Expression of muscarinic receptor subtypes in tree shrew ocular tissues and their regulation during the development of myopia. Mol. Vis. 2009, 15, 464–475. [Google Scholar]
- Vessey, K.A.; Cottriall, C.L.; McBrien, N.A. Muscarinic receptor protein expression in the ocular tissues of the chick during normal and myopic eye development. Dev. Brain Res. 2002, 135, 79–86. [Google Scholar] [CrossRef]
- He, L.; Frost, M.R.; Siegwart, J.T.; Norton, T.T. Gene expression signatures in tree shrew choroid during lens-induced myopia and recovery. Exp. Eye Res. 2014, 123, 56–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhang, G.; Zhou, X.; Xu, R.; Wang, S.; Guan, Z.; Lu, J.; Srinivasalu, N.; Shen, M.; Jin, Z.; et al. Changes in Choroidal Thickness and Choroidal Blood Perfusion in Guinea Pig Myopia. Investig. Opthalmol. Vis. Sci. 2019, 60, 3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Kong, X.; Jiang, C.; Li, M.; Yu, J.; Sun, X. Is the peripapillary retinal perfusion related to myopia in healthy eyes? A prospective comparative study. BMJ Open 2016, 6, e010791. [Google Scholar] [CrossRef] [PubMed]
- Barathi, V.A.; Kwan, J.L.; Tan, Q.S.; Weon, S.R.; Seet, L.F.; Goh, L.K.; Vithana, E.N.; Beuerman, R.W. Muscarinic cholinergic receptor (M2) plays a crucial role in the development of myopia in mice. Dis. Model Mech. 2013, 6, 1146–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Li, J.; Tan, D.T.; Beuerman, R.W. Expression and function of muscarinic receptor subtypes on human cornea and conjunctiva. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2987–2996. [Google Scholar] [CrossRef] [PubMed]
- De Ligt, R.A.; Kourounakis, A.P.; IJzerman, A.P. Inverse agonism at G protein-coupled receptors: (patho)physiological relevance and implications for drug discovery. Br. J. Pharmacol. 2000, 130, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kinzer-Ursem, T.L.; Linderman, J.J. Both ligand- and cell-specific parameters control ligand agonism in a kinetic model of g protein-coupled receptor signaling. PLoS Comput. Biol. 2007, 3, e6. [Google Scholar] [CrossRef] [Green Version]
- Vutipongsatorn, K.; Yokoi, T.; Ohno-Matsui, K. Current and emerging pharmaceutical interventions for myopia. Br. J. Ophthalmol. 2019, 103, 1539–1548. [Google Scholar] [CrossRef]
- Kumaran, A.; Htoon, H.M.; Tan, D.; Chia, A. Analysis of Changes in Refraction and Biometry of Atropine- and Placebo-Treated Eyes. Investig. Opthalmol. Vis. Sci. 2015, 56, 5650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trier, K. 5. 7-methylxanthine treatment. Acta Ophthalmol. 2015, 93, s25. [Google Scholar] [CrossRef]
- Burgalassi, S.; Panichi, L.; Chetoni, P.; Saettone, M.F.; Boldrini, E. Development of a Simple Dry Eye Model in the Albino Rabbit and Evaluation of Some Tear Substitutes. Ophthalmic Res. 1999, 31, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.-L.; Wen, Q.; Fan, T.-J. Cytotoxicity of atropine to human corneal epithelial cells by inducing cell cycle arrest and mitochondrion-dependent apoptosis. Exp. Toxicol. Pathol. 2015, 67, 517–524. [Google Scholar] [CrossRef] [PubMed]
- García-Martín, E.; Pernía-López, S.; Romero Jiménez, R.; García-Valcárcel, B.; Martínez-Ortega, P.A.; Sanjurjo-Saez, M. The use of autologous serum eye drops for the treatment of ocular surface disorders. Eur. J. Hosp. Pharm. 2018, 26, 314–317. [Google Scholar] [CrossRef] [PubMed]
- Buzzi, M.; Versura, P.; Grigolo, B.; Cavallo, C.; Terzi, A.; Pellegrini, M.; Giannaccare, G.; Randi, V.; Campos, E.C. Comparison of growth factor and interleukin content of adult peripheral blood and cord blood serum eye drops for cornea and ocular surface diseases. Transfus. Apheresis Sci. 2018, 57, 549–555. [Google Scholar] [CrossRef]
- Sacerdote, P.; Mussano, F.; Franchi, S.; Panerai, A.E.; Bussolati, G.; Carossa, S.; Bartorelli, A.; Bussolati, B. Biological components in a standardized derivative of bovine colostrum. J. Dairy Sci. 2013, 96, 1745–1754. [Google Scholar] [CrossRef] [Green Version]
- Elwakiel, M.; Hageman, J.A.; Wang, W.; Szeto, I.M.; van Goudoever, J.B.; Hettinga, K.A.; Schols, H.A. Human Milk Oligosaccharides in Colostrum and Mature Milk of Chinese Mothers: Lewis Positive Secretor Subgroups. J. Agric. Food Chem. 2018, 66, 7036–7043. [Google Scholar] [CrossRef]
- Chaumeil, C.; Liotet, S.; Kogbe, O. Treatment of Severe Eye Dryness and Problematic Eye Lesions with Enriched Bovine Colostrum Lactoserum. Adv. Exp. Med. Biol. 1994, 595–599. [Google Scholar]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cristaldi, M.; Olivieri, M.; Pezzino, S.; Spampinato, G.; Lupo, G.; Anfuso, C.D.; Rusciano, D. Atropine Differentially Modulates ECM Production by Ocular Fibroblasts, and Its Ocular Surface Toxicity Is Blunted by Colostrum. Biomedicines 2020, 8, 78. https://doi.org/10.3390/biomedicines8040078
Cristaldi M, Olivieri M, Pezzino S, Spampinato G, Lupo G, Anfuso CD, Rusciano D. Atropine Differentially Modulates ECM Production by Ocular Fibroblasts, and Its Ocular Surface Toxicity Is Blunted by Colostrum. Biomedicines. 2020; 8(4):78. https://doi.org/10.3390/biomedicines8040078
Chicago/Turabian StyleCristaldi, Martina, Melania Olivieri, Salvatore Pezzino, Giorgia Spampinato, Gabriella Lupo, Carmelina Daniela Anfuso, and Dario Rusciano. 2020. "Atropine Differentially Modulates ECM Production by Ocular Fibroblasts, and Its Ocular Surface Toxicity Is Blunted by Colostrum" Biomedicines 8, no. 4: 78. https://doi.org/10.3390/biomedicines8040078