Experimental Study on Blue Light Interaction with Human Keloid-Derived Fibroblasts

 ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Blue LED Light Device and Its Application on Human Cultured Fibroblasts
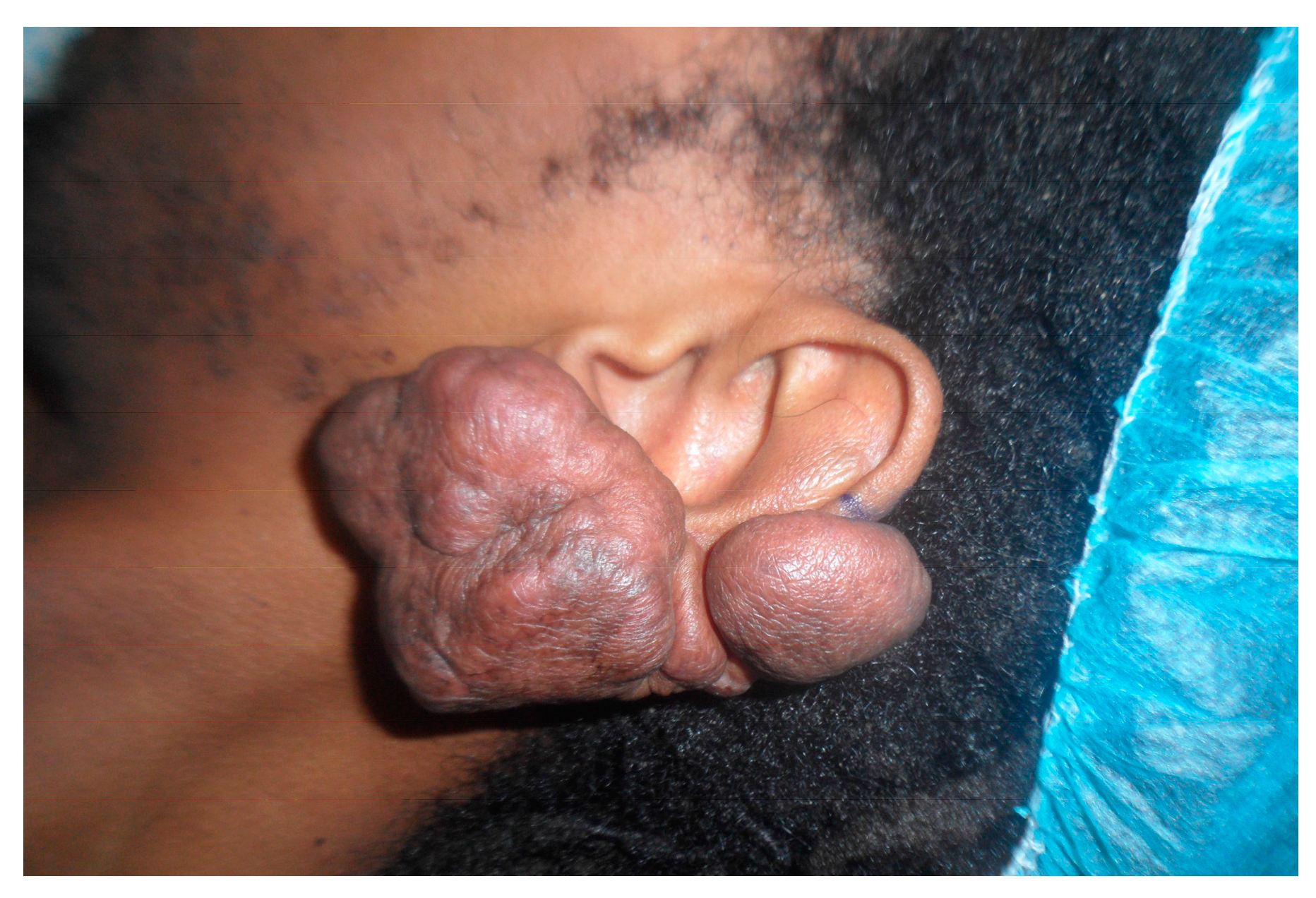
2.2. Human Keloid Samples
2.3. Cell Cultures
2.4. Immunocytochemical Staining
2.5. CCK-8 Assay
2.6. Sulforhodamine B Assay
2.7. Viability Assay
2.8. Electrophysiological Recordings
2.9. Raman Spectroscopy Measurements
2.10. Statistical Analysis
3. Results
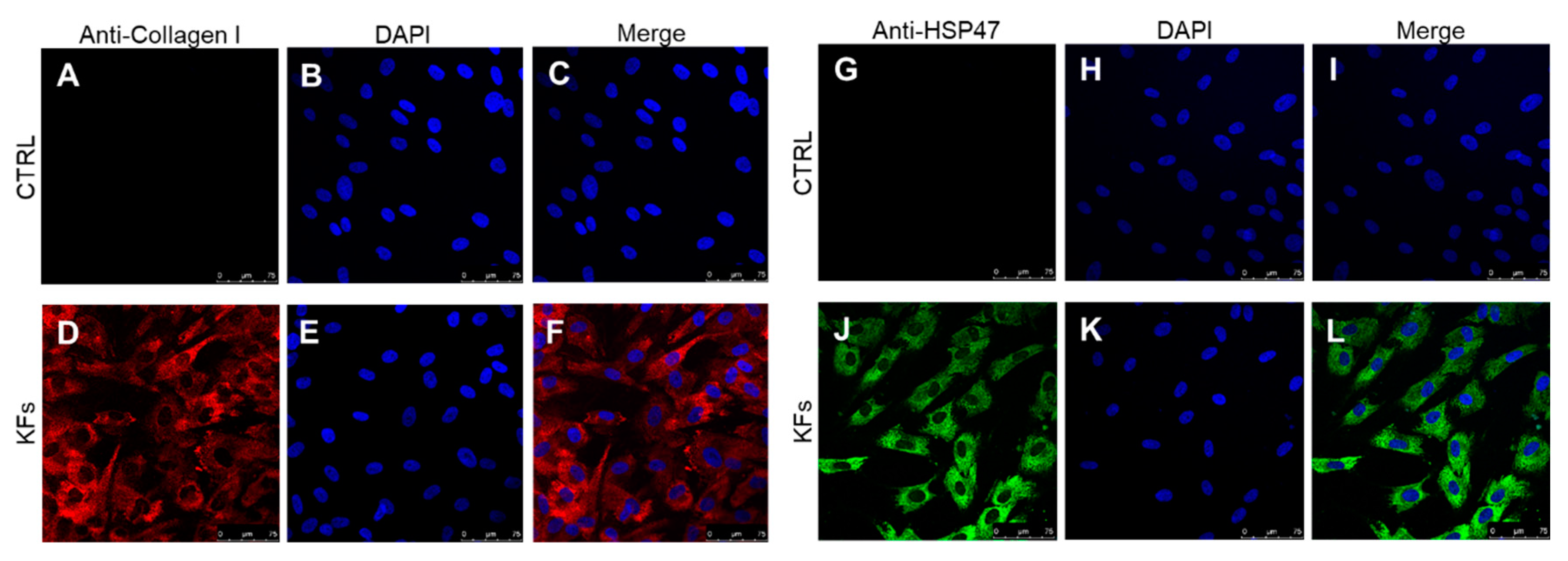
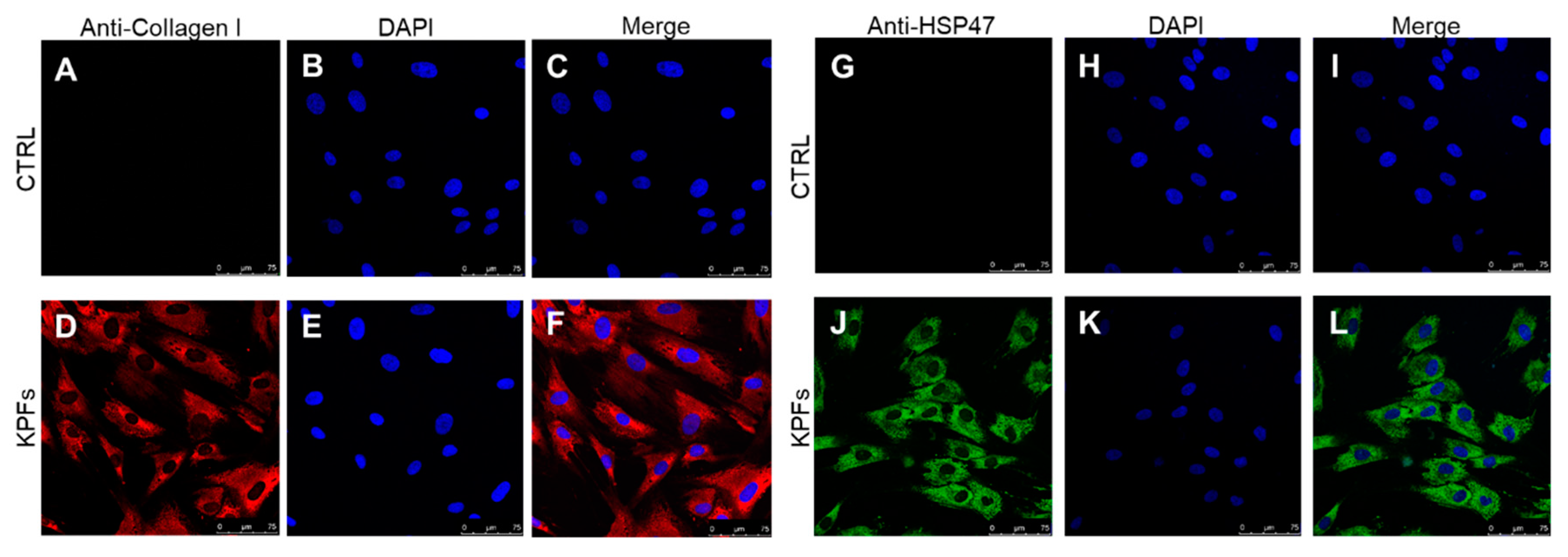
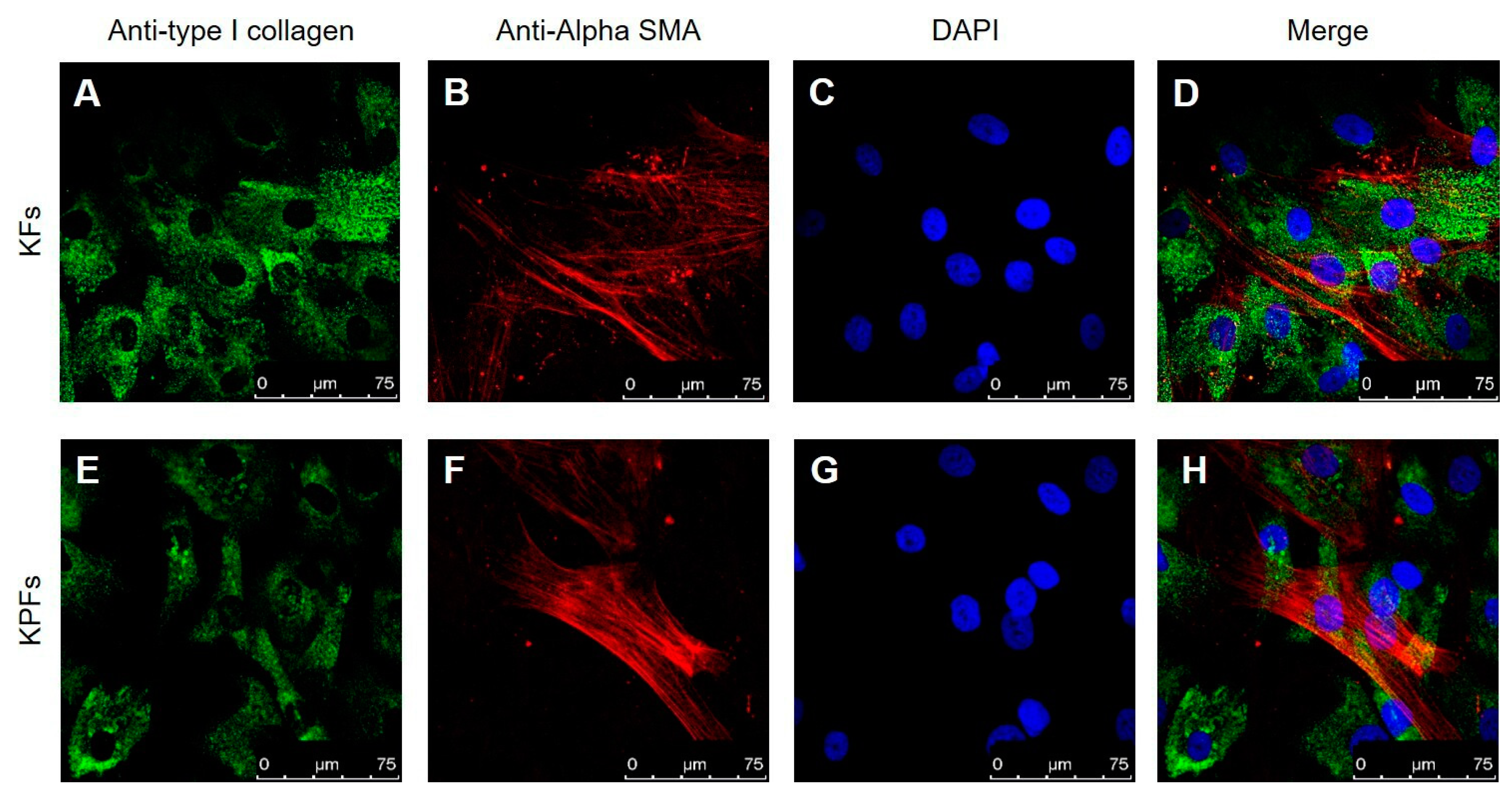
3.1. Characterization of Cell Cultures Isolated from Keloid and Perilesional Tissues by Immunocytochemical Staining
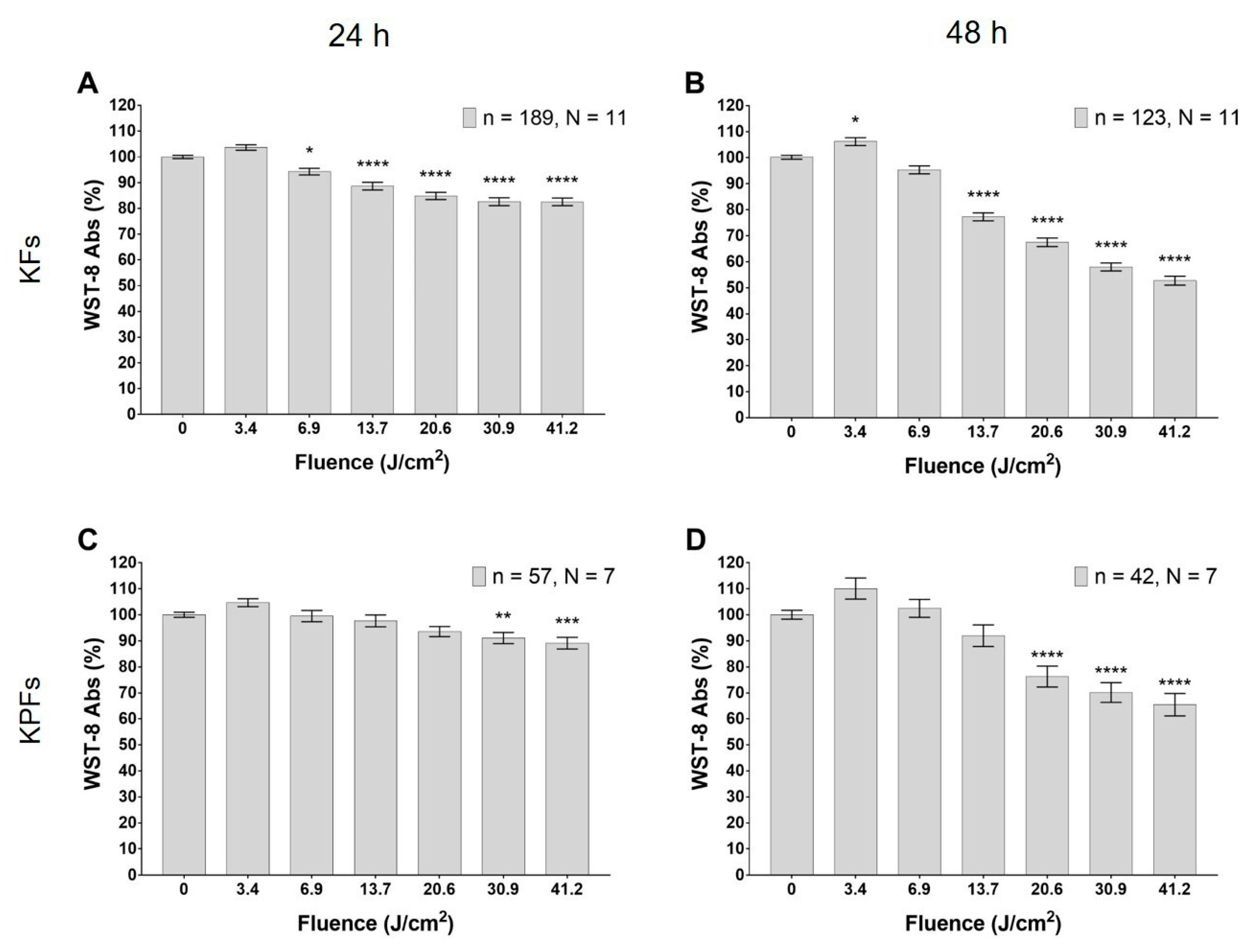
3.2. Blue LED Light Modulates Cell Metabolism in a Dose-Dependent Manner
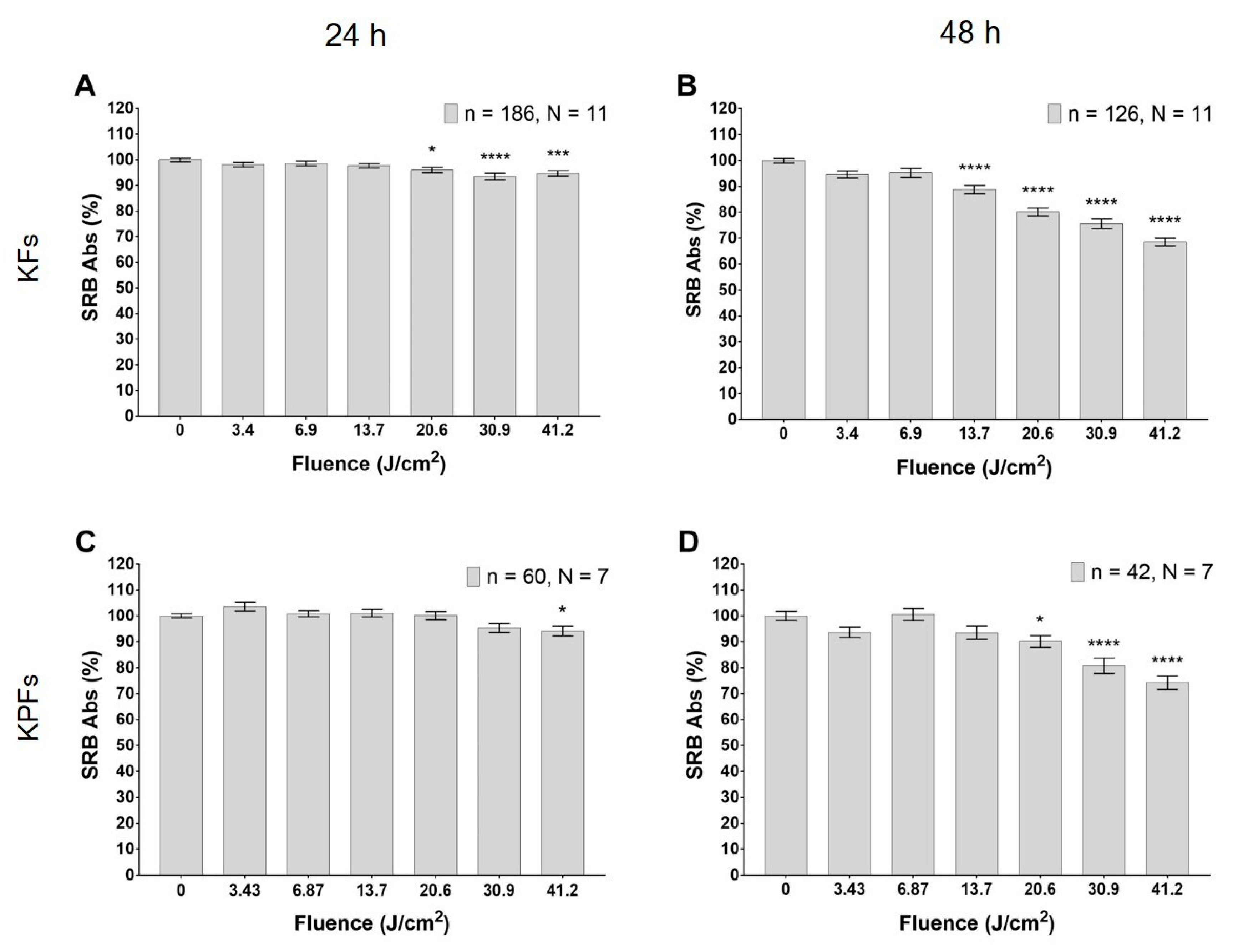
3.3. Blue LED Light Modulates Cell Proliferation in a Dose-Dependent Manner
3.4. High Dose of Blue Led Light Reduces Cells Viability in Fibroblasts Isolated from Keloids and Perilesional Tissues
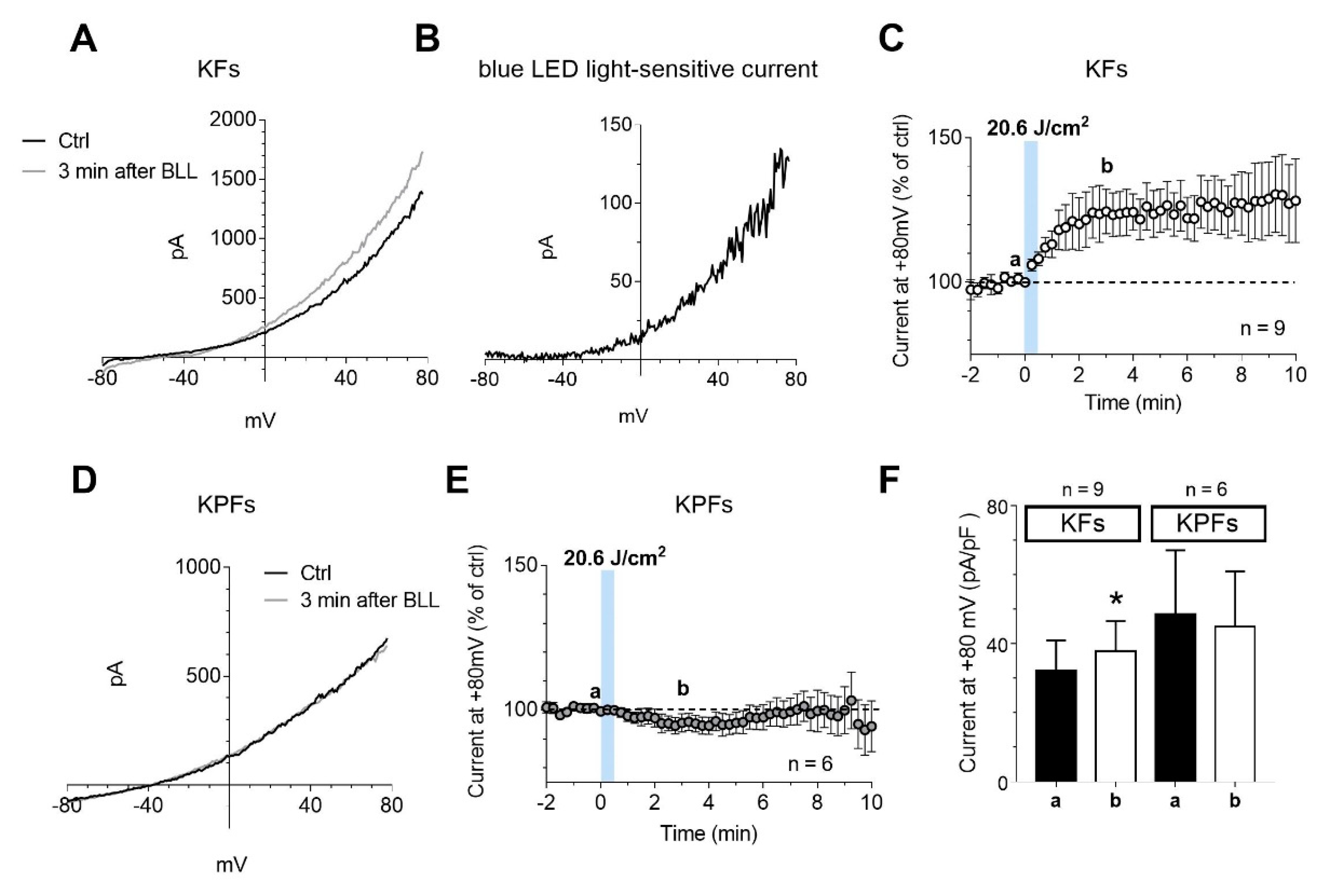
3.5. Blue LED Light Increases the Outward Currents in Fibroblasts Isolated from Keloid Tissues
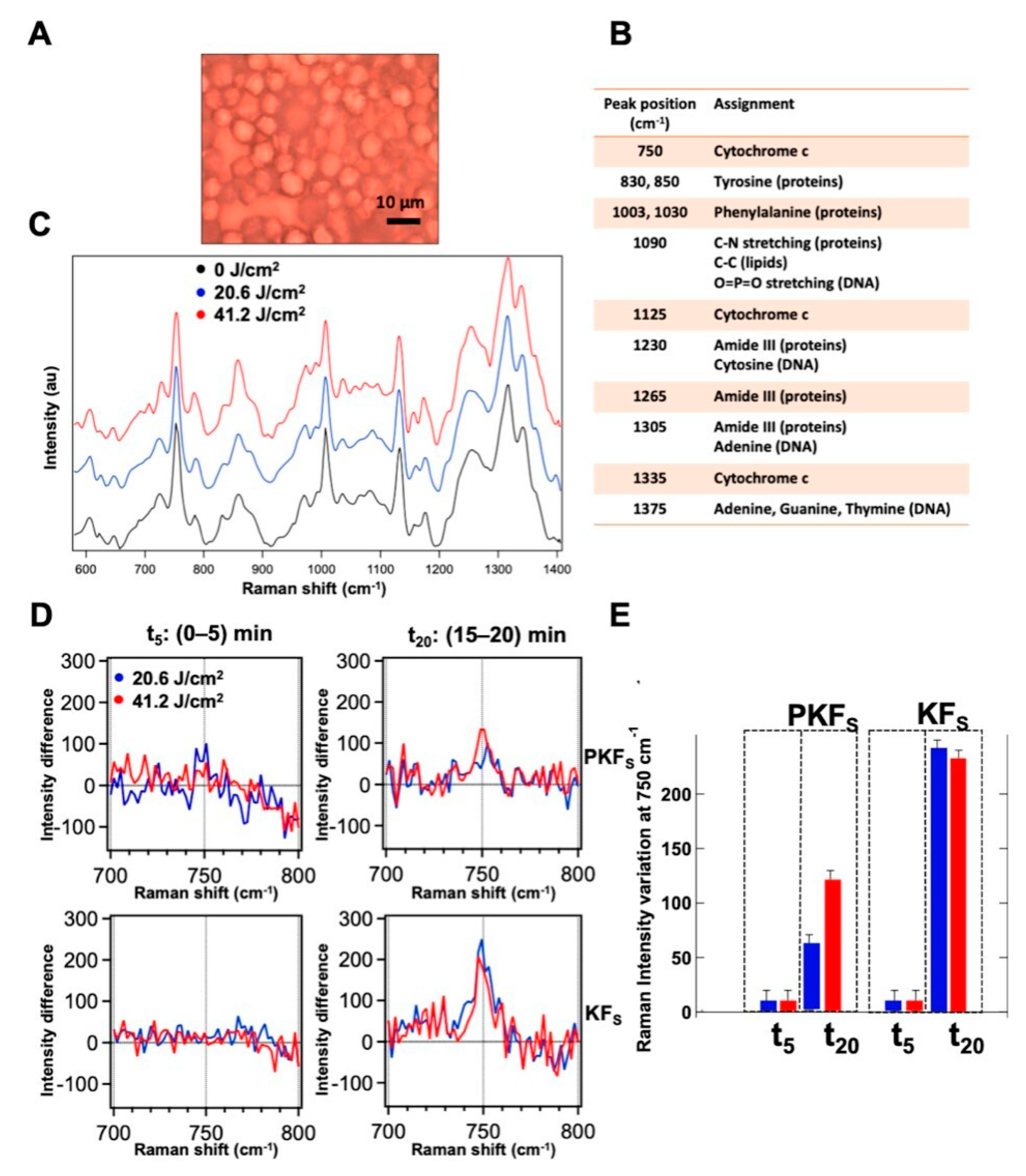
3.6. Modulation of the Redox State of Cytochrome C
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| LED | light emitting diode |
| PBM | Photobiomodulation |
| Cyt C | Cytochrome C |
| ATP | adenosine triphosphate |
| ECM | extracellular matrix |
| KFs | Keloid fibroblast cells |
| KPFs | Keloid Perilesional fibroblast cells |
| DMEM | Dulbecco Modified Eagle Medium |
| FBS | Fetal Serum Bovine |
| RT | Room Temperature |
| PFA | paraformaldehyde |
| BSA | Bovine Serum Albumin |
| HSP-47 | Heat Shock Protein-47 |
| αSMA | Alpha-smooth muscle actin |
| SFM | serum-free medium |
| SRB | Sulforhodamine B based assay |
| CCK-8 | Cell Counting Kit-8 |
| DAPI | 4′,6-diamidino-2-phenylindole |
| GTP | Guanosine triphosphate |
| EGTA | ethylene glycol tetraacetic acid |
| HEPES | 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid |
| EK | potassium equilibrium potential |
| ECl | chlorine equilibrium potential |
References
- Alster, T.S.; Tanzi, E.L. Hypertrophic scars and keloids. Am. J. Clin. Dermatol. 2003, 4, 235–243. [Google Scholar] [CrossRef]
- Gauglitz, G.G.; Korting, H.C.; Pavicic, T.; Ruzicka, T.; Jeschke, M.G. Hypertrophic scarring and keloids: Pathomechanisms and current and emerging treatment strategies. Mol. Med. 2011, 17, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Berman, B.; Maderal, A.; Raphael, B. Keloids and hypertrophic scars: Pathophysiology, classification, and treatment. Dermatol. Surg. 2017, 43, S3–S18. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.Y.; Yang, C.C.; Chao, S.C.; Wong, T.W. Histopathological differential diagnosis of keloid and hypertrophic scar. Am. J. Dermatopathol. 2004, 26, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Jfri, A.; Alajmi, A. Spontaneous keloids: A literature review. Dermatology 2018, 234, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, G.; Luber, A.J. Use of intralesional cryosurgery as an innovative therapy for keloid scars and a review of current treatments. J. Clin. Aesthetic Dermatol. 2013, 6, 23. [Google Scholar]
- Ud-Din, S.; Bayat, A. Strategic management of keloid disease in ethnic skin: A structured approach supported by the emerging literature. Br. J. Dermatol. 2013, 169, 71–81. [Google Scholar] [CrossRef]
- Lee, S.S.; Yosipovitch, G.; Chan, Y.H.; Goh, C.L. Pruritus, pain, and small nerve fiber function in keloids: A controlled study. J. Am. Acad. Dermatol. 2004, 51, 1002–1006. [Google Scholar] [CrossRef]
- Bijlard, E.; Kouwenberg, C.A.; Timman, R.; Hovius, S.E.; Busschbach, J.J.; Mureau, M.A. Burden of keloid disease: A cross-sectional health-related quality of life assessment. Acta Derm. Venereol. 2017, 97, 225–229. [Google Scholar] [CrossRef] [Green Version]
- Betarbet, U.; Blalock, T.W. Keloids: A Review of Etiology, Prevention, and Treatment. J. Clin. Aesthetic Dermatol. 2020, 13, 33. [Google Scholar]
- Supp, D.M. Animal models for studies of keloid scarring. Adv. Wound Care 2019, 8, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Piaggesi, A.; Lauchli, S.; Basselto, F.; Biedermann, T.; Marques, A.; Najati, B.; Palla, H.; Scarpa, C.; Seimetz, D.; Triulzi, I.; et al. Advanced therapies in wound management. J. Wound Care 2018, 27, S6–S137. [Google Scholar] [CrossRef] [PubMed]
- Serrage, H.; Heiskanen, V.; Palin, W.M.; Cooper, P.R.; Milward, M.R.; Hadis, M.; Hamblin, M.R. Under the spotlight: Mechanisms of photobiomodulation concentrating on blue and green light. Photochem. Photobiol. Sci. 2019, 18, 1877–1909. [Google Scholar] [CrossRef] [PubMed]
- Magni, G.; Banchelli, M.; Cherchi, F.; Coppi, E.; Fraccalvieri, M.; Pugliese, A.M.; Pedata, F.; Mangia, A.; Gasperini, S.; Pavone, F.S.; et al. Human keloid cultured fibroblasts irradiated with blue LED light: Evidence from an in vitro study. In European Conference on Biomedical Optics; Optical Society of America: Washington, DC, USA, 2019; p. 11079_31. [Google Scholar]
- Magni, G.; Cherchi, F.; Coppi, E.; Fraccalvieri, M.; Tatini, F.; Fusco, I.; Pini, R.; Pugliese, A.M.; Pedata, F.; Mangia, A.; et al. Blue light effects in human keloid fibroblasts. In Mechanisms of Photobiomodulation Therapy XIV; International Society for Optics and Photonics: Washington, DC, USA, 2019; Volume 10861, p. 1086107. [Google Scholar]
- Keira, S.M.; Ferreira, L.M.; Gragnani, A.; Duarte, I.D.S.; Santos, I.A.N.D. Experimental model for fibroblast culture. Acta Cir. Bras. 2004, 19, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Krassovka, J.; Borgschulze, A.; Sahlender, B.; Lögters, T.; Windolf, J.; Grotheer, V. Blue light irradiation and its beneficial effect on Dupuytren’s fibroblasts. PLoS ONE 2019, 14, e0209833. [Google Scholar] [CrossRef] [Green Version]
- Jo, H.Y.; Kim, Y.; Park, H.W.; Moon, H.E.; Bae, S.; Kim, J.; Kim, D.G.; Paek, S.H. The unreliability of MTT assay in the cytotoxic test of primary cultured glioblastoma cells. Exp. Neurobiol. 2015, 24, 235–245. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. JNCI J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Orellana, E.A.; Kasinski, A.L. Sulforhodamine B (SRB) assay in cell culture to investigate cell proliferation. Bio-Protocol 2016, 6, e1984. [Google Scholar] [CrossRef] [Green Version]
- Coppi, E.; Cherchi, F.; Fusco, I.; Dettori, I.; Gaviano, L.; Magni, G.; Catarzi, D.; Colotta, V.; Varano, F.; Rossi, F.; et al. Adenosine A2B receptors inhibit K+ currents and cell differentiation in cultured oligodendrocyte precursor cells and modulate sphingosine-1-phosphate signaling pathway. Biochem. Pharmacol. 2020, 177, 113956. [Google Scholar] [CrossRef]
- Chertkova, R.V.; Brazhe, N.A.; Bryantseva, T.V.; Nekrasov, A.N.; Dolgikh, D.A.; Yusipovich, A.I.; Sosnovtseva, O.; Maksimov, G.V.; Rubin, A.B.; Kirpichnikov, M.P. New insight into the mechanism of mitochondrial cytochrome c function. PLoS ONE 2017, 12, e0178280. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, K.J.; Syed, F.; Bayat, A. Site-specific keloid fibroblasts alter the behaviour of normal skin and normal scar fibroblasts through paracrine signalling. PLoS ONE 2013, 8, e75600. [Google Scholar] [CrossRef]
- Shin, J.U.; Lee, W.J.; Tran, T.N.; Jung, I.; Lee, J.H. Hsp70 knockdown by siRNA decreased collagen production in keloid fibroblasts. Yonsei Med. J. 2015, 56, 1619–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasab, M.; Batten, M.R.; Bulleid, N.J. Hsp47: A molecular chaperone that interacts with and stabilizes correctly-folded procollagen. EMBO J. 2000, 19, 2204–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verrico, A.; Haylett, A.K.; Moore, J.V. In vivo expression of the collagen-related heat shock protein HSP47, following hyperthermia or photodynamic therapy. Lasers Med. Sci. 2001, 16, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.F.; Tsai, C.H.; Chang, Y.C. The upregulation of heat shock protein 47 expression in human buccal fibroblasts stimulated with arecoline. J. Oral Pathol. Med. 2008, 37, 206–210. [Google Scholar] [CrossRef]
- Hinz, B. Formation and function of the myofibroblast during tissue repair. J. Investig. Dermatol. 2007, 127, 526–537. [Google Scholar] [CrossRef]
- Grasman, J.M.; Williams, M.D.; Razis, C.G.; Bonzanni, M.; Golding, A.S.; Cairns, D.M.; Levin, M.; Kaplan, D.L. Hyperosmolar Potassium Inhibits Myofibroblast Conversion and Reduces Scar Tissue Formation. ACS Biomater. Sci. Eng. 2019, 5, 5327–5336. [Google Scholar] [CrossRef]
- Lee, H.J.; Jang, Y.J. Recent understandings of biology, prophylaxis and treatment strategies for hypertrophic scars and keloids. Int. J. Mol. Sci. 2018, 19, 711. [Google Scholar] [CrossRef] [Green Version]
- Thulabandu, V.; Chen, D.; Atit, R.P. Dermal fibroblast in cutaneous development and healing. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e307. [Google Scholar] [CrossRef]
- Stunova, A.; Vistejnova, L. Dermal fibroblasts—A heterogeneous population with regulatory function in wound healing. Cytokine Growth Factor Rev. 2018, 39, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.P.; Marttala, J.; Macarak, E.; Rosenbloom, J.; Uitto, J. Keloids: The paradigm of skin fibrosis—Pathomechanisms and treatment. Matrix Biol. 2016, 51, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coentro, J.Q.; Pugliese, E.; Hanley, G.; Raghunath, M.; Zeugolis, D.I. Current and upcoming therapies to modulate skin scarring and fibrosis. Adv. Drug Deliv. Rev. 2019, 146, 37–59. [Google Scholar] [CrossRef] [PubMed]
- Mignon, C.; Uzunbajakava, N.E.; Castellano-Pellicena, I.; Botchkareva, N.V.; Tobin, D.J. Differential response of human dermal fibroblast subpopulations to visible and near-infrared light: Potential of photobiomodulation for addressing cutaneous conditions. Lasers Surg. Med. 2018, 50, 859–882. [Google Scholar] [CrossRef]
- Alexiades, M. Laser and light-based treatments of acne and acne scarring. Clin. Dermatol. 2017, 35, 183–189. [Google Scholar] [CrossRef]
- Lebeko, M.; Khumalo, N.P.; Bayat, A. Multi-dimensional models for functional testing of keloid scars: In silico, in vitro, organoid, organotypic, ex vivo organ culture, and in vivo models. Wound Repair Regen. 2019, 27, 298–308. [Google Scholar] [CrossRef]
- Chen, C.H.; Tsai, J.L.; Wang, Y.H.; Lee, C.L.; Chen, J.K.; Huang, M.H. Low-level laser irradiation promotes cell proliferation and mRNA expression of type I collagen and decorin in porcine achilles tendon fibroblasts in vitro. J. Orthop. Res. 2009, 27, 646–650. [Google Scholar] [CrossRef]
- Gavish, L.; Perez, L.; Gertz, S.D. Low-level laser irradiation modulates matrix metalloproteinase activity and gene expression in porcine aortic smooth muscle cells. Lasers Surg. Med. 2006, 38, 779–786. [Google Scholar] [CrossRef]
- Gavish, L.; Perez, L.S.; Reissman, P.; Gertz, S.D. Irradiation with 780 nm Diode Laser Attenuates Inflammatory Cytokines While Upregulating Nitric Oxide in LPS-Stimulated Macrophages: Implications for the Prevention of Aneurysm Progression. In Proceedings of Light-Activated Tissue Regeneration and Therapy Conference; Springer: Berlin/Heidelberg, Germany, 2008; pp. 53–57. [Google Scholar]
- Huang, Y.Y.; Chen, A.C.H.; Sharma, S.K.; Wu, Q.; Hamblin, M.R. Comparison of cellular responses induced by low level light in different cell types. In Mechanisms for Low-Light Therapy V; International Society for Optics and Photonics: Washington, DC, USA, 2010; Volume 7552, p. 75520A. [Google Scholar]
- Kreisler, M.; Christoffers, A.B.; Willershausen, B.; d’Hoedt, B. Effect of low-level GaAlAs laser irradiation on the proliferation rate of human periodontal ligament fibroblasts: An in vitro study. J. Clin. Periodontol. 2003, 30, 353–358. [Google Scholar] [CrossRef]
- Peplow, P.V.; Chung, T.Y.; Baxter, G.D. Laser photobiomodulation of wound healing: A review of experimental studies in mouse and rat animal models. Photomed. Laser Surg. 2010, 28, 291–325. [Google Scholar] [CrossRef]
- Stein, A.; Benayahu, D.; Maltz, L.; Oron, U. Low-level laser irradiation promotes proliferation and differentiation of human osteoblasts in vitro. Photomed. Laser Ther. 2005, 23, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Vinck, E.M.; Cagnie, B.J.; Cornelissen, M.J.; Declercq, H.A.; Cambier, D.C. Increased fibroblast proliferation induced by light emitting diode and low power laser irradiation. Lasers Med. Sci. 2003, 18, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Passarella, S.; Karu, T. Absorption of monochromatic and narrow band radiation in the visible and near IR by both mitochondrial and non-mitochondrial photoacceptors results in photobiomodulation. J. Photochem. Photobiol. B Biol. 2014, 140, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Karu, T. Mitochondrial Mechanisms of Photobiomodulation in Context of New Data about Multiple Roles of ATP; Mary Ann Liebert, Inc.: New Rochelle, NY, USA, 2010. [Google Scholar]
- Quirk, B.J.; Whelan, H.T. Near-Infrared Irradiation Photobiomodulation: The Need for Basic Science; Mary Ann Liebert, Inc.: New Rochelle, NY, USA, 2011. [Google Scholar]
- Baxter, G.D.; Liu, L.; Petrich, S.; Gisselman, A.S.; Chapple, C.; Anders, J.J.; Tumilty, S. Low level laser therapy (Photobiomodulation therapy) for breast cancer-related lymphedema: A systematic review. BMC Cancer 2017, 17, 833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Pauli Paglioni, M.; Araújo, A.L.D.; Arboleda, L.P.A.; Palmier, N.R.; Fonsêca, J.M.; Gomes-Silva, W.; Madrid-Troconis, C.C.; Silveira, F.M.; Martins, M.D.; Faria, K.M.; et al. Tumor safety and side effects of photobiomodulation therapy used for prevention and management of cancer treatment toxicities. A systematic review. Oral Oncol. 2019, 93, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.F.; Zhang, H.; Yuan, X.; Li, J.; Wei, Y.J.; Hu, S.S. In vitro effects of low-level laser irradiation for bone marrow mesenchymal stem cells: Proliferation, growth factors secretion and myogenic differentiation. Lasers Surg. Med. 2008, 40, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Júnior, A.M.R.; Vieira, B.J.; de Andrade, L.C.F.; Aarestrup, F.M. Low-level laser therapy increases transforming growth factor-β 2 expression and induces apoptosis of epithelial cells during the tissue repair process. Photomed. Laser Surg. 2009, 27, 303–307. [Google Scholar] [CrossRef]
- Saygun, I.; Karacay, S.; Serdar, M.; Ural, A.U.; Sencimen, M.; Kurtis, B. Effects of laser irradiation on the release of basic fibroblast growth factor (bFGF), insulin like growth factor-1 (IGF-1), and receptor of IGF-1 (IGFBP3) from gingival fibroblasts. Lasers Med. Sci. 2008, 23, 211–215. [Google Scholar] [CrossRef]
- Hawkins, D.H.; Abrahamse, H. The role of laser fluence in cell viability, proliferation, and membrane integrity of wounded human skin fibroblasts following helium-neon laser irradiation. Lasers Surg. Med. 2006, 38, 74–83. [Google Scholar] [CrossRef]
- Gao, X.; Xing, D. Molecular mechanisms of cell proliferation induced by low power laser irradiation. J. Biomed. Sci. 2009, 16, 4. [Google Scholar] [CrossRef] [Green Version]
- Karu, T. Primary and secondary mechanisms of action of visible to near-IR radiation on cells. J. Photochem. Photobiol. B Biol. 1999, 49, 1–17. [Google Scholar] [CrossRef]
- Karu, T.I. Ten Lectures on Basic Science of Laser Phototherapy; Prima Books AB: Grangesberg, Sweden, 2007. [Google Scholar]
- Opländer, C.; Hidding, S.; Werners, F.B.; Born, M.; Pallua, N.; Suschek, C.V. Effects of blue light irradiation on human dermal fibroblasts. J. Photochem. Photobiol. B Biol. 2011, 103, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Masson-Meyers, D.S.; Bumah, V.V.; Enwemeka, C.S. A comparison of four methods for determining viability in human dermal fibroblasts irradiated with blue light. J. Pharmacol. Toxicol. Methods 2016, 79, 15–22. [Google Scholar] [CrossRef]
- Avola, R.; Graziano, A.C.E.; Pannuzzo, G.; Bonina, F.; Cardile, V. Hydroxytyrosol from olive fruits prevents blue-light-induced damage in human keratinocytes and fibroblasts. J. Cell. Physiol. 2019, 234, 9065–9076. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Park, H.; Ko, J.H.; Lee, W.; Kim, K.; Kim, T.; Shin, J.; Song, J.H.; Noh, Y.H.; Bang, H.; et al. Expression of Ca2+-activated K+ channels in human dermal fibroblasts and their roles in apoptosis. Skin Pharmacol. Physiol. 2010, 23, 91–104. [Google Scholar] [CrossRef]
- Urrego, D.; Tomczak, A.P.; Zahed, F.; Stühmer, W.; Pardo, L.A. Potassium channels in cell cycle and cell proliferation. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130094. [Google Scholar] [CrossRef] [Green Version]
- Mamalis, A.; Garcha, M.; Jagdeo, J. Light emitting diode-generated blue light modulates fibrosis characteristics: Fibroblast proliferation, migration speed, and reactive oxygen species generation. Lasers Surg. Med. 2015, 47, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Jung, S.E.; Kim, S.K.; Kim, Y.S.; Sohn, S.; Kim, Y.C. Low-level light therapy with 410 nm light emitting diode suppresses collagen synthesis in human keloid fibroblasts: An in vitro study. Ann. Dermatol. 2017, 29, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Bonatti, S.; Hochman, B.; Tucci-Viegas, V.M.; Furtado, F.; Pinfildi, C.E.; Pedro, A.C.; Ferreira, L.M. In vitro effect of 470 nm LED (Light Emitting Diode) in keloid fibroblasts. Acta Cir. Bras. 2011, 26, 25–30. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (s) | Fluence (J/cm2) |
|---|---|
| 5 | 3.43 |
| 10 | 6.87 |
| 20 | 13.7 |
| 30 | 20.6 |
| 45 | 30.9 |
| 60 | 41.2 |
| Fitzpatrick Score | Year of Birth/Sex | Anatomical Site/Size (cm) | Surgery |
|---|---|---|---|
| IV | 2004/m | hand/8x4 and perilesional tissue | surgical removal |
| II | 1972/m | helix/1.5 × 1 | cryoexcision |
| II | 1975/f | earlobe/1 × 1 | cryoexcision |
| II | 1987/m | gluteus/6 × 7 and perilesional tissue | surgical removal |
| II | 1987/m | scapula/5 × 5 and perilesional tissue | surgical removal |
| V | 1980/f | auricle/4 × 2 and perilesional tissue | cryoexcision |
| V | 1980/m | helix/12 × 7 | cryoexcision |
| VI | 1989/m | nape/2 × 3.5 and perilesional tissue | cryoexcision |
| V | 1999/m | mandibular/5 × 1.5 and perilesional tissue | cryoexcision |
| V | 1982/m | mandibular/2.5 × 2.5 and perilesional tissue | cryoexcision |
| V | 1977/m | mandibular/8 × 3 and perilesional tissue | cryoexcision |
| Sample | Control | Treated J/cm2 | p Value |
|---|---|---|---|
| Keloid 24 h | 36.55 (17.91) | 37.37 (10.42) | 0.761 |
| Keloid 48 h | 55.83 (17.39) | 40.40 (15.56) | <0.001 |
| Perilesional 24 h | 53.20 (17.94) | 29.62 (11.02) | <0.001 |
| Perilesional 48 h | 53.38 (17.57) | 24.13 (12.91) | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magni, G.; Banchelli, M.; Cherchi, F.; Coppi, E.; Fraccalvieri, M.; Rossi, M.; Tatini, F.; Pugliese, A.M.; Rossi Degl’Innocenti, D.; Alfieri, D.; et al. Experimental Study on Blue Light Interaction with Human Keloid-Derived Fibroblasts. Biomedicines 2020, 8, 573. https://doi.org/10.3390/biomedicines8120573
Magni G, Banchelli M, Cherchi F, Coppi E, Fraccalvieri M, Rossi M, Tatini F, Pugliese AM, Rossi Degl’Innocenti D, Alfieri D, et al. Experimental Study on Blue Light Interaction with Human Keloid-Derived Fibroblasts. Biomedicines. 2020; 8(12):573. https://doi.org/10.3390/biomedicines8120573
Chicago/Turabian StyleMagni, Giada, Martina Banchelli, Federica Cherchi, Elisabetta Coppi, Marco Fraccalvieri, Michele Rossi, Francesca Tatini, Anna Maria Pugliese, Duccio Rossi Degl’Innocenti, Domenico Alfieri, and et al. 2020. "Experimental Study on Blue Light Interaction with Human Keloid-Derived Fibroblasts" Biomedicines 8, no. 12: 573. https://doi.org/10.3390/biomedicines8120573