
Antitumor Efficacy of Arylquin 1 through Dose-Dependent Cytotoxicity, Apoptosis Induction, and Synergy with Radiotherapy in Glioblastoma Models

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Cell Viability Assay
2.3. Cell Migration Assay
2.4. Invasion Assay
2.5. Colony Formation
2.6. Flow Cytometry
2.7. Animal Models
2.8. Data Analysis
3. Results
3.1. Dose-Dependent Cytotoxicity of Arylquin 1 in GBM8401 and A172 Cells
3.2. Induction of Apoptosis and Cell Cycle Arrest by Arylquin 1 in GBM8401 and A172 Cells
3.3. Inhibitory Effect of Arylquin 1 on Invasion and Migration of GBM8401 and A172 Cells
3.4. Synergistic Effects of Arylquin 1 and Radiation on Cell Survival in GBM8401 and A172 Cells
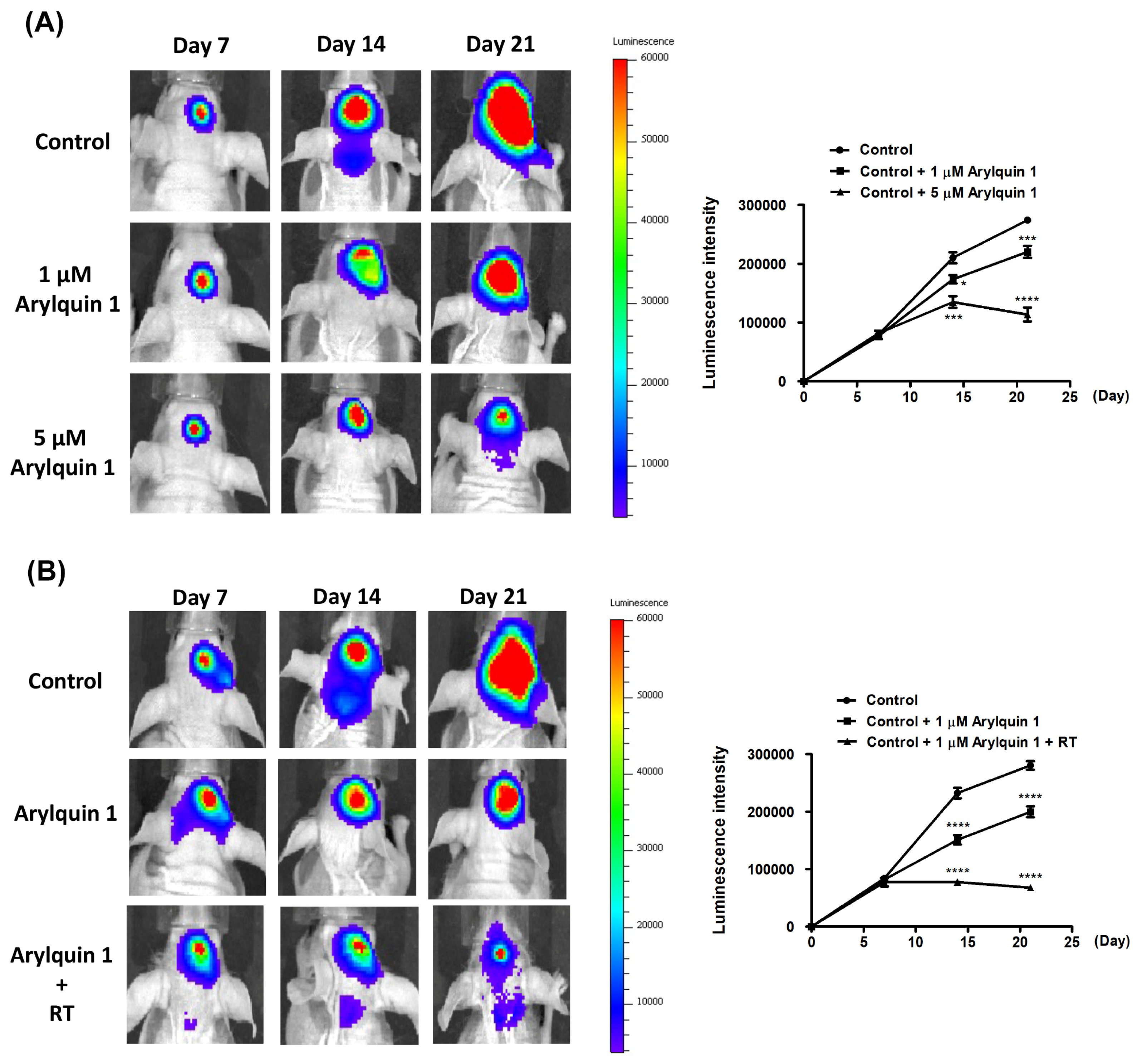
3.5. Synergistic Inhibition of Intracranial Tumor Growth by Arylquin 1 and Radiation Therapy in a Murine Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Price, M.; Neff, C.; Cioffi, G.; Waite, K.A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2016–2020. Neuro-Oncology 2023, 25 (Suppl. S4), iv1–iv99. [Google Scholar] [CrossRef]
- Wirsching, H.G.; Galanis, E.; Weller, M. Glioblastoma. Handb. Clin. Neurol. 2016, 134, 381–397. [Google Scholar]
- Lacroix, M.; Abi-Said, D.; Fourney, D.R.; Gokaslan, Z.L.; Shi, W.; DeMonte, F.; Lang, F.F.; McCutcheon, I.E.; Hassenbusch, S.J.; Holland, E.; et al. A multivariate analysis of 416 patients with glioblastoma multiforme: Prognosis, extent of resection, and survival. J. Neurosurg. 2001, 95, 190–198. [Google Scholar] [CrossRef]
- Lamborn, K.R.; Chang, S.M.; Prados, M.D. Prognostic factors for survival of patients with glioblastoma: Recursive partitioning analysis. Neuro-Oncology 2004, 6, 227–235. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; Farah, P.; Ondracek, A.; Chen, Y.; Wolinsky, Y.; Stroup, N.E.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and central nervous system tumors diagnosed in the United States in 2006–2010. Neuro-Oncology 2013, 15 (Suppl. S2), ii1–ii56. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Cote, D.J.; Ascha, M.; Kruchko, C.; Barnholtz-Sloan, J.S. Adult Glioma Incidence and Survival by Race or Ethnicity in the United States from 2000 to 2014. JAMA Oncol. 2018, 1, 1254–1262. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; Truitt, G.; Boscia, A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2011–2015. Neuro-Oncology 2018, 20 (Suppl. S4), iv1–iv86. [Google Scholar] [CrossRef]
- Burikhanov, R.; Sviripa, V.M.; Hebbar, N.; Zhang, W.; Layton, W.J.; Hamza, A.; Zhan, C.-G.; Watt, D.S.; Liu, C.; Rangnekar, V.M. Arylquins target vimentin to trigger Par-4 secretion for tumor cell apoptosis. Nat. Chem. Biol. 2014, 10, 924–926. [Google Scholar] [CrossRef]
- Sviripa, V.M.; Burikhanov, R.; Obiero, J.M.; Yuan, Y.; Nickell, J.R.; Dwoskin, L.P.; Zhan, C.-G.; Liu, C.; Tsodikov, O.V.; Rangnekar, V.M.; et al. Par-4 secretion: Stoichiometry of 3-arylquinoline binding to vimentin. Org. Biomol. Chem. 2016, 7, 74–84. [Google Scholar] [CrossRef]
- Sells, S.F.; Wood, D.P., Jr.; Joshi-Barve, S.S.; Muthukumar, S.; Jacob, R.J.; Crist, S.A.; Humphreys, S.; Rangnekar, V.M. Commonality of the gene programs induced by effectors of apoptosis in androgen-dependent and -independent prostate cells. Cell Growth Differ. 1994, 5, 457–466. [Google Scholar]
- Chakraborty, M.; Qiu, S.G.; Vasudevan, K.M.; Rangnekar, V.M. Par-4 drives trafficking and activation of Fas and Fasl to induce prostate cancer cell apoptosis and tumor regression. Cancer Res. 2001, 1, 7255–7263. [Google Scholar]
- Burikhanov, R.; Zhao, Y.; Goswami, A.; Qiu, S.; Schwarze, S.R.; Rangnekar, V.M. The tumor suppressor Par-4 activates an extrinsic pathway for apoptosis. Cell 2009, 23, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Greene, J.T.; Mani, R.; Ramaswamy, R.; Frissora, F.; Yano, M.; Zapolnik, K.; Harrington, B.; Wasmuth, R.; Tran, M.; Mo, X.; et al. Par-4 overexpression impedes leukemogenesis in the Eµ-TCL1 leukemia model through downregulation of NF-κB signalling. Blood Adv. 2019, 23, 1255–1266. [Google Scholar] [CrossRef] [PubMed]
- Katoch, A.; Suklabaidya, S.; Chakraborty, S.; Nayak, D.; Rasool, R.U.; Sharma, D.; Mukherjee, D.; Faheem, M.M.; Kumar, A.; Sharma, P.R.; et al. Dual role of Par-4 in abrogation of EMT and switching on Mesenchymal to Epithelial Transition (MET) in metastatic pancreatic cancer cells. Mol. Carcinog. 2018, 57, 1102–1115. [Google Scholar] [CrossRef] [PubMed]
- Rah, B.; Amin, H.; Yousuf, K.; Khan, S.; Jamwal, G.; Mukherjee, D.; Goswami, A. A novel MMP-2 inhibitor 3-azidowithaferin A (3-azidoWA) abrogates cancer cell invasion and angiogenesis by modulating extracellular Par-4. PLoS ONE 2012, 7, e44039. [Google Scholar] [CrossRef]
- Guicciardi, M.E.; Leist, M.; Gores, G.J. Lysosomes in cell death. Oncogene 2004, 12, 2881–2890. [Google Scholar] [CrossRef]
- Boya, P.; Kroemer, G. Lysosomal membrane permeabilization in cell death. Oncogene 2008, 27, 6434–6451. [Google Scholar] [CrossRef]
- Dielschneider, R.F.; Henson, E.S.; Gibson, S.B. Lysosomes as Oxidative Targets for Cancer Therapy. Oxidative Med. Cell. Longev. 2017, 2017, 3749157. [Google Scholar] [CrossRef]
- Goswami, A.; Ranganathan, P.; Rangnekar, V.M. The phosphoinositide 3-kinase/Akt1/Par-4 axis: A cancer-selective therapeutic target. Cancer Res. 2006, 15, 2889–2892. [Google Scholar] [CrossRef]
- Santos, R.V.C.; de Sena, W.L.; dos Santos, F.A.; da Silva Filho, A.F.; da Rocha Pitta, M.G.; da Rocha Pitta, M.G.; de Melo Rego, M.B.; Pereira, M.C. Potential Therapeutic Agents against Par-4 Target for Cancer Treatment: Where Are We Going? Curr. Drug Targets 2019, 20, 635–654. [Google Scholar] [CrossRef]
- Chen, Y.T.; Tseng, T.T.; Tsai, H.P.; Huang, M.Y. Arylquin 1 (Potent Par-4 Secretagogue) Inhibits Tumor Progression and Induces Apoptosis in Colon Cancer Cells. Int. J. Mol. Sci. 2022, 18, 5645. [Google Scholar] [CrossRef] [PubMed]
- Burikhanov, R.; Hebbar, N.; Noothi, S.K.; Shukla, N.; Sledziona, J.; Araujo, N.; Kudrimoti, M.; Wang, Q.J.; Watt, D.S.; Welch, D.R.; et al. Chloroquine-Inducible Par-4 Secretion Is Essential for Tumor Cell Apoptosis and Inhibition of Metastasis. Cell Rep. 2017, 10, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Cheratta, A.R.; Thayyullathil, F.; Pallichankandy, S.; Subburayan, K.; Alakkal, A.; Galadari, S. Prostate apoptosis response-4 and tumor suppression: It’s not just about apoptosis anymore. Cell Death Dis. 2021, 7, 47. [Google Scholar] [CrossRef]
- Azmi, A.S.; Philip, P.A.; Zafar, S.F.; Sarkar, F.H.; Mohammad, R.M. PAR-4 as a possible new target for pancreatic cancer therapy. Expert Opin. Ther. Targets 2010, 14, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Jagtap, J.C.; Dawood, P.; Shah, R.D.; Chandrika, G.; Natesh, K.; Shiras, A.; Hegde, A.S.; Ranade, D.; Shastry, P. Expression and regulation of prostate apoptosis response-4 (Par-4) in human glioma stem cells in drug-induced apoptosis. PLoS ONE 2014, 9, e88505. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gilbert, M.R.; Kyprianou, N.; Rangnekar, V.M.; Horbinski, C. The tumor suppressor prostate apoptosis response-4 (Par-4) is regulated by mutant IDH1 and kills glioma stem cells. Acta Neuropathol. 2014, 128, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, V.; Ghosalkar, J.; Achrekar, S.; Joshi, K. Ketorolac modulates Rac-1/HIF-1alpha/DDX3/beta-catenin signalling via a tumor suppressor prostate apoptosis response-4 (Par-4) in renal cell carcinoma. Sci. Rep. 2023, 13, 5659. [Google Scholar] [CrossRef]
- Alvarez, J.V.; Pan, T.-C.; Ruth, J.; Feng, Y.; Zhou, A.; Pant, D.; Grimley, J.S.; Wandless, T.J.; DeMichele, A.; Chodosh, L.A. Par-4 downregulation promotes breast cancer recurrence by preventing multinucleation following targeted therapy. Cancer Cell 2013, 8, 30–44. [Google Scholar] [CrossRef]
- Irby, R.B.; Kline, C.L. Par-4 as a potential target for cancer therapy. Expert Opin. Ther. Targets 2013, 17, 77–87. [Google Scholar] [CrossRef]
- Guo, H.; Treude, F.; Kramer, O.H.; Luscher, B.; Hartkamp, J. PAR-4 overcomes chemo-resistance in breast cancer cells by antagonizing cIAP1. Sci. Rep. 2019, 19, 8755. [Google Scholar] [CrossRef]
- Lee, A.S. The Par-4-GRP78 TRAIL, more twists and turns. Cancer Biol. Ther. 2009, 8, 2103–2105. [Google Scholar] [CrossRef] [PubMed]
- Shrestha-Bhattarai, T.; Rangnekar, V.M. Cancer-selective apoptotic effects of extracellular and intracellular Par-4. Oncogene 2010, 8, 3873–3880. [Google Scholar] [CrossRef] [PubMed]
- Strouhalova, K.; Prechova, M.; Gandalovicova, A.; Brabek, J.; Gregor, M.; Rosel, D. Vimentin Intermediate Filaments as Potential Target for Cancer Treatment. Cancers 2020, 11, 184. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lieu, A.-S.; Pan, Y.-C.; Lee, J.-H.; Hsieh, Y.-C.; Lin, C.-J.; Hsu, Y.-L.; Chang, K.-C.; Kuo, S.-H.; Tseng, T.-T.; Tsai, H.-P. Antitumor Efficacy of Arylquin 1 through Dose-Dependent Cytotoxicity, Apoptosis Induction, and Synergy with Radiotherapy in Glioblastoma Models. Biomedicines 2024, 12, 907. https://doi.org/10.3390/biomedicines12040907
Lieu A-S, Pan Y-C, Lee J-H, Hsieh Y-C, Lin C-J, Hsu Y-L, Chang K-C, Kuo S-H, Tseng T-T, Tsai H-P. Antitumor Efficacy of Arylquin 1 through Dose-Dependent Cytotoxicity, Apoptosis Induction, and Synergy with Radiotherapy in Glioblastoma Models. Biomedicines. 2024; 12(4):907. https://doi.org/10.3390/biomedicines12040907
Chicago/Turabian StyleLieu, Ann-Shung, Yu-Chi Pan, Jia-Hau Lee, Yuan-Chin Hsieh, Chien-Ju Lin, Ya-Ling Hsu, Kung-Chao Chang, Shih-Hsun Kuo, Tzu-Ting Tseng, and Hung-Pei Tsai. 2024. "Antitumor Efficacy of Arylquin 1 through Dose-Dependent Cytotoxicity, Apoptosis Induction, and Synergy with Radiotherapy in Glioblastoma Models" Biomedicines 12, no. 4: 907. https://doi.org/10.3390/biomedicines12040907