
MARCKS Inhibition Alters Bovine Neutrophil Responses to Salmonella Typhimurium
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Peptide Treatment
2.3. Bovine Neutrophil Isolation
2.4. Pooled Whole Serum Preparation
2.5. Bacterial Strains and Growth Conditions
2.6. Propidium Iodide Staining
2.7. Neutrophil Adhesion
2.8. Chemotaxis
2.9. Phagocytosis—Flow Cytometry
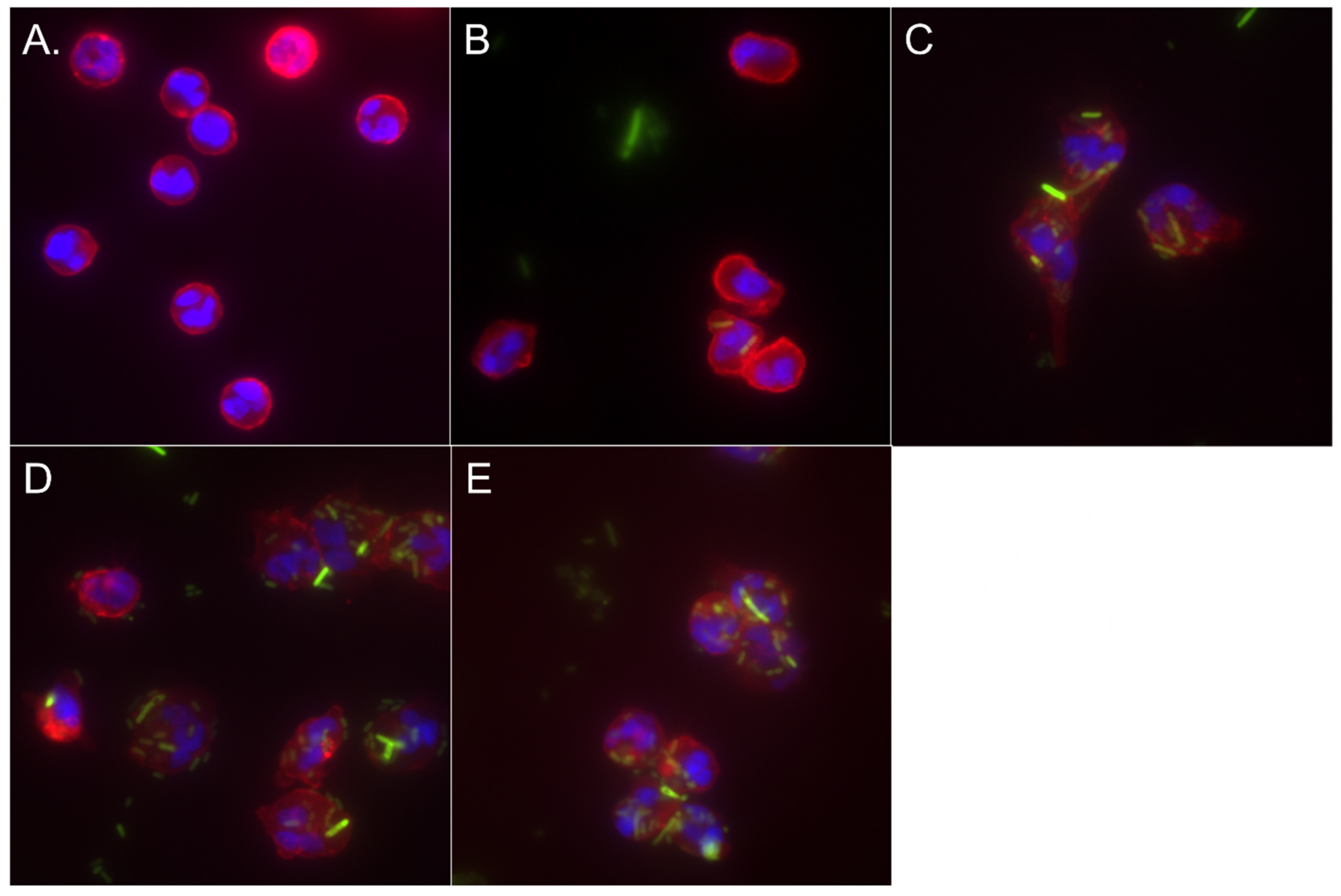
2.10. Phagocytosis—Microscopy
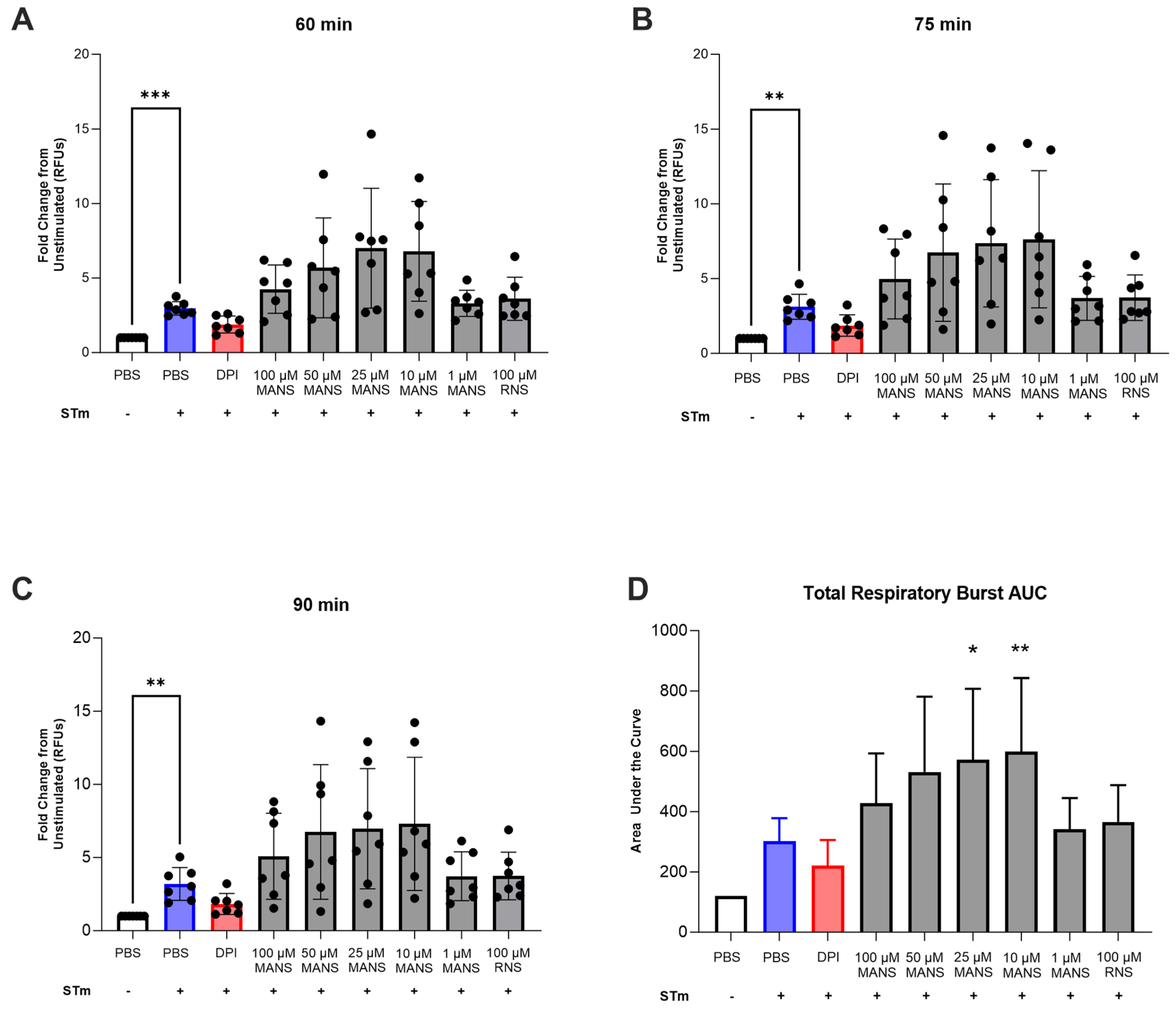
2.11. Respiratory Burst
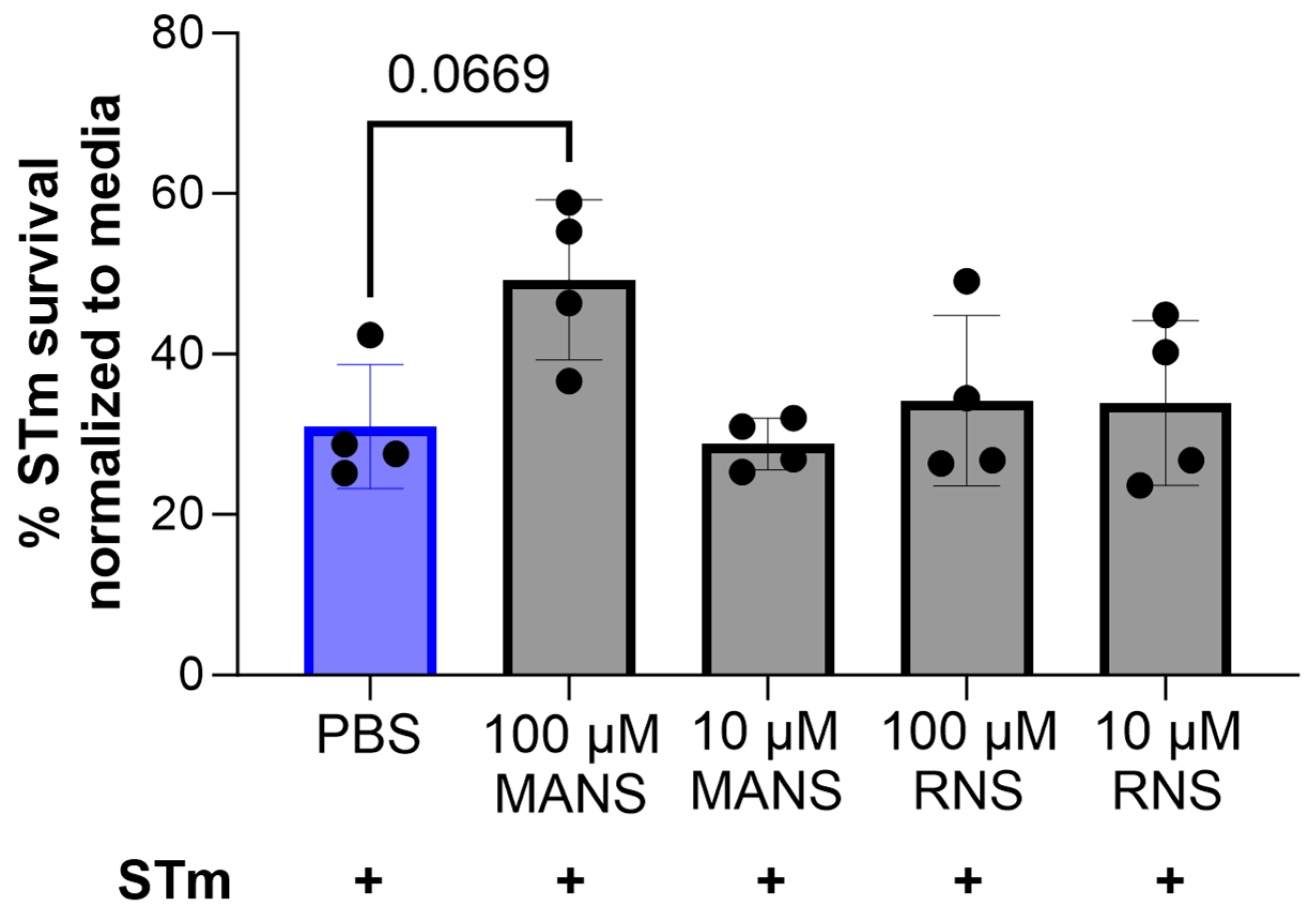
2.12. Salmonella Survival
2.13. Statistical Analyses
3. Results
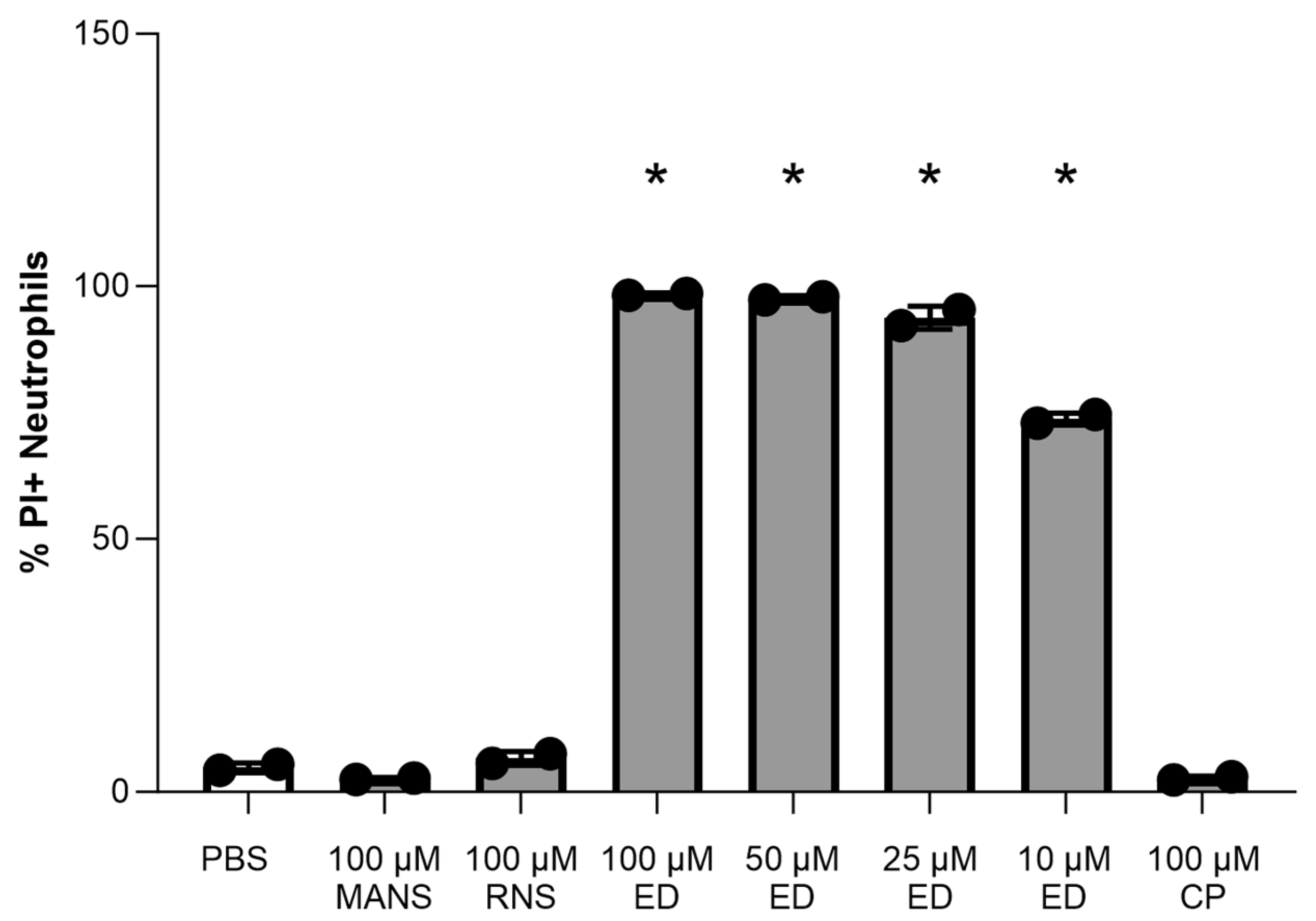
3.1. Effect of MARCKS Inhibitor Peptides on Primary Neutrophil Viability
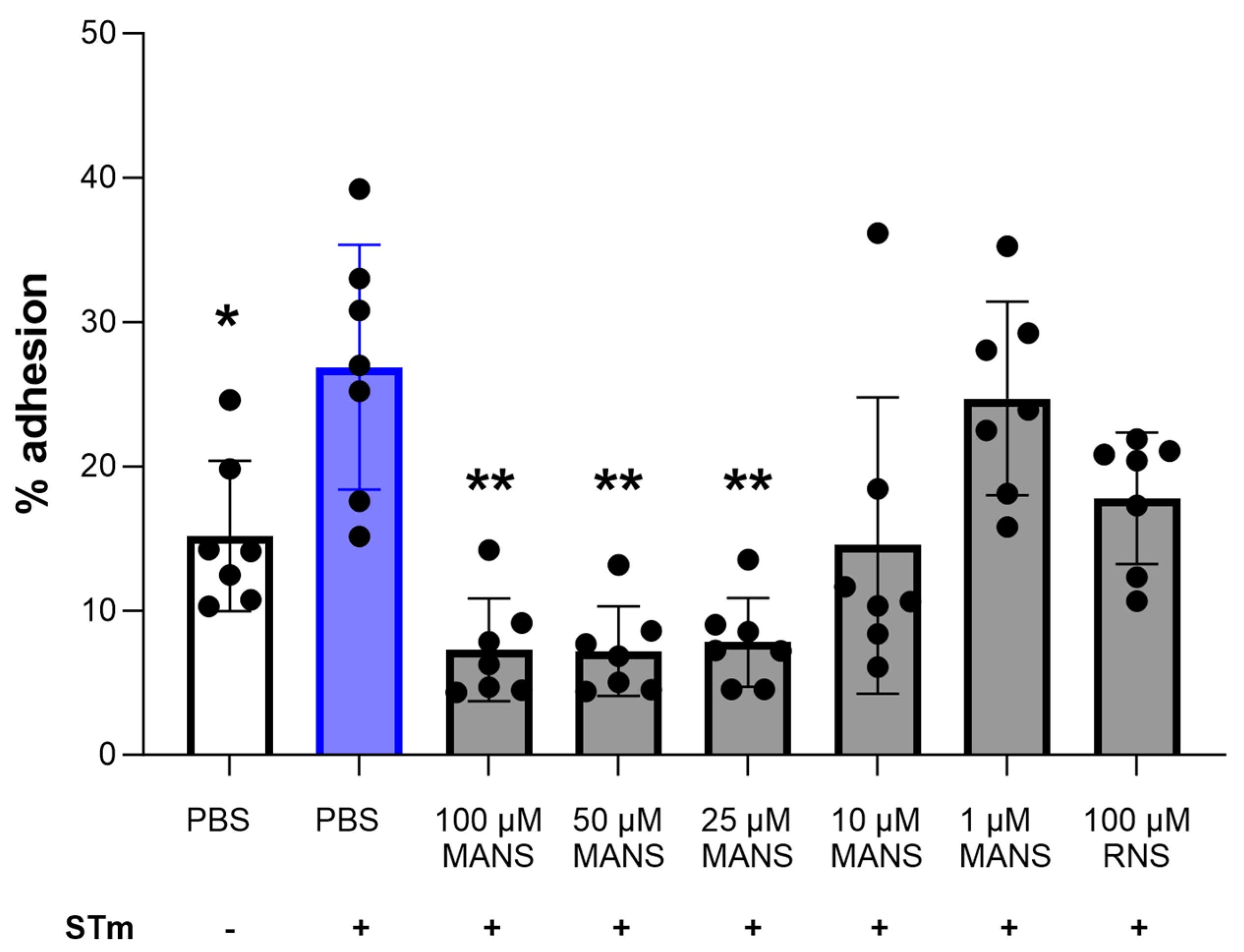
3.2. MARCKS Inhibition Attenuates STm-Induced Neutrophil Adhesion
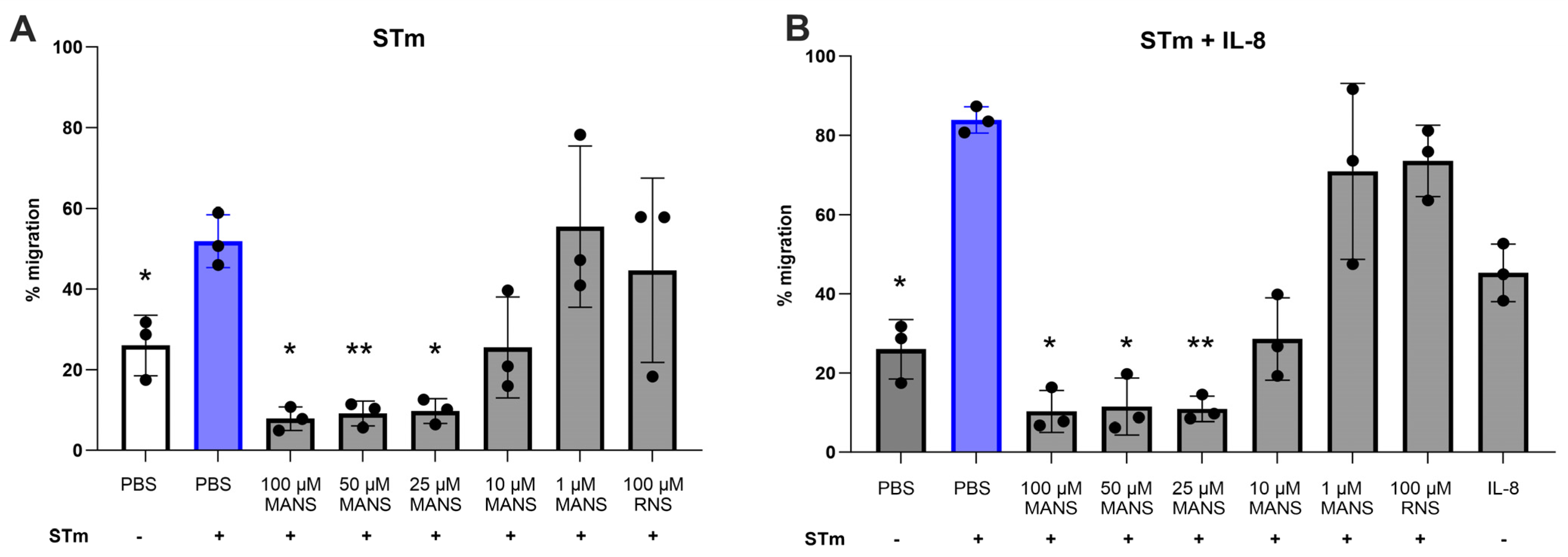
3.3. MARCKS Inhibition Attenuates STm-Induced Neutrophil Migration
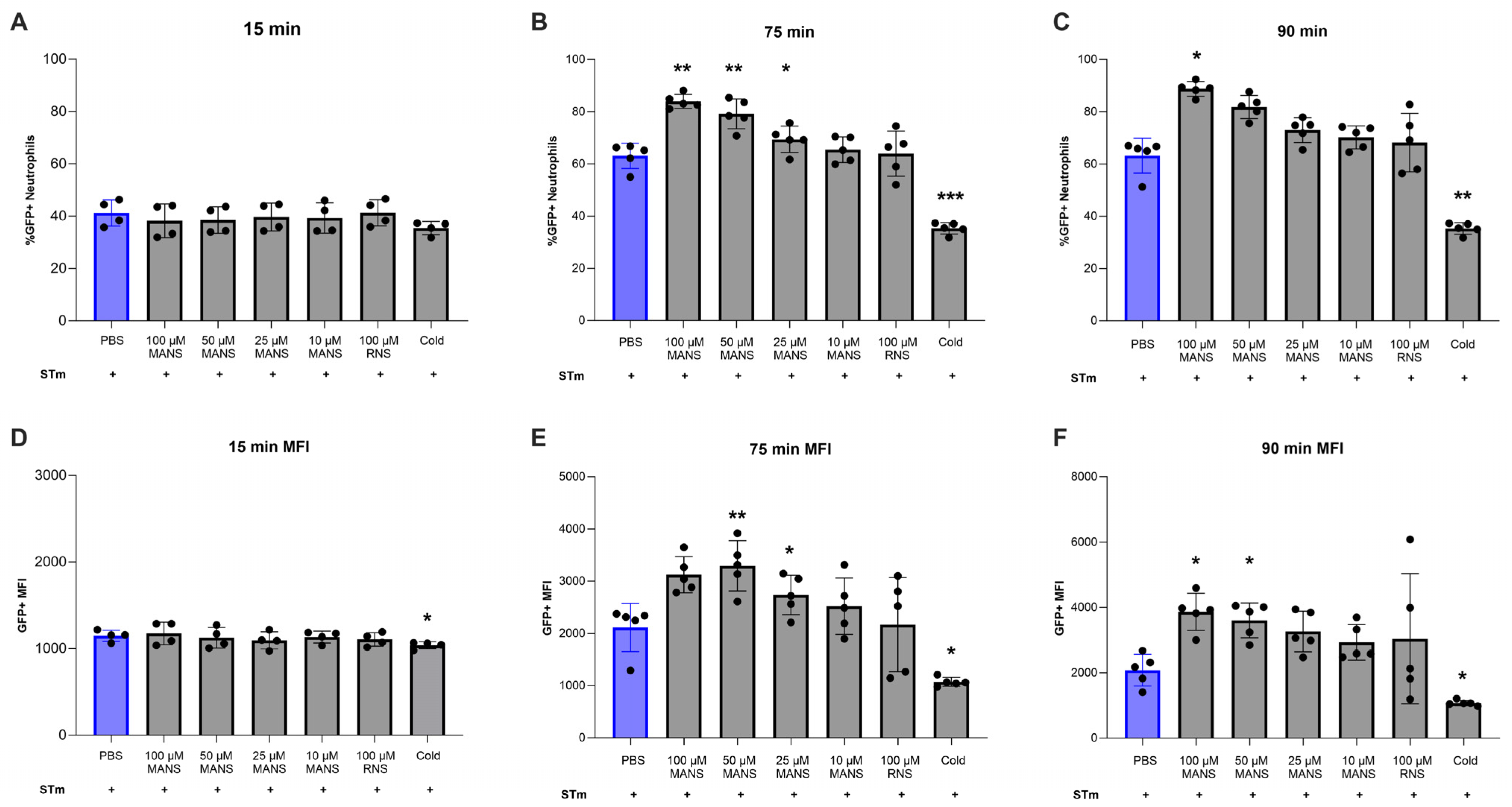
3.4. MARCKS Inhibition Enhances Neutrophil Phagocytosis of STm
3.5. MARCKS Inhibition Enhances Neutrophil Respiratory Burst in Response to STm
3.6. MARCKS Inhibition Does Not Impact Survival of STm following Co-Incubation with Neutrophils
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Döpfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 22 Foodborne Bacterial, Protozoal, and Viral Diseases, 2010: A Data Synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar] [CrossRef]
- Voetsch, A.C.; Van Gilder, T.J.; Angulo, F.J.; Farley, M.M.; Shallow, S.; Marcus, R.; Cieslak, P.R.; Deneen, V.C.; Tauxe, R.V. FoodNet estimate of the burden of illness caused by nontyphoidal Salmonella infections in the United States. Clin. Infect. Dis. 2004, 38, S127–S134. [Google Scholar] [CrossRef]
- Sell, J.; Dolan, B. Common Gastrointestinal Infections. Prim. Care Clin. Off. Pract. 2018, 45, 519–532. [Google Scholar] [CrossRef]
- Plumb, I.; Fields, P.; Bruce, B.B. Salmonellosis, Nontyphoidal. Available online: https://wwwnc.cdc.gov/travel/yellowbook/2024/infections-diseases/salmonellosis-nontyphoidal#:~:text=Nontyphoidalsalmonellaeareoneof,57%2C000deathsgloballyeachyear (accessed on 27 June 2023).
- Hoelzer, K.; Switt, A.I.M.; Wiedmann, M. Animal contact as a source of human non-typhoidal salmonellosis. Vet. Res. 2011, 42, 34. [Google Scholar] [CrossRef]
- Henao, O.L.; Jones, T.F.; Vugia, D.J.; Griffin, P.M. Foodborne diseases active surveillance network—2 decades of achievements, 1996–2015. Emerg. Infect. Dis. 2015, 21, 1529–1536. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.C.; Kulp, S.K.; Soni, S.; Wang, D.; Gunn, J.S.; Schlesinger, L.S.; Chen, C.S. Eradication of intracellular Salmonella enterica serovar typhimurium with a small-molecule, host cell-directed agent. Antimicrob. Agents Chemother. 2009, 53, 5236–5244. [Google Scholar] [CrossRef] [PubMed]
- Coburn, B.; Grassl, G.A.; Finlay, B.B. Salmonella, the host and disease: A brief review. Immunol. Cell Biol. 2007, 85, 112–118. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH oxidase activation and bacterial resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef] [PubMed]
- Langereis, J.D. Neutrophil integrin affinity regulation in adhesion, migration, and bacterial clearance. Cell Adhes. Migr. 2013, 7, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, S.; Galdiero, M.R.; Del Prete, D.; Cassatella, M.A.; Garlanda, C.; Mantovani, A. Neutrophils in innate and adaptive immunity. Semin. Immunopathol. 2013, 35, 377–394. [Google Scholar] [CrossRef] [PubMed]
- Geddes, K.; Cruz III, F.; Heffron, F.; Cruz, F.; Heffron, F. Analysis of cells targeted by Salmonella type III secretion in vivo. PLoS Pathog. 2007, 3, 2017–2028. [Google Scholar] [CrossRef]
- Stecher, B.; Robbiani, R.; Walker, A.W.; Westendorf, A.M.; Barthel, M.; Kremer, M.; Chaffron, S.; Macpherson, A.J.; Buer, J.; Parkhill, J.; et al. Salmonella enterica serovar typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biol. 2007, 5, 2177–2189. [Google Scholar] [CrossRef]
- Gallois, A.; Klein, J.R.; Allen, L.-A.H.; Jones, B.D.; Nauseef, W.M. Salmonella Pathogenicity Island 2-Encoded Type III Secretion System Mediates Exclusion of NADPH Oxidase Assembly from the Phagosomal Membrane. J. Immunol. 2001, 166, 5741–5748. [Google Scholar] [CrossRef]
- Winter, S.E.; Thiennimitr, P.; Winter, M.G.; Butler, B.P.; Huseby, D.L.; Crawford, R.W.; Russell, J.M.; Bevins, C.L.; Adams, L.G.; Tsolis, R.M.; et al. Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature 2010, 467, 426–429. [Google Scholar] [CrossRef]
- Diaz-Ochoa, V.E.; Lam, D.; Lee, C.S.; Klaus, S.; Behnsen, J.; Liu, J.Z.; Chim, N.; Nuccio, S.P.; Rathi, S.G.; Mastroianni, J.R.; et al. Salmonella Mitigates Oxidative Stress and Thrives in the Inflamed Gut by Evading Calprotectin-Mediated Manganese Sequestration. Cell Host Microbe 2016, 19, 814–825. [Google Scholar] [CrossRef] [PubMed]
- Faber, F.; Tran, L.; Byndloss, M.X.; Lopez, C.A.; Velazquez, E.M.; Kerrinnes, T.; Nuccio, S.-P.; Wangdi, T.; Fiehn, O.; Tsolis, R.M.; et al. Host-mediated sugar oxidation promotes post-antibiotic pathogen expansion. Nature 2016, 534, 697–699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Adams, L.G.; Nunes, J.; Khare, S.; Tsolis, R.M.; Bäumler, A.J. Secreted effector proteins of Salmonella enterica serotype typhimurium elicit host-specific chemokine profiles in animal models of typhoid fever and enterocolitis. Infect. Immun. 2003, 71, 4795–4803. [Google Scholar] [CrossRef] [PubMed]
- Loetscher, Y.; Wieser, A.; Lengefeld, J.; Kaiser, P.; Schubert, S.; Heikenwalder, M.; Hardt, W.D.; Stecher, B. Salmonella transiently reside in luminal neutrophils in the inflamed gut. PLoS ONE 2012, 7, e34812. [Google Scholar] [CrossRef] [PubMed]
- Spees, A.M.; Kingsbury, D.D.; Wangdi, T.; Xavier, M.N.; Tsolis, R.M.; Bäumler, A.J. Neutrophils are a source of gamma interferon during acute Salmonella enterica serovar typhimurium colitis. Infect. Immun. 2014, 82, 1692–1697. [Google Scholar] [CrossRef] [PubMed]
- Nairn, A.C.; Aderem, A. Calmodulin and protein kinase C cross-talk: The MARCKS protein is an actin filament and plasma membrane cross-linking protien regulated by protein kinase C phosphorylation and calmodulin. Ciba Found. Symp. 1992, 164, 145–154; discussion 154–161. [Google Scholar] [CrossRef] [PubMed]
- Arbuzova, A.; Schmitz, A.A.P.; Res, G.V. Cross-talk unfolded: MARCKS proteins. Biochem. J. 2002, 362, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.A.H.; Aderem, A. A role for MARCKS, the α isozyme of protein kinase c and myosin I in zymosan phagocytosis by macrophages. J. Exp. Med. 1995, 182, 829–840. [Google Scholar] [CrossRef]
- Sheats, M.K.; Sung, E.J.; Adler, K.B.; Jones, S.L. In vitro neutrophil migration requires protein kinase c-delta (δ-PKC) mediated MARCKS (Myristoylated Alanine Rich C-Kinase Substrate) phosphorylation. Inflammation 2015, 38, 1126–1141. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Arbuzova, A.; Hangyas-Mihalyne, G.; McLaughlin, S. The Effector Domain of Myristoylated Alanine-rich C Kinase Substrate Binds Strongly to Phosphatidylinositol 4,5-Bisphosphate. J. Biol. Chem. 2001, 276, 5012–5019. [Google Scholar] [CrossRef]
- Hartwig, J.H.; Thelen, M.; Rosen, A.; Janmey, P.A.; Nairn, A.C.; Aderem, A. MARCKS is an actin filament crosslinking protein regulated by protein kinase C and calcium-calmodulin. Nature 1992, 356, 618–622. [Google Scholar] [CrossRef]
- Disatnik, M.-H.M.-H.; Boutet, S.C.; Pacio, W.; Chan, A.Y.; Ross, L.B.; Lee, C.H.; Rando, T.A. The bi-directional translocation of MARCKS between membrane and cytosol regulates integrin-mediated muscle cell spreading. J. Cell Sci. 2004, 117, 4469–4479. [Google Scholar] [CrossRef]
- Sheats, M.K.; Pescosolido, K.C.; Hefner, E.M.; Sung, J.; Adler, K.B.; Jones, S.L. Myristoylated Alanine Rich C Kinase Substrate (MARCKS) is essential to β2-integrin dependent responses of equine neutrophils. Vet. Immunol. Immunopathol. 2014, 160, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; D’Annibale-Tolhurst, M.; Adler, K.; Al, E. A myristoylated alanine-rich C kinase substrate-related peptide suppresses cytokine mRNA and protein expression in LPS-activated canine neutrophils. Am. J. Respir. Cell Mol. Biol. 2013, 48, 314–321. [Google Scholar] [CrossRef]
- Conley, H.; Till, R.L.; Berglund, A.K.; Jones, S.L.; Sheats, M.K. A myristoylated alanine-rich C-kinase substrate (MARCKS) inhibitor peptide attenuates neutrophil outside-in β2-integrin activation and signaling. Cell Adhes. Migr. 2023, 17, 1–16. [Google Scholar] [CrossRef]
- Eckert, R.E.; Neuder, L.E.; Park, J.; Adler, K.B.; Jones, S.L. Myristoylated Alanine-Rich C-Kinase Substrate (MARCKS) Protein Regulation of Human Neutrophil Migration. Am. J. Respir. Cell Mol. Biol. 2010, 42, 586–594. [Google Scholar] [CrossRef]
- Wang, C.-N.; Lin, Y.-C.; Chang, B.-C.; Chen, C.-H.; Wu, R.; Lee, C.-C. Targeting the phosphorylation site of myristoylated alanine-rich C kinase substrate alleviates symptoms in a murine model of steroid-resistant asthma. Br. J. Pharmacol. 2019, 176, 1122–1134. [Google Scholar] [CrossRef]
- Damera, G.; Jester, W.F.; Jiang, M.; Zhao, H.; Fogle, H.W.; Mittelman, M.; Haczku, A.; Murphy, E.; Parikh, I.; Panettieri, R.A. Inhibition of myristoylated alanine-rich C kinase substrate (MARCKS) protein inhibits ozone-induced airway neutrophilia and inflammation. Exp. Lung Res. 2010, 36, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Adler, K.; Kraft, M.; Parikh, I.; Murphy, E.; Van As, A. Results of a phase 2a clinical trial with a peptide inhibitor of MARCKS protein indicate improvement of indices of bronchitis and lung function in patients with COPD. Eur. Respir. J. 2011, 38, 4901. [Google Scholar]
- Adler, K.B.; Parikh, I.; Crews, A.; Fang, S.; Yin, Q.; Dickson, B. Effect of an anti-MARCKS peptide (BIO-11006) on metastasis of lung cancer in vivo. J. Clin. Oncol. 2017, 35 (Suppl. 15), e23017. [Google Scholar] [CrossRef]
- Yin, Q.; Fang, S.; Park, J.; Crews, A.L.; Parikh, I.; Adler, K.B. An Inhaled Inhibitor of Myristoylated Alanine-Rich C Kinase Substrate Reverses LPS-Induced Acute Lung Injury in Mice. Am. J. Respir. Cell Mol. Biol. 2016, 55, 617–622. [Google Scholar] [CrossRef]
- Anderson, W.H.; Ceppe, A.; Rice, C.; Meyer, N.J.; Rice, T.W.; Gibbs, K.; Adler, K.B.; Carson, S.S. A Phase II, Placebo-Controlled, Multicenter Pilot Study to Evaluate the Safety and Efficacy of BIO&# 173;-11006 Inhalation Solution in Patients with Acute Respiratory Distress Syndrome. In TP52. TP052 ARDS STUDIES; American Thoracic Society: New York, NY, USA, 2021; p. 2668. [Google Scholar]
- Bogomolnaya, L.M.; Santiviago, C.A.; Yang, H.J.; Baumler, A.J.; Andrews-Polymenis, H.L. “Form variation” of the O12 antigen is critical for persistence of Salmonella Typhimurium in the murine intestine. Mol. Microbiol. 2008, 70, 1105–1119. [Google Scholar] [CrossRef]
- Westerman, T.L.; Bogomolnaya, L.; Andrews-Polymenis, H.L.; Sheats, M.K.; Elfenbein, J.R. The Salmonella type-3 secretion system-1 and flagellar motility influence the neutrophil respiratory burst. PLoS ONE 2018, 13, e0203698. [Google Scholar] [CrossRef]
- Westerman, T.L.; Sheats, M.K.; Elfenbein, J.R. Sulfate import in salmonella typhimurium impacts bacterial aggregation and the respiratory burst in human neutrophils. Infect. Immun. 2021, 89, e00701-20. [Google Scholar] [CrossRef]
- Staali, L.; Mörgelin, M.; Björck, L.; Tapper, H. Streptococcus pyogenes expressing M and M-like surface proteins are phagocytosed but survive inside human neutrophils. Cell. Microbiol. 2003, 5, 253–265. [Google Scholar] [CrossRef]
- Hartmann, P.; Becker, R.; Franzen, C.; Schell-Frederick, E.; Römer, J.; Jacobs, M.; Fätkenheuer, G.; Plum, G. Phagocytosis and killing of Mycobacterium avium complex by human neutrophils. J. Leukoc. Biol. 2001, 69, 397–404. [Google Scholar] [CrossRef]
- Jin, Q.; Jiang, L.; Chen, Q.; Li, X.; Xu, Y.; Sun, X.; Zhao, Z.; Wei, L. Rapid flow cytometry-based assay for the evaluation of γδ T cell-mediated cytotoxicity. Mol. Med. Rep. 2018, 17, 3555–3562. [Google Scholar] [CrossRef] [PubMed]
- Boero, E.; Brinkman, I.; Juliet, T.; van Yperen, E.; van Strijp, J.A.G.; Rooijakkers, S.H.M.; van Kessel, K.P.M. Use of Flow Cytometry to Evaluate Phagocytosis of Staphylococcus aureus by Human Neutrophils. Front. Immunol. 2021, 12, 635825. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Thai, P.; Yoneda, K.; Adler, K.B.; Yang, P.-C.; Wu, R. A peptide that inhibits function of Myristoylated Alanine-Rich C Kinase Substrate (MARCKS) reduces lung cancer metastasis. Oncogene 2014, 33, 3696–3706. [Google Scholar] [CrossRef]
- Li, Y.; Martin, L.D.; Spizz, G.; Adler, K.B. MARCKS Protein Is a Key Molecule Regulating Mucin Secretion by Human Airway Epithelial Cells in Vitro. J. Biol. Chem. 2001, 276, 40982–40990. [Google Scholar] [CrossRef] [PubMed]
- Rohrbach, T.D.; Jones, R.B.; Hicks, P.H.; Weaver, A.N.; Cooper, T.S.; Eustace, N.J.; Yang, E.S.; Jarboe, J.S.; Anderson, J.C.; Willey, C.D. MARCKS phosphorylation is modulated by a peptide mimetic of MARCKS effector domain leading to increased radiation sensitivity in lung cancer cell lines. Oncol. Lett. 2017, 13, 1216–1222. [Google Scholar] [CrossRef]
- Manček-Keber, M.; Benčina, M.; Japelj, B.; Panter, G.; Andrä, J.; Brandenburg, K.; Triantafilou, M.; Triantafilou, K.; Jerala, R. MARCKS as a Negative Regulator of Lipopolysaccharide Signaling. J. Immunol. 2012, 188, 3893–3902. [Google Scholar] [CrossRef]
- Rohrbach, T.D.; Shah, N.; Jackson, W.P.; Feeney, E.V.; Scanlon, S.; Gish, R.; Khodadadi, R.; Hyde, S.O.; Hicks, P.H.; Anderson, J.C.; et al. The effector domain of MARCKS Is a nuclear localization signal that regulates cellular PIP2 levels and nuclear PIP2 localization. PLoS ONE 2015, 10, e0140870. [Google Scholar] [CrossRef]
- Schmidt, S.; Moser, M.; Sperandio, M. The molecular basis of leukocyte recruitment and its deficiencies. Mol. Immunol. 2013, 55, 49–58. [Google Scholar] [CrossRef]
- Fournier, B.M.; Parkos, C.A. The role of neutrophils during intestinal inflammation. Mucosal Immunol. 2012, 5, 354–366. [Google Scholar] [CrossRef]
- Adusumilli, S.; Mve-Obiang, A.; Sparer, T.; Meyers, W.; Hayman, J.; Small, P.L.C. Mycobacterium ulcerans toxic macrolide, mycolactone modulates the host immune response and cellular location of M. ulcerans in vitro and in vivo. Cell. Microbiol. 2005, 7, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- McHugh, B.J.; Wang, R.; Li, H.N.; Beaumont, P.E.; Kells, R.; Stevens, H.; Young, L.; Rossi, A.G.; Gray, R.D.; Dorin, J.R.; et al. Cathelicidin is a “fire alarm”, generating protective NLRP3-dependent airway epithelial cell inflammatory responses during infection with Pseudomonas aeruginosa. PLoS Pathog. 2019, 15, e1007694. [Google Scholar] [CrossRef] [PubMed]
- van Bruggen, R.; Zweers, D.; Van Diepen, A.; Van Dissel, J.T.; Roos, D.; Verhoeven, A.J.; Kuijpers, T.W. Complement Receptor 3 and Toll-Like Receptor 4 Act Sequentially in Uptake and Intracellular Killing of Unopsonized Salmonella enterica Serovar Typhimurium by Human Neutrophils ᰔ. Infect. Immun. 2007, 75, 2655–2660. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Arai, T. A possible mechanism for host-specific pathogenesis of Salmonella serovars. Microb. Pathog. 1996, 21, 435–446. [Google Scholar] [CrossRef]
- Grossman, N.; Joiner, K.; Frank, M.; Leive, L. C3b binding, but not its breakdown, is affected by the structure of the O-antigen polysaccharide in lipopolysaccharide from Salmonellae. J. Immunol. 1986, 136, 2208–2215. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, L.; Huber-Lang, M.; Weiss, M.E.; Hohmann, H.; Schmolz, M.; Schneider, E.M. Phagocytosis and digestion of pH-sensitive fluorescent dye (Eos-FP) transfected E. coli in whole blood assays from patients with severe sepsis and septic shock. J. Cell Commun. Signal. 2011, 5, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Suk, K.; Lee, W.H. Myristoylated alanine-rich C kinase substrate (MARCKS) regulates the expression of proinflammatory cytokines in macrophages through activation of p38/JNK MAPK and NF-κB. Cell Immunol. 2015, 296, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Elzagallaai, A.; Rosé, S.D.; Trifaró, J.M. Platelet secretion induced by phorbol esters stimulation is mediated through phosphorylation of MARCKS: A MARCKS-derived peptide blocks MARCKS phosphorylation and serotonin release without affecting pleckstrin phosphorylation. Blood 2000, 95, 894–902. [Google Scholar] [CrossRef]
- Chen, C.H.; Statt, S.; Chiu, C.L.; Thai, P.; Arif, M.; Adler, K.B.; Wu, R. Targeting myristoylated alanine-rich C kinase substrate phosphorylation site domain in lung cancer: Mechanisms and therapeutic implications. Am. J. Respir. Crit. Care Med. 2014, 190, 1127–1138. [Google Scholar] [CrossRef]
- Eustace, N.J.; Anderson, J.C.; Warram, J.M.; Widden, H.N.; Pedersen, R.T.; Alrefai, H.; Patel, Z.; Hicks, P.H.; Placzek, W.J.; Gillespie, G.Y.; et al. A cell-penetrating MARCKS mimetic selectively triggers cytolytic death in glioblastoma. Oncogene 2020, 39, 6961–6974. [Google Scholar] [CrossRef]
- Tamura, R. 1. Study on the effector domain of MARCKS protein as a cell penetrating peptide. 2. Development of photoactivatable prodrugs of doxazolidine. Ph.D. Thesis, University of Colorado, Boulder, CO, USA, 2017. [Google Scholar]
- Darveau, R.P.; Cunningham, M.D.; Bailey, T.; Seachord, C.; Ratcliffe, K.; Bainbridge, B.; Dietsch, M.; Page, R.C.; Aruffo, A. Ability of Bacteria Associated with Chronic Inflammatory Disease To Stimulate E-Selectin Expression and Promote Neutrophil Adhesion. Infect. Immun. 1995, 63, 1311–1317. [Google Scholar] [CrossRef]
- Drevets, D.A. Listeria monocytogenes infection of cultured endothelial cells stimulates neutrophil adhesion and adhesion molecule expression. J. Immunol. 1997, 158, 5305–5313. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.A.; Silva, M.; Siber, A.M.; Kelly, A.J.; Galyov, E.; McCormick, B.A. A secreted Salmonella protein induces a proinflammatory response in epithelial cells, which promotes neutrophil migration. Proc. Natl. Acad. Sci. USA 2000, 97, 12283–12288. [Google Scholar] [CrossRef] [PubMed]
- Hiyoshi, H.; Wangdi, T.; Lock, G.; Saechao, C.; Raffatellu, M.; Cobb, B.A.; Bäumler, A.J. Mechanisms to Evade the Phagocyte Respiratory Burst Arose by Convergent Evolution in Typhoidal Salmonella Serovars. Cell Rep. 2018, 22, 1787–1797. [Google Scholar] [CrossRef]
- Carballo, E.; Pitterle, D.M.; Stumpo, D.J.; Sperling, R.T.; Blackshear, P.J. Phagocytic and macropinocytic activity in MARCKS-deficient macrophages and fibroblasts. Am. J. Physiol. 1999, 277, C163–C173. [Google Scholar] [CrossRef]
- Gresham, H.D.; Dale, B.M.; Potter, J.W.; Chang, P.W.; Vines, C.M.; Lowell, C.A.; Lagenaur, C.F.; Willman, C.L. Negative regulation of phagocytosis in murine macrophages by the Src kinase family member, Fgr. J. Exp. Med. 2000, 191, 515–528. [Google Scholar] [CrossRef]
- Cox, D.; Dale, B.M.; Kashiwada, M.; Helgason, C.D.; Greenberg, S. A regulatory role for Src homology 2 domain-containing inositol 5′-phosphatase (SHIP) in phagocytosis mediated by Fcγ receptors and complement receptor 3 (αMβ2; CD11b/CD18). J. Exp. Med. 2001, 193, 61–71. [Google Scholar] [CrossRef] [PubMed]
- May, R.C.; Machesky, L.M. Phagocytosis and the actin cytoskeleton. J. Cell Sci. 2001, 114, 1061–1077. [Google Scholar] [CrossRef]
- Zaldívar-López, S.; Herrera-Uribe, J.; Bautista, R.; Jiménez, Á.; Moreno, Á.; Claros, M.G.; Garrido, J.J. Salmonella Typhimurium induces genome-wide expression and phosphorylation changes that modulate immune response, intracellular survival and vesicle transport in infected neutrophils. Dev. Comp. Immunol. 2023, 140, 104597. [Google Scholar] [CrossRef]
- Blander, J.M.; Medzhitov, R. Regulation of Phagosome Maturation by Signals from Toll-Like Receptors. Science 2004, 304, 1014–1018. [Google Scholar] [CrossRef]
- Arpaia, N.; Godec, J.; Lau, L.; Sivick, K.E.; McLaughlin, L.M.; Jones, M.B.; Dracheva, T.; Peterson, S.N.; Monack, D.M.; Barton, G.M. TLR signaling is required for Salmonella typhimurium virulence. Cell 2011, 144, 675–688. [Google Scholar] [CrossRef]
- Feuillet, V.; Medjane, S.; Mondor, I.; Demaria, O.; Pagni, P.P.; Galán, J.E.; Flavell, R.A.; Alexopoulou, L. Involvement of toll-like receptor 5 in the recognition of flagellated bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 12487–12492. [Google Scholar] [CrossRef] [PubMed]
- Torres-Gomez, A.; Luis Sanchez-Trincado, J.; Toribio, V.; Torres-Ruiz, R.; Rodríguez-Perales, S.; Yáñez-Mó, M.; Reche, P.A.; Cabañas, C.; Lafuente, E.M. RIAM-VASP Module Relays Integrin Complement Receptors in Outside-In Signaling Driving Particle Engulfment. Cells 2020, 9, 1166. [Google Scholar] [CrossRef] [PubMed]
- Conley, H.E. A Comparative Investigation of MARCKS as a Therapeutic Target for Dysregulated Neutrophils. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 2022. [Google Scholar]
- Schmidt, A.; Caron, E.; Hall, A. Lipopolysaccharide-Induced Activation of β2-Integrin Function in Macrophages Requires Irak Kinase Activity, p38 Mitogen- Activated Protein Kinase, and the Rap1 GTPase. Mol. Cell. Biol. 2001, 21, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.A.H.; DeLeo, F.R.; Gallois, A.; Toyoshima, S.; Suzuki, K.; Nauseef, W.M. Transient association of the nicotinamide adenine dinucleotide phosphate oxidase subunits p47phox and p67phox with phagosomes in neutrophils from patients with X-linked chronic granulomatous disease. Blood 1999, 93, 3521–3530. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, F.R.; Allen, L.A.; Apicella, M.; Nauseef, W.M. NADPH oxidase activation and assembly during phagocytosis. J. Immunol. 1999, 163, 6732–6740. [Google Scholar] [CrossRef]
- Karimi, G.; Houée Levin, C.; Dagher, M.C.; Baciou, L.; Bizouarn, T. Assembly of phagocyte NADPH oxidase: A concerted binding process? Biochim. Biophys. Acta—Gen. Subj. 2014, 1840, 3277–3283. [Google Scholar] [CrossRef]
- Tuinema, C.R.; Sa, R.-Y.; Coombes, B.K. Salmonella evades D-amino acid oxidase to promote infection in neutrophils. MBio 2014, 5, e01886-14. [Google Scholar] [CrossRef]
- Rhen, M. Salmonella and Reactive Oxygen Species: A Love-Hate Relationship. J. Innate Immun. 2019, 11, 216–226. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conley, H.E.; Brown, C.F.; Westerman, T.L.; Elfenbein, J.R.; Sheats, M.K. MARCKS Inhibition Alters Bovine Neutrophil Responses to Salmonella Typhimurium. Biomedicines 2024, 12, 442. https://doi.org/10.3390/biomedicines12020442
Conley HE, Brown CF, Westerman TL, Elfenbein JR, Sheats MK. MARCKS Inhibition Alters Bovine Neutrophil Responses to Salmonella Typhimurium. Biomedicines. 2024; 12(2):442. https://doi.org/10.3390/biomedicines12020442
Chicago/Turabian StyleConley, Haleigh E., Chalise F. Brown, Trina L. Westerman, Johanna R. Elfenbein, and M. Katie Sheats. 2024. "MARCKS Inhibition Alters Bovine Neutrophil Responses to Salmonella Typhimurium" Biomedicines 12, no. 2: 442. https://doi.org/10.3390/biomedicines12020442