
Breaking Barriers: The Promise and Challenges of Immune Checkpoint Inhibitors in Triple-Negative Breast Cancer


Abstract
:1. Introduction
2. Rationale Shift from Chemotherapy to Immunotherapy
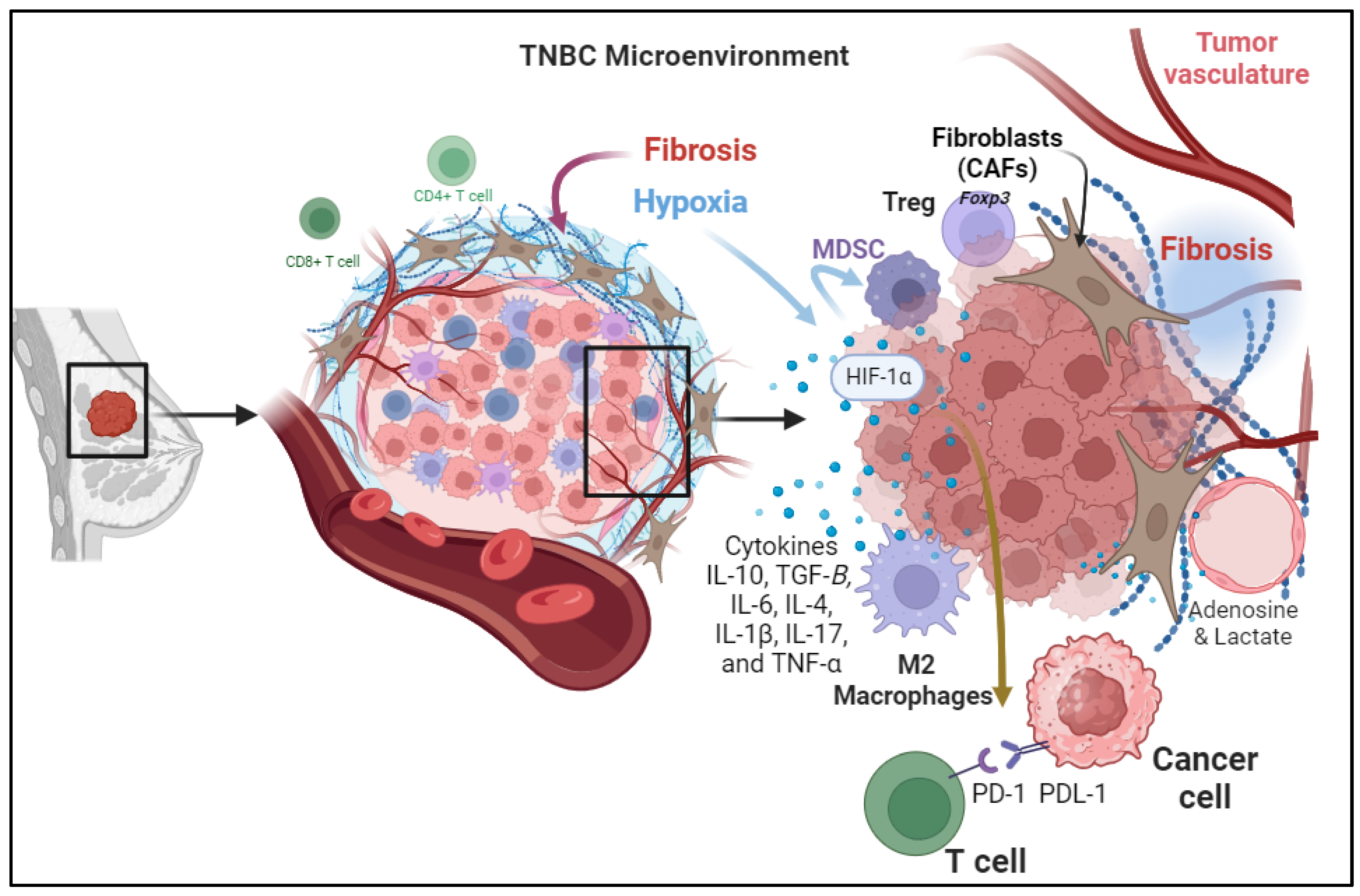
3. Immunosuppressive Mechanisms within TME of TNBC Patient
3.1. Cellular Components of the TME
3.2. Tumor-Induced Hypoxia
3.3. Secretion of Immunosuppressive Cytokines
3.4. Metabolic Reprogramming
3.5. Changes in the Configuration of the Extracellular Matrix in the TME in TNBC
4. Overcoming Mechanisms of Immune Resistance within TME of TNBC Patients
4.1. Monotherapy-ICIs
4.2. Dual ICIs
4.3. Chemotherapy Combined ICIs
4.4. Cancer Vaccine Combined ICIs
4.5. Combining ICIs with TME Metabolites
4.6. Cytokines–IFN/TGF Beta Crosstalk with ICIs
4.7. Anti CD25/Anti CD47 Combined with ICIs
4.8. Chemokine Inhibitors Cross-Talk with ICIs
4.9. Gut Microbiota Crosstalk with ICIs
5. Ongoing Challenges of Immunotherapy in TNBC Patients
Author Contributions
Funding
Conflicts of Interest
References
- Liu, Y.; Hu, Y.; Xue, J.; Li, J.; Yi, J.; Bu, J.; Zhang, Z.; Qiu, P.; Gu, X. Advances in immunotherapy for triple-negative breast cancer. Mol. Cancer 2023, 22, 145. [Google Scholar] [CrossRef]
- Eliyatkın, N.; Yalçın, E.; Zengel, B.; Aktaş, S.; Vardar, E. Molecular Classification of Breast Carcinoma: From Traditional, Old-Fashioned Way to A New Age, and A New Way. J. Breast Health 2015, 11, 59–66. [Google Scholar] [CrossRef]
- Zhang, X. Molecular Classification of Breast Cancer: Relevance and Challenges. Arch. Pathol. Lab. Med. 2023, 147, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Dixon-Douglas, J.; Loi, S. Immunotherapy in Early-Stage Triple-Negative Breast Cancer: Where Are We Now and Where Are We Headed? Curr. Treat. Options Oncol. 2023, 24, 1004–1020. [Google Scholar] [CrossRef]
- Fan, Y.; He, S. The Characteristics of Tumor Microenvironment in Triple Negative Breast Cancer. Cancer Manag. Res. 2022, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Vikas, P.; Korde, L.A.; Somerfield, M.R.; Hershman, D.L. Use of Immune Checkpoint Inhibitors in the Treatment of High-Risk, Early-Stage Triple-Negative Breast Cancer: ASCO Guideline Rapid Recommendation Update Q and A. JCO Oncol. Pract. 2022, 18, 649–651. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Al-Khadairi, G.; Decock, J. Immune Checkpoint Inhibitors in Triple Negative Breast Cancer Treatment: Promising Future Prospects. Front. Oncol. 2020, 10, 600573. [Google Scholar] [CrossRef]
- Barroso-Sousa, R.; Pacífico, J.P.; Sammons, S.; Tolaney, S.M. Tumor Mutational Burden in Breast Cancer: Current Evidence, Challenges, and Opportunities. Cancers 2023, 15, 3997. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Yi, Z.; Wang, L.; Li, Z.; Niu, B.; Ren, G. The co-expression characteristics of LAG3 and PD-1 on the T cells of patients with breast cancer reveal a new therapeutic strategy. Int. Immunopharmacol. 2020, 78, 106113. [Google Scholar] [CrossRef]
- Dongre, A.; Rashidian, M.; Reinhardt, F.; Bagnato, A.; Keckesova, Z.; Ploegh, H.L.; Weinberg, R.A. Epithelial-to-Mesenchymal Transition Contributes to Immunosuppression in Breast Carcinomas. Cancer Res. 2017, 77, 3982–3989. [Google Scholar] [CrossRef]
- Said, S.S.; Ibrahim, W.N. Cancer Resistance to Immunotherapy: Comprehensive Insights with Future Perspectives. Pharmaceutics 2023, 15, 1143. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, S.; Tang, H.; Meng, X.; Zheng, Q. Molecular mechanisms of immunotherapy resistance in triple-negative breast cancer. Front. Immunol. 2023, 14, 1153990. [Google Scholar] [CrossRef]
- Savas, P.; Salgado, R.; Denkert, C.; Sotiriou, C.; Darcy, P.K.; Smyth, M.J.; Loi, S. Clinical relevance of host immunity in breast cancer: From TILs to the clinic. Nat. Rev. Clin. Oncol. 2016, 13, 228–241. [Google Scholar] [CrossRef]
- Bense, R.D.; Sotiriou, C.; Piccart-Gebhart, M.J.; Haanen, J.; van Vugt, M.; de Vries, E.G.E.; Schröder, C.P.; Fehrmann, R.S.N. Relevance of Tumor-Infiltrating Immune Cell Composition and Functionality for Disease Outcome in Breast Cancer. J. Natl. Cancer Inst. 2017, 109, djw192. [Google Scholar] [CrossRef]
- Zheng, H.; Siddharth, S.; Parida, S.; Wu, X.; Sharma, D. Tumor Microenvironment: Key Players in Triple Negative Breast Cancer Immunomodulation. Cancers 2021, 13, 3357. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Wilkes, D.W.; Samuel, N.; Blanco, M.A.; Nayak, A.; Alicea-Torres, K.; Gluck, C.; Sinha, S.; Gabrilovich, D.; Chakrabarti, R. ΔNp63-driven recruitment of myeloid-derived suppressor cells promotes metastasis in triple-negative breast cancer. J. Clin. Investig. 2018, 128, 5095–5109. [Google Scholar] [CrossRef]
- Fu, L.Q.; Du, W.L.; Cai, M.H.; Yao, J.Y.; Zhao, Y.Y.; Mou, X.Z. The roles of tumor-associated macrophages in tumor angiogenesis and metastasis. Cell. Immunol. 2020, 353, 104119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Wang, X.H.; Gao, S.T.; Chen, C.; Xu, X.Y.; Sun, Q.; Zhou, Z.H.; Wu, G.Z.; Yu, Q.; Xu, G.; et al. Tumor-associated macrophages correlate with phenomenon of epithelial-mesenchymal transition and contribute to poor prognosis in triple-negative breast cancer patients. J. Surg. Res. 2018, 222, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, N.; Usmani, S.S.; Subbarayan, R.; Saini, R.; Pandey, P.K. Hypoxia: Syndicating triple negative breast cancer against various therapeutic regimens. Front. Oncol. 2023, 13, 1199105. [Google Scholar] [CrossRef]
- de Heer, E.C.; Jalving, M.; Harris, A.L. HIFs, angiogenesis, and metabolism: Elusive enemies in breast cancer. J. Clin. Investig. 2020, 130, 5074–5087. [Google Scholar] [CrossRef]
- Cao, Q.; Mushajiang, M.; Tang, C.Q.; Ai, X.Q. Role of hypoxia-inducible factor-1α and survivin in breast cancer recurrence and prognosis. Heliyon 2023, 9, e14132. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Gilkes, D.M.; Takano, N.; Xiang, L.; Luo, W.; Bishop, C.J.; Chaturvedi, P.; Green, J.J.; Semenza, G.L. Hypoxia-inducible factors and RAB22A mediate formation of microvesicles that stimulate breast cancer invasion and metastasis. Proc. Natl. Acad. Sci. USA 2014, 111, E3234–E3242. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, L.; He, X.; Zhang, P.; Sun, C.; Xu, X.; Lu, Y.; Li, F. TGF-β plays a vital role in triple-negative breast cancer (TNBC) drug-resistance through regulating stemness, EMT and apoptosis. Biochem. Biophys. Res. Commun. 2018, 502, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, Y.Y.; Chen, Y.; Wang, J.; Wang, Q.; Lu, H. TGF-β Signaling and Resistance to Cancer Therapy. Front. Cell Dev. Biol. 2021, 9, 786728. [Google Scholar] [CrossRef] [PubMed]
- Chakravarthy, A.; Khan, L.; Bensler, N.P.; Bose, P.; De Carvalho, D.D. TGF-β-associated extracellular matrix genes link cancer-associated fibroblasts to immune evasion and immunotherapy failure. Nat. Commun. 2018, 9, 4692. [Google Scholar] [CrossRef]
- Chang, C.M.; Lam, H.Y.P.; Hsu, H.J.; Jiang, S.J. Interleukin-10: A double-edged sword in breast cancer. Tzu Chi Med. J. 2021, 33, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Cao, L. The role of tumor-infiltrating lymphocytes in triple-negative breast cancer and the research progress of adoptive cell therapy. Front. Immunol. 2023, 14, 1194020. [Google Scholar] [CrossRef]
- Cha, Y.J.; Koo, J.S. Role of Tumor-Associated Myeloid Cells in Breast Cancer. Cells 2020, 9, 1785. [Google Scholar] [CrossRef]
- Little, A.C.; Pathanjeli, P.; Wu, Z.; Bao, L.; Goo, L.E.; Yates, J.A.; Oliver, C.R.; Soellner, M.B.; Merajver, S.D. IL-4/IL-13 Stimulated Macrophages Enhance Breast Cancer Invasion Via Rho-GTPase Regulation of Synergistic VEGF/CCL-18 Signaling. Front. Oncol. 2019, 9, 456. [Google Scholar] [CrossRef]
- Jeon, M.; Han, J.; Nam, S.J.; Lee, J.E.; Kim, S. Elevated IL-1β expression induces invasiveness of triple negative breast cancer cells and is suppressed by zerumbone. Chem. Biol. Interact. 2016, 258, 126–133. [Google Scholar] [CrossRef]
- Popović, M.; Dedić Plavetić, N.; Vrbanec, D.; Marušić, Z.; Mijatović, D.; Kulić, A. Interleukin 17 in early invasive breast cancer. Front. Oncol. 2023, 13, 1171254. [Google Scholar] [CrossRef]
- Welte, T.; Zhang, X.H. Interleukin-17 Could Promote Breast Cancer Progression at Several Stages of the Disease. Mediat. Inflamm. 2015, 2015, 804347. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, Z.; Zhou, Y.; Yang, Y. MDSCs in breast cancer: An important enabler of tumor progression and an emerging therapeutic target. Front. Immunol. 2023, 14, 1199273. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Giatromanolaki, A.; Gkegka, A.G.; Pouliliou, S.; Biziota, E.; Kakolyris, S.; Koukourakis, M. Hypoxia and anaerobic metabolism relate with immunologically cold breast cancer and poor prognosis. Breast Cancer Res. Treat. 2022, 194, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chu, Z.; Liu, M.; Zou, Q.; Li, J.; Liu, Q.; Wang, Y.; Wang, T.; Xiang, J.; Wang, B. Amino acid metabolism in immune cells: Essential regulators of the effector functions, and promising opportunities to enhance cancer immunotherapy. J. Hematol. Oncol. 2023, 16, 59. [Google Scholar] [CrossRef]
- Hasmim, M.; Xiao, M.; Van Moer, K.; Kumar, A.; Oniga, A.; Mittelbronn, M.; Duhem, C.; Chammout, A.; Berchem, G.; Thiery, J.P.; et al. SNAI1-dependent upregulation of CD73 increases extracellular adenosine release to mediate immune suppression in TNBC. Front. Immunol. 2022, 13, 982821. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Cheng, Z.; He, J.; Yao, X.; Liu, Y.; Cai, K.; Li, M.; Hu, Y.; Luo, Z. Inhibition of glycolysis-driven immunosuppression with a nano-assembly enhances response to immune checkpoint blockade therapy in triple negative breast cancer. Nat. Commun. 2023, 14, 7021. [Google Scholar] [CrossRef]
- Ding, J.H.; Xiao, Y.; Zhao, S.; Xu, Y.; Xiao, Y.L.; Shao, Z.M.; Jiang, Y.Z.; Di, G.H. Integrated analysis reveals the molecular features of fibrosis in triple-negative breast cancer. Mol. Ther. Oncolytics 2022, 24, 624–635. [Google Scholar] [CrossRef]
- Sheng, G.; Yuan, H.; Jin, L.; Ranjit, S.; Panov, J.; Lu, X.; Levi, M.; Glazer, R.I. Reduction of fibrosis and immune suppressive cells in ErbB2-dependent tumorigenesis by an LXR agonist. PLoS ONE 2021, 16, e0248996. [Google Scholar] [CrossRef]
- Huang, P.; Zhou, X.; Zheng, M.; Yu, Y.; Jin, G.; Zhang, S. Regulatory T cells are associated with the tumor immune microenvironment and immunotherapy response in triple-negative breast cancer. Front. Immunol. 2023, 14, 1263537. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Jiang, J.; Yao, W.; Hu, Y.; Kong, S.; Fan, Q.; Yan, X.; Li, F.; Shi, Z. Pirfenidone inhibits stromal collagen deposition and improves intra-tumoral delivery and antitumor efficacy of Pegylated liposomal doxorubicin. Biomed. Pharmacother. 2023, 157, 114015. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.; Carvalho, M.J.; Goncalves, J.; Moreira, J.N. Immunotherapy in triple-negative breast cancer: Insights into tumor immune landscape and therapeutic opportunities. Front. Mol. Biosci. 2022, 9, 903065. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, J.; Hu, T.; Wang, H.; Long, M.; Liang, B. Adverse Events of PD-1 or PD-L1 Inhibitors in Triple-Negative Breast Cancer: A Systematic Review and Meta-Analysis. Life 2022, 12, 1990. [Google Scholar] [CrossRef]
- Zhu, K.; Chen, H.; Xu, C.; Chen, D.; Jin, Z.; Ren, S.; Witharana, P.; Chen, B.; Shen, J. Efficacy and safety of immune checkpoint inhibitors versus chemotherapy in the second-line treatment of advanced esophageal squamous cell carcinoma: A meta-analysis and systematic review. J. Thorac. Dis. 2023, 15, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Dirix, L.Y.; Takacs, I.; Jerusalem, G.; Nikolinakos, P.; Arkenau, H.T.; Forero-Torres, A.; Boccia, R.; Lippman, M.E.; Somer, R.; Smakal, M.; et al. Avelumab, an anti-PD-L1 antibody, in patients with locally advanced or metastatic breast cancer: A phase 1b JAVELIN Solid Tumor study. Breast Cancer Res. Treat. 2018, 167, 671–686. [Google Scholar] [CrossRef]
- Lotfinejad, P.; Kazemi, T.; Safaei, S.; Amini, M.; Roshani Asl, E.; Baghbani, E.; Sandoghchian Shotorbani, S.; Jadidi Niaragh, F.; Derakhshani, A.; Abdoli Shadbad, M.; et al. PD-L1 silencing inhibits triple-negative breast cancer development and upregulates T-cell-induced pro-inflammatory cytokines. Biomed. Pharmacother. 2021, 138, 111436. [Google Scholar] [CrossRef] [PubMed]
- Alkaabi, D.; Arafat, K.; Sulaiman, S.; Al-Azawi, A.M.; Attoub, S. PD-1 Independent Role of PD-L1 in Triple-Negative Breast Cancer Progression. Int. J. Mol. Sci. 2023, 24, 6420. [Google Scholar] [CrossRef] [PubMed]
- Santa-Maria, C.A.; Kato, T.; Park, J.H.; Kiyotani, K.; Rademaker, A.; Shah, A.N.; Gross, L.; Blanco, L.Z.; Jain, S.; Flaum, L.; et al. A pilot study of durvalumab and tremelimumab and immunogenomic dynamics in metastatic breast cancer. Oncotarget 2018, 9, 18985–18996. [Google Scholar] [CrossRef]
- Egelston, C.A.; Guo, W.; Yost, S.E.; Ge, X.; Lee, J.S.; Frankel, P.H.; Cui, Y.; Ruel, C.; Schmolze, D.; Murga, M.; et al. Immunogenicity and efficacy of pembrolizumab and doxorubicin in a phase I trial for patients with metastatic triple-negative breast cancer. Cancer Immunol. Immunother. 2023, 72, 3013–3027. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Crosby, E.J.; Wei, J.; Yang, X.Y.; Lei, G.; Wang, T.; Liu, C.X.; Agarwal, P.; Korman, A.J.; Morse, M.A.; Gouin, K.; et al. Complimentary mechanisms of dual checkpoint blockade expand unique T-cell repertoires and activate adaptive anti-tumor immunity in triple-negative breast tumors. Oncoimmunology 2018, 7, e1421891. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, J.; Cho, E.Y.; Park, Y.H.; Ahn, J.S.; Kim, K.M.; Im, Y.H. Lymphocyte-activating gene 3 expression in tumor cells predicts immune checkpoint inhibitor response in triple negative breast cancer. Front. Oncol. 2023, 13, 1146934. [Google Scholar] [CrossRef] [PubMed]
- Tagliamento, M.; Agostinetto, E.; Borea, R.; Brandão, M.; Poggio, F.; Addeo, A.; Lambertini, M. VISTA: A Promising Target for Cancer Immunotherapy? Immunotargets Ther. 2021, 10, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Pilones, K.A.; Hensler, M.; Daviaud, C.; Kraynak, J.; Fucikova, J.; Galluzzi, L.; Demaria, S.; Formenti, S.C. Converging focal radiation and immunotherapy in a preclinical model of triple negative breast cancer: Contribution of VISTA blockade. Oncoimmunology 2020, 9, 1830524. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Gao, S.; Gou, J.; Yin, T.; He, H.; Wang, Y.; Zhang, Y.; Tang, X.; Wu, R. Platinum-based chemotherapy in combination with PD-1/PD-L1 inhibitors: Preclinical and clinical studies and mechanism of action. Expert Opin. Drug Deliv. 2021, 18, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Merkher, Y.; Chen, L.; Liu, N.; Leonov, S.; Chen, Y. Recent advances in therapeutic strategies for triple-negative breast cancer. J. Hematol. Oncol. 2022, 15, 121. [Google Scholar] [CrossRef] [PubMed]
- Bardia, A.; Hurvitz, S.A.; Tolaney, S.M.; Loirat, D.; Punie, K.; Oliveira, M.; Brufsky, A.; Sardesai, S.D.; Kalinsky, K.; Zelnak, A.B.; et al. Sacituzumab Govitecan in Metastatic Triple-Negative Breast Cancer. N. Engl. J. Med. 2021, 384, 1529–1541. [Google Scholar] [CrossRef]
- Hattori, M.; Masuda, N.; Takano, T.; Tsugawa, K.; Inoue, K.; Matsumoto, K.; Ishikawa, T.; Itoh, M.; Yasojima, H.; Tanabe, Y.; et al. Pembrolizumab plus chemotherapy in Japanese patients with triple-negative breast cancer: Results from KEYNOTE-355. Cancer Med. 2023, 12, 10280–10293. [Google Scholar] [CrossRef]
- Miles, D.; Gligorov, J.; André, F.; Cameron, D.; Schneeweiss, A.; Barrios, C.; Xu, B.; Wardley, A.; Kaen, D.; Andrade, L.; et al. Primary results from IMpassion131, a double-blind, placebo-controlled, randomised phase III trial of first-line paclitaxel with or without atezolizumab for unresectable locally advanced/metastatic triple-negative breast cancer. Ann. Oncol. 2021, 32, 994–1004. [Google Scholar] [CrossRef]
- Wu, S.Y.; Xu, Y.; Chen, L.; Fan, L.; Ma, X.Y.; Zhao, S.; Song, X.Q.; Hu, X.; Yang, W.T.; Chai, W.J.; et al. Combined angiogenesis and PD-1 inhibition for immunomodulatory TNBC: Concept exploration and biomarker analysis in the FUTURE-C-Plus trial. Mol. Cancer 2022, 21, 84. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Chen, H.; Xie, Y.; Zhou, X.; Wang, Y.; Zhou, J.; Long, S.; Hu, Z.; Zhang, S.; Qiu, W.; et al. Combination of CTLA-4 blockade with MUC1 mRNA nanovaccine induces enhanced anti-tumor CTL activity by modulating tumor microenvironment of triple negative breast cancer. Transl. Oncol. 2022, 15, 101298. [Google Scholar] [CrossRef]
- Huang, R.; Wang, H.; Hong, J.; Wu, J.; Huang, O.; He, J.; Chen, W.; Li, Y.; Chen, X.; Shen, K.; et al. Targeting glutamine metabolic reprogramming of SLC7A5 enhances the efficacy of anti-PD-1 in triple-negative breast cancer. Front. Immunol. 2023, 14, 1251643. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Torres, J.; Penet, M.F.; Mironchik, Y.; Krishnamachary, B.; Bhujwalla, Z.M. The PD-L1 metabolic interactome intersects with choline metabolism and inflammation. Cancer Metab. 2021, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Song, M.; Dong, Q.; Xiang, G.; Li, J.; Ma, X.; Wei, F. UBR5 promotes tumor immune evasion through enhancing IFN-γ-induced PDL1 transcription in triple negative breast cancer. Theranostics 2022, 12, 5086–5102. [Google Scholar] [CrossRef]
- Yi, M.; Wu, Y.; Niu, M.; Zhu, S.; Zhang, J.; Yan, Y.; Zhou, P.; Dai, Z.; Wu, K. Anti-TGF-β/PD-L1 bispecific antibody promotes T cell infiltration and exhibits enhanced antitumor activity in triple-negative breast cancer. J. Immunother. Cancer 2022, 10, e005543. [Google Scholar] [CrossRef] [PubMed]
- Fattori, S.; Le Roy, A.; Houacine, J.; Robert, L.; Abes, R.; Gorvel, L.; Granjeaud, S.; Rouvière, M.S.; Ben Amara, A.; Boucherit, N.; et al. CD25high Effector Regulatory T Cells Hamper Responses to PD-1 Blockade in Triple-Negative Breast Cancer. Cancer Res. 2023, 83, 3026–3044. [Google Scholar] [CrossRef]
- Stirling, E.; Wilson, A.; Cook, K.; Thomas, A.; Triozzi, P.; Soto-Pantoja, D. 616 CD47 blockade modulates immunosuppressive checkpoint molecules and cellular metabolism to sensitize triple-negative breast cancer tumors to immune checkpoint blockade therapy. J. ImmunoTherapy Cancer 2021, 9 (Suppl 2), A646. [Google Scholar] [CrossRef]
- Lu, G.; Qiu, Y.; Su, X. Targeting CXCL12-CXCR4 Signaling Enhances Immune Checkpoint Blockade Therapy Against Triple Negative Breast Cancer. Eur. J. Pharm. Sci. 2021, 157, 105606. [Google Scholar] [CrossRef]
- Nandi, D.; Parida, S.; Sharma, D. The gut microbiota in breast cancer development and treatment: The good, the bad, and the useful! Gut Microbes 2023, 15, 2221452. [Google Scholar] [CrossRef]
- Meng, Z.; Ye, Z.; Zhu, P.; Zhu, J.; Fang, S.; Qiu, T.; Li, Y.; Meng, L. New Developments and Opportunities of Microbiota in Treating Breast Cancers. Front. Microbiol. 2022, 13, 818793. [Google Scholar] [CrossRef]
- Wang, H.; Rong, X.; Zhao, G.; Zhou, Y.; Xiao, Y.; Ma, D.; Jin, X.; Wu, Y.; Yan, Y.; Yang, H.; et al. The microbial metabolite trimethylamine N-oxide promotes antitumor immunity in triple-negative breast cancer. Cell Metab. 2022, 34, 581–594. [Google Scholar] [CrossRef]
- Clear, K.; Stirling, E.; Wilson, A.; Soto-Pantoja, D.; Cook, K. 1304 Dietary patterns associated with increased abundance of Akkermansia muciniphila potentiates anti-PD-L1 immune checkpoint blockade response in triple-negative breast cancer. J. ImmunoTherapy Cancer 2022, 10, A1354. [Google Scholar] [CrossRef]


{kind=link}
| Therapeutic Approach | Potential Mechanisms and Outcomes | References |
|---|---|---|
| PD-L1 blockade (pembrolizumab) PD-1—blockade (pembrolizumab) | ORR 18.5% ORR 5.7%, modest immune response | [7] |
| PD-L1-siRNA | Increase TNF-α, IL-2, IFN-γ decrease IL-10 and TGF-β, | [48] |
| CTLA4/PD-1blockade | Increase T cell population decrease T regs cells | [53] |
| VISTA blockade (Clone 13F3) VISTA blockade+ Cycloheximide+ radiotherapy, | decrease MDSCs in TME Increase CD8+ T-cells+ decrease MDSCs | [56] |
| LAG3/PD-1 blockade | decrease tumor weight & size | [54] |
| Chemotherapy -ICIs (Pembrolizumab) | pCR up to 64.8% increase in Pembrolizumab-group/51.2% in placebo group | [60] |
| Angiogenesis/PD-1 blockade | ORR up to 81.3% in TNBC CD8+ T cells > 10% | [62] |
| CTLA4 blockade & MUC1 mRNA vaccine | Increased cytotoxic CD8+ T cells Increase IL-12 & IFNγ decrease Tregs, TNF-α, IL-6, TGF-β, STAT3 and phospho STAT3 levels. | [63] |
| UBR5/PD-L1 blockade | Increase PD-L1 & Increase immune suppression Increase survival rate | [66] |
| PD-1-TGF-β blockade | decrease collagen deposition Increase CD8+ cells & Increase TILs | [67] |
| CD25- PD-1 blockade | Increase CD8+ T cells & decrease CD25hig | [68] |
| CD47- PD-L1 blockade | Increase cytotoxic T cells | [69] |
| anti-PD1 with liposomal AMD3100 Plerixafor, | Promote an effective immune response | [69] |
| Gut Microbiota (Akkermansia muciniphila) & PD-L1 blockade | Increase efficacy of PD-L1 blockade by up to 70–40% | [74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Said, S.S.; Ibrahim, W.N. Breaking Barriers: The Promise and Challenges of Immune Checkpoint Inhibitors in Triple-Negative Breast Cancer. Biomedicines 2024, 12, 369. https://doi.org/10.3390/biomedicines12020369
Said SS, Ibrahim WN. Breaking Barriers: The Promise and Challenges of Immune Checkpoint Inhibitors in Triple-Negative Breast Cancer. Biomedicines. 2024; 12(2):369. https://doi.org/10.3390/biomedicines12020369
Chicago/Turabian StyleSaid, Sawsan Sudqi, and Wisam Nabeel Ibrahim. 2024. "Breaking Barriers: The Promise and Challenges of Immune Checkpoint Inhibitors in Triple-Negative Breast Cancer" Biomedicines 12, no. 2: 369. https://doi.org/10.3390/biomedicines12020369