
Transcriptome-Wide N6-Methyladenosine Alternations in Pulmonary Arteries of Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats and Novel Therapeutic Targets †

Abstract
:1. Introduction
2. Materials and Methods
2.1. Pulmonary Arterial Hypertension Models and Hemodynamic Measurement
2.2. Histology Staining, Immunohistochemistry, and Immunofluorescence
2.3. RNA Extraction, MeRIP-Seq and RNA-Seq
2.4. Sequencing Data Analyses
2.5. Data Collection and DEGs Identification
2.6. Protein Isolation and Western Blots
2.7. Quantitative Real-Time PCR
2.8. Primary Rat and Human PASMC Culture and siRNA Knockdown of Lrpprc
2.9. Proliferation Assays
2.10. Statistical Analysis
3. Results
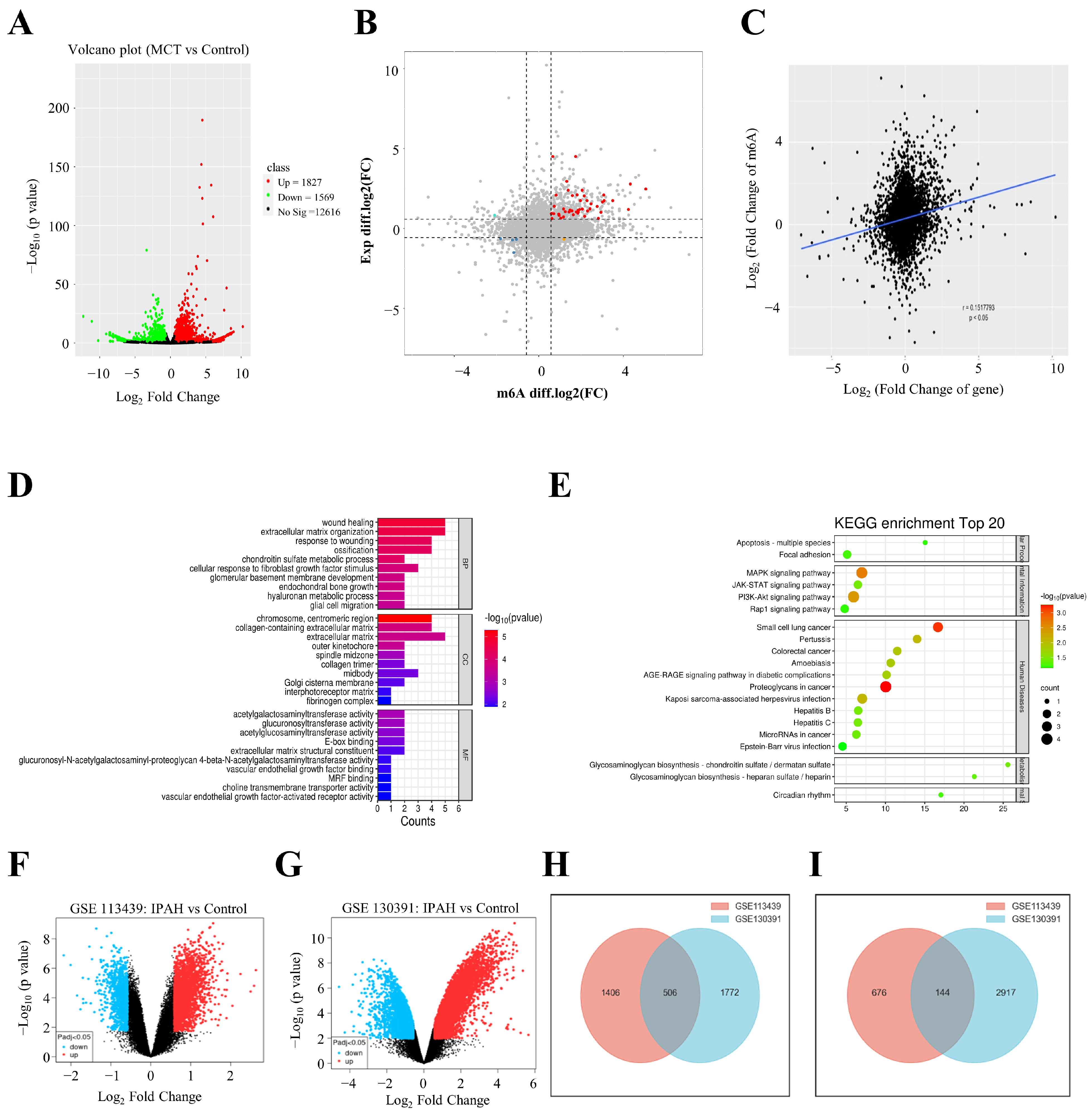
3.1. Basic Characteristics of m6A in PAH Rat Models
3.2. Differential m6A Analysis in PAH
3.3. Transcriptional Profile of m6A-Modified Genes
3.4. PPI Network Establishment and Candidate Gene Identification
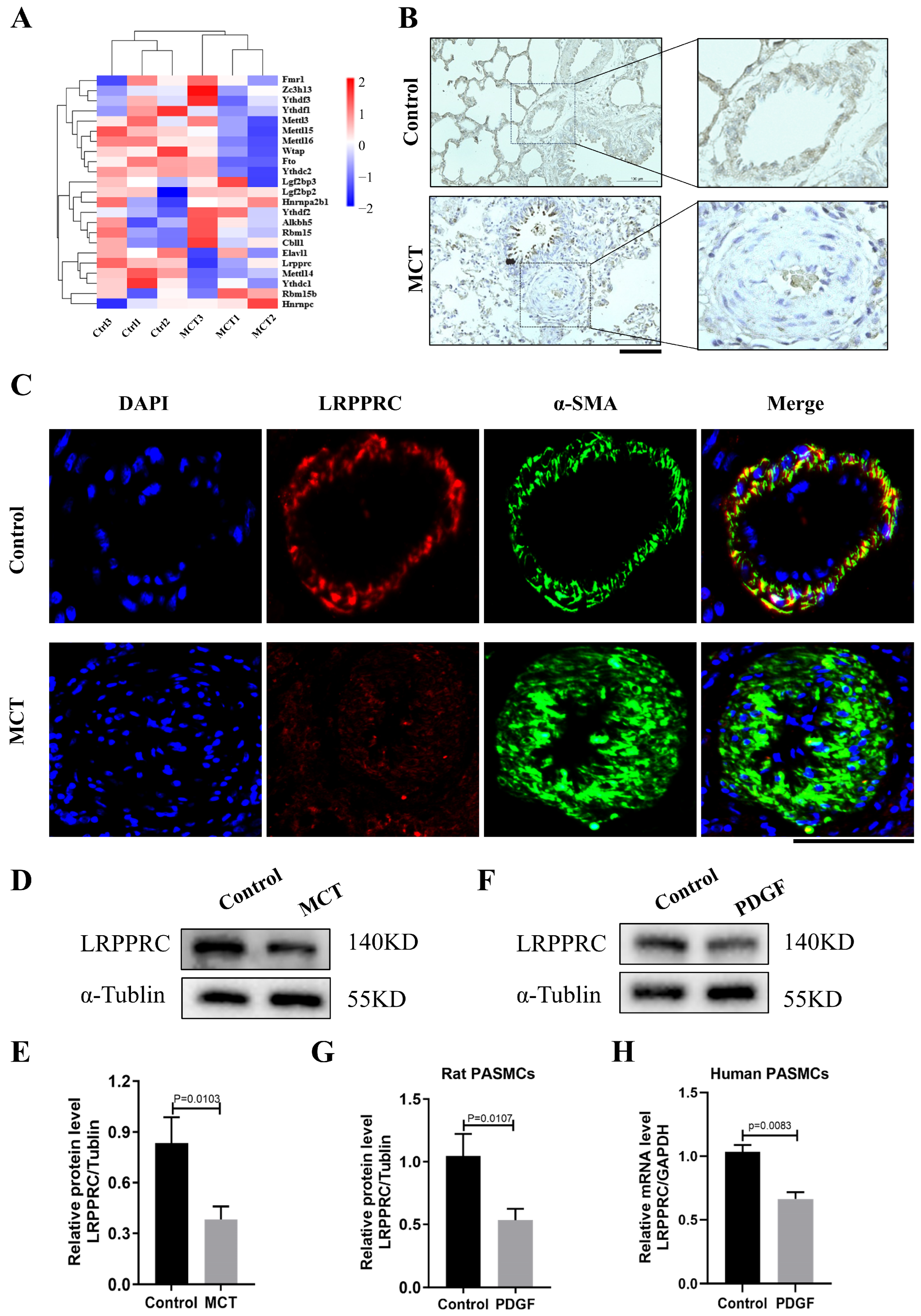
3.5. LRPPRC Is Decreased in the Pulmonary Arteries of PAH
3.6. Downregulation of LRPPRC Promotes PASMC Proliferation and Cenpf Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hodgson, J.; Swietlik, E.M.; Salmon, R.M.; Hadinnapola, C.; Nikolic, I.; Wharton, J.; Guo, J.; Liley, J.; Haimel, M.; Bleda, M.; et al. Characterization of GDF2 Mutations and Levels of BMP9 and BMP10 in Pulmonary Arterial Hypertension. Am. J. Respir. Crit. Care Med. 2020, 201, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Le Ribeuz, H.; Courboulin, A.; Ghigna, M.R.; Lambert, M.; Hautefort, A.; Humbert, M.; Montani, D.; Cohen-Kaminsky, S.; Perros, F.; Antigny, F. In vivo miR-138-5p inhibition alleviates monocrotaline-induced pulmonary hypertension and normalizes pulmonary KCNK3 and SLC45A3 expression. Respir. Res. 2020, 21, 186. [Google Scholar] [CrossRef]
- Thenappan, T.; Ormiston, M.L.; Ryan, J.J.; Archer, S.L. Pulmonary arterial hypertension: Pathogenesis and clinical management. BMJ 2018, 360, j5492. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.A.R.; Lawrie, A. Targeting Vascular Remodeling to Treat Pulmonary Arterial Hypertension. Trends Mol. Med. 2017, 23, 31–45. [Google Scholar] [CrossRef]
- Kim, J.D.; Lee, A.; Choi, J.; Park, Y.; Kang, H.; Chang, W.; Lee, M.S.; Kim, J. Epigenetic modulation as a therapeutic approach for pulmonary arterial hypertension. Exp. Mol. Med. 2015, 47, e175. [Google Scholar] [CrossRef]
- Benincasa, G.; DeMeo, D.L.; Glass, K.; Silverman, E.K.; Napoli, C. Epigenetics and pulmonary diseases in the horizon of precision medicine: A review. Eur. Respir. J. 2021, 57, 2003406. [Google Scholar] [CrossRef]
- Zhao, B.S.; Roundtree, I.A.; He, C. Post-transcriptional gene regulation by mRNA modifications. Nat. Rev. Mol. Cell Biol. 2017, 18, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.X.; Zhang, X.S.; Sui, N. Advances in the profiling of N6-methyladenosine (m6A) modifications. Biotechnol. Adv. 2020, 45, 107656. [Google Scholar] [CrossRef]
- He, P.C.; He, C. m6A RNA methylation: From mechanisms to therapeutic potential. EMBO J. 2021, 40, e105977. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, B.; Nie, Z.; Duan, L.; Xiong, Q.; Jin, Z.; Yang, C.; Chen, Y. The role of m6A modification in the biological functions and diseases. Signal Transduct. Target. Ther. 2021, 6, 74. [Google Scholar] [CrossRef]
- Eichstaedt, C.A.; Song, J.; Benjamin, N.; Harutyunova, S.; Fischer, C.; Grunig, E.; Hinderhofer, K. EIF2AK4 mutation as “second hit” in hereditary pulmonary arterial hypertension. Respir. Res. 2016, 17, 141. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hsu, P.J.; Chen, Y.S.; Yang, Y.G. Dynamic transcriptomic m6A decoration: Writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018, 28, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Zhang, J.; Zhu, J.S. The role of m6A RNA methylation in human cancer. Mol. Cancer 2019, 18, 103. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Flavell, R.A.; Li, H.B. RNA m6A modification and its function in diseases. Front. Med. 2018, 12, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Lv, B.; Qin, Y.; Zhang, B. Emerging Roles and Mechanism of m6A Methylation in Cardiometabolic Diseases. Cells 2022, 11, 1101. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tang, A.; Cui, Y.; Gong, H.; Li, H. LRPPRC facilitates tumor progression and immune evasion through upregulation of m6A modification of PD-L1 mRNA in hepatocellular carcinoma. Front. Immunol. 2023, 14, 1144774. [Google Scholar] [CrossRef] [PubMed]
- Pinello, N.; Sun, S.; Wong, J.J. Aberrant expression of enzymes regulating m6A mRNA methylation: Implication in cancer. Cancer Biol. Med. 2018, 15, 323–334. [Google Scholar] [CrossRef]
- Fu, Y.; Dominissini, D.; Rechavi, G.; He, C. Gene expression regulation mediated through reversible m6A RNA methylation. Nat. Rev. Genet. 2014, 15, 293–306. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, Q.; Li, B.; Wang, D.; Wang, L.; Zhou, Y.L. m6A regulator-mediated methylation modification patterns and tumor microenvironment infiltration characterization in gastric cancer. Mol. Cancer 2020, 19, 53. [Google Scholar] [CrossRef]
- Xia, Y.; Zhang, Y.; Huang, J.; Chen, B.; Jiang, Y.; Sun, Z.; Liu, Y. N6-Methyladenosine Modifications in Pulmonary Hypertension. Pharmacology 2023, 108, 497–503. [Google Scholar] [CrossRef]
- Qin, Y.; Qiao, Y.; Li, L.; Luo, E.; Wang, D.; Yao, Y.; Tang, C.; Yan, G. The m6A methyltransferase METTL3 promotes hypoxic pulmonary arterial hypertension. Life Sci. 2021, 274, 119366. [Google Scholar] [CrossRef]
- Su, H.; Wang, G.; Wu, L.; Ma, X.; Ying, K.; Zhang, R. Transcriptome-wide map of m6A circRNAs identified in a rat model of hypoxia mediated pulmonary hypertension. BMC Genom. 2020, 21, 39. [Google Scholar] [CrossRef]
- Xu, S.; Xu, X.; Zhang, Z.; Yan, L.; Zhang, L.; Du, L. The role of RNA m6A methylation in the regulation of postnatal hypoxia-induced pulmonary hypertension. Respir. Res. 2021, 22, 121. [Google Scholar] [CrossRef]
- Boucly, A.; Gerges, C.; Savale, L.; Jais, X.; Jevnikar, M.; Montani, D.; Sitbon, O.; Humbert, M. Pulmonary arterial hypertension. Presse Med. 2023, 52, 104168. [Google Scholar] [CrossRef]
- Beh, L.Y.; Debelouchina, G.T.; Clay, D.M.; Thompson, R.E.; Lindblad, K.A.; Hutton, E.R.; Bracht, J.R.; Sebra, R.P.; Muir, T.W.; Landweber, L.F. Identification of a DNA N6-Adenine Methyltransferase Complex and Its Impact on Chromatin Organization. Cell 2019, 177, 1781–1796.e1725. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Meng, J.; Cui, X.; Rao, M.K.; Chen, Y.; Huang, Y. Exome-based analysis for RNA epigenome sequencing data. Bioinformatics 2013, 29, 1565–1567. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Lu, Z.; Liu, H.; Zhang, L.; Zhang, S.; Chen, Y.; Rao, M.K.; Huang, Y. A protocol for RNA methylation differential analysis with MeRIP-Seq data and exomePeak R/Bioconductor package. Methods 2014, 69, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; He, Q.Y. ChIPseeker: An R/Bioconductor package for ChIP peak annotation, comparison and visualization. Bioinformatics 2015, 31, 2382–2383. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Veno, M.T.; Damgaard, C.K.; Kjems, J. Comparison of circular RNA prediction tools. Nucleic Acids Res. 2016, 44, e58. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdottir, H.; Turner, D.; Mesirov, J.P. igv.js: An embeddable JavaScript implementation of the Integrative Genomics Viewer (IGV). Bioinformatics 2023, 39, btac830. [Google Scholar] [CrossRef] [PubMed]
- Mura, M.; Cecchini, M.J.; Joseph, M.; Granton, J.T. Osteopontin lung gene expression is a marker of disease severity in pulmonary arterial hypertension. Respirology 2019, 24, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Halliday, S.J.; Matthews, D.T.; Talati, M.H.; Austin, E.D.; Su, Y.R.; Absi, T.S.; Fortune, N.L.; Gailani, D.; Matafonov, A.; West, J.D.; et al. A multifaceted investigation into molecular associations of chronic thromboembolic pulmonary hypertension pathogenesis. JRSM Cardiovasc. Dis. 2020, 9, 2048004020906994. [Google Scholar] [CrossRef]
- Zong, X.; Xiao, X.; Shen, B.; Jiang, Q.; Wang, H.; Lu, Z.; Wang, F.; Jin, M.; Min, J.; Wang, F.; et al. The N6-methyladenosine RNA-binding protein YTHDF1 modulates the translation of TRAF6 to mediate the intestinal immune response. Nucleic Acids Res. 2021, 49, 5537–5552. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Hu, L.; Wang, J.; Huang, H.; Yu, Y.; Ding, J.; Yu, Y.; Li, K.; Wei, D.; Ye, Q.; Wang, F.; et al. YTHDF1 Regulates Pulmonary Hypertension through Translational Control of MAGED1. Am. J. Respir. Crit. Care Med. 2021, 203, 1158–1172. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zeng, Y.; Yang, Y.; Fan, H.; Deng, Y.; Chen, W.; Liu, J.; Yang, F.; Li, W.; Xiao, Y. Comprehensive analysis of m6A methylomes in idiopathic pulmonary arterial hypertension. Epigenetics 2023, 18, 2242225. [Google Scholar] [CrossRef]
- Zeng, Y.; Huang, T.; Zuo, W.; Wang, D.; Xie, Y.; Wang, X.; Xiao, Z.; Chen, Z.; Liu, Q.; Liu, N.; et al. Integrated analysis of m6A mRNA methylation in rats with monocrotaline-induced pulmonary arterial hypertension. Aging 2021, 13, 18238–18256. [Google Scholar] [CrossRef]
- Thenappan, T.; Chan, S.Y.; Weir, E.K. Role of extracellular matrix in the pathogenesis of pulmonary arterial hypertension. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1322–H1331. [Google Scholar] [CrossRef]
- Hussein, D.; Taylor, S.S. Farnesylation of Cenp-F is required for G2/M progression and degradation after mitosis. J. Cell Sci. 2002, 115 Pt 17, 3403–3414. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Liu, L.; Zeng, T.; Zhu, Y.H.; Li, J.; Chen, L.; Li, Y.; Yuan, Y.F.; Ma, S.; Guan, X.Y. Characterization of the oncogenic function of centromere protein F in hepatocellular carcinoma. Biochem. Biophys. Res. Commun. 2013, 436, 711–718. [Google Scholar] [CrossRef]
- Yang, X.; Miao, B.S.; Wei, C.Y.; Dong, R.Z.; Gao, P.T.; Zhang, X.Y.; Lu, J.C.; Gao, C.; Wang, X.Y.; Sun, H.C.; et al. Lymphoid-specific helicase promotes the growth and invasion of hepatocellular carcinoma by transcriptional regulation of centromere protein F expression. Cancer Sci. 2019, 110, 2133–2144. [Google Scholar] [CrossRef] [PubMed]
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, writing and erasing mRNA methylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 608–624. [Google Scholar] [CrossRef]
- Siira, S.J.; Spahr, H.; Shearwood, A.J.; Ruzzenente, B.; Larsson, N.G.; Rackham, O.; Filipovska, A. LRPPRC-mediated folding of the mitochondrial transcriptome. Nat. Commun. 2017, 8, 1532. [Google Scholar] [CrossRef] [PubMed]
- Chujo, T.; Ohira, T.; Sakaguchi, Y.; Goshima, N.; Nomura, N.; Nagao, A.; Suzuki, T. LRPPRC/SLIRP suppresses PNPase-mediated mRNA decay and promotes polyadenylation in human mitochondria. Nucleic Acids Res. 2012, 40, 8033–8047. [Google Scholar] [CrossRef]
- Wei, W.S.; Wang, N.; Deng, M.H.; Dong, P.; Liu, J.Y.; Xiang, Z.; Li, X.D.; Li, Z.Y.; Liu, Z.H.; Peng, Y.L.; et al. LRPPRC regulates redox homeostasis via the circANKHD1/FOXM1 axis to enhance bladder urothelial carcinoma tumorigenesis. Redox Biol. 2021, 48, 102201. [Google Scholar] [CrossRef]
- Song, K.; Li, B.; Chen, Y.Y.; Wang, H.; Liu, K.C.; Tan, W.; Zou, J. LRPPRC regulates metastasis and glycolysis by modulating autophagy and the ROS/HIF1-alpha pathway in retinoblastoma. Mol. Ther. Oncolytics 2021, 22, 582–591. [Google Scholar] [CrossRef]
- Li, W.; Dai, Y.; Shi, B.; Yue, F.; Zou, J.; Xu, G.; Jiang, X.; Wang, F.; Zhou, X.; Liu, L. LRPPRC sustains Yap-P27-mediated cell ploidy and P62-HDAC6-mediated autophagy maturation and suppresses genome instability and hepatocellular carcinomas. Oncogene 2020, 39, 3879–3892. [Google Scholar] [CrossRef]
- Tian, T.; Ikeda, J.; Wang, Y.; Mamat, S.; Luo, W.; Aozasa, K.; Morii, E. Role of leucine-rich pentatricopeptide repeat motif-containing protein (LRPPRC) for anti-apoptosis and tumourigenesis in cancers. Eur. J. Cancer 2012, 48, 2462–2473. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | Sample | GEO | Platform | IPAH | Control |
|---|---|---|---|---|---|
| Mura, M. et al. [33]. | Lung tissue | GSE113439 | GPL6244 | 6 | 11 |
| Halliday, S. et al. [34]. | Lung tissue | GSE130391 | GPL570 | 4 | 4 |
| Gene (Species) | Primer | Gene Sequence |
|---|---|---|
| β-actin (rat) | Forward | ACATCCGTAAAGACCTCTATGCC |
| Reverse | TACTCCTGCTTGCTGATCCAC | |
| Cenpf (rat) | Forward | TTTGTGAGGAGCTCGGCG |
| Reverse | AGCATCTAGAAGTAAACTGAGCG | |
| Lrpprc (rat) | Forward | TGGTCTTCATCAACAACATTGCTCTG |
| Reverse | GGCTCCATCTGCTCCTCTATCACT | |
| GAPDH (human) | Forward | CTTCGCTCTCTGCTCCTCCTGTTCG |
| Reverse | ACCAGGCGCCCAATACGACCAAAT | |
| CENPF (human) | Forward | CGTCCCCGAGAGCAAGTTTA |
| Reverse | GTAGGCAGCCCTTCTTTCCA | |
| LRPPRC (human) | Forward | GACGTTCGAGCAATGGCAG |
| Reverse | CTCAGTAGTCCTCCGGCCAC | |
| Cenpf: centromere protein F; Lrpprc: leucine rich pentatricopeptide repeat containing. | ||
| Pair | Hypermethylated Peaks | Hypermethylated Genes | Hypomethylated Peaks | Hypomethylated Genes |
|---|---|---|---|---|
| MCT-Control | 173 | 164 | 45 | 45 |
| Gene ID | DGEs | GSE113439 | GSE130391 |
|---|---|---|---|
| CENPF | 2.46979 | 0.775 | 0.945 |
| MKI67 | 2.39000 | 0.747 | 1.271 |
| PARP14 | 0.81945 | 0.886 | 1.321 |
| TRPS1 | 0.64099 | 0.750 | 1.860 |
| Gene ID | Regulation | Base Mean | Log2(FC) | p |
|---|---|---|---|---|
| Mettl14 | Writer | 866.635 | −0.475 | 0.008 |
| Mettl15 | Writer | 97.199 | −0.993 | 0.020 |
| Mettl16 | Writer | 463.047 | −0.652 | 0.007 |
| Lrpprc | Reader | 1884.530 | −0.820 | 0.002 |
| Hnrnpc | Reader | 399.525 | 1.345 | 0.021 |
| Ythdf2 | Reader | 688.647 | 0.494 | 0.043 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Y.; Yu, Z.; Tang, M.; Li, J.; Peng, B.; Juaiti, M.; Tang, Y.; Liang, B.; Ouyang, M.; Liu, Q.; et al. Transcriptome-Wide N6-Methyladenosine Alternations in Pulmonary Arteries of Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats and Novel Therapeutic Targets. Biomedicines 2024, 12, 364. https://doi.org/10.3390/biomedicines12020364
Feng Y, Yu Z, Tang M, Li J, Peng B, Juaiti M, Tang Y, Liang B, Ouyang M, Liu Q, et al. Transcriptome-Wide N6-Methyladenosine Alternations in Pulmonary Arteries of Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats and Novel Therapeutic Targets. Biomedicines. 2024; 12(2):364. https://doi.org/10.3390/biomedicines12020364
Chicago/Turabian StyleFeng, Yilu, Zaixin Yu, Mi Tang, Jiang Li, Baohua Peng, Mukamengjiang Juaiti, Yiyang Tang, Benhui Liang, Mingqi Ouyang, Qingqing Liu, and et al. 2024. "Transcriptome-Wide N6-Methyladenosine Alternations in Pulmonary Arteries of Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats and Novel Therapeutic Targets" Biomedicines 12, no. 2: 364. https://doi.org/10.3390/biomedicines12020364