
Fibromyalgia Animal Models Using Intermittent Cold and Psychological Stress

Abstract
:1. Introduction
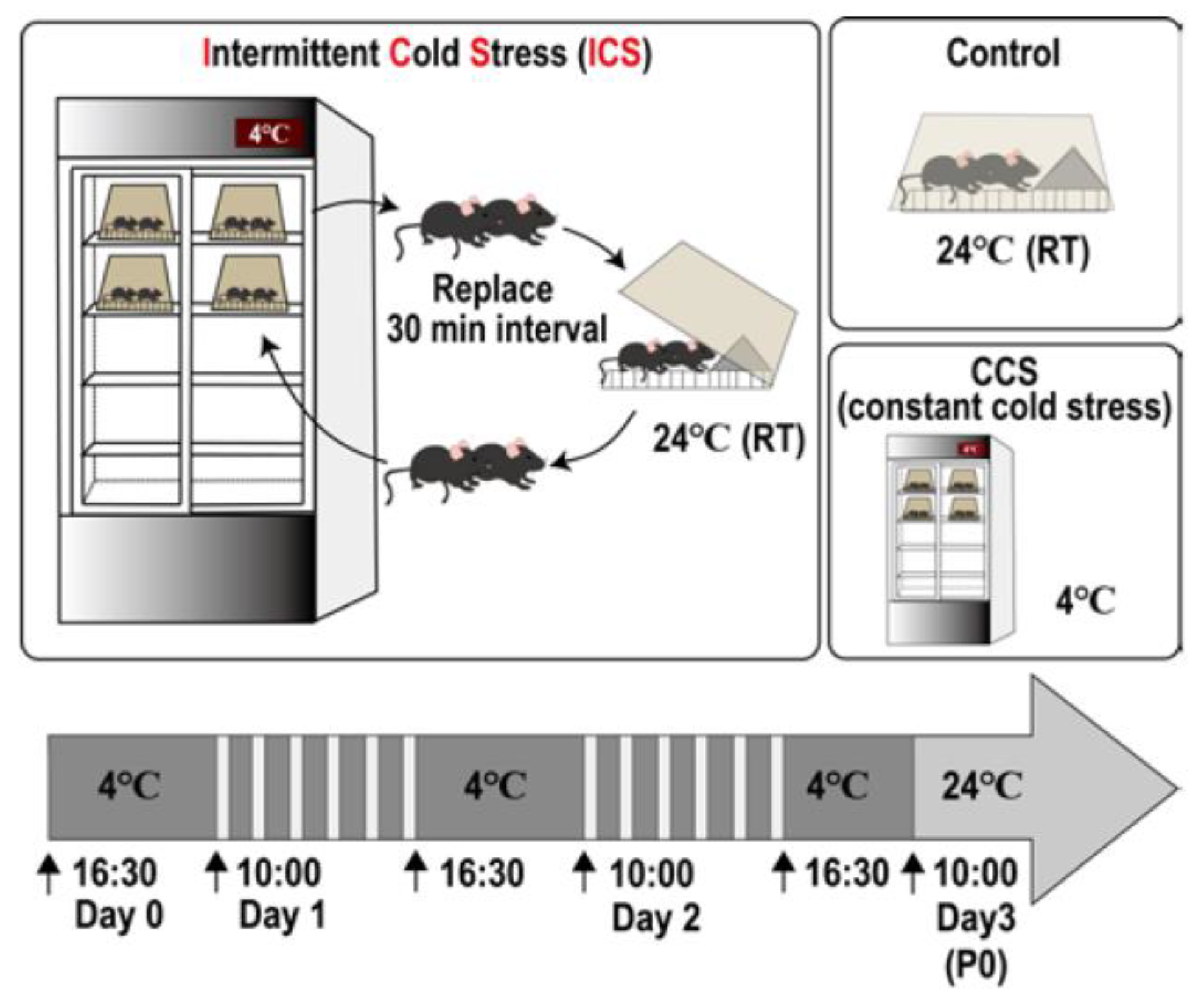
2. Intermittent Cold Stress (ICS)-Induced Generalized Pain (ICGP) Model
2.1. Protocol [51]
2.2. Assessment of Stress-Related Behaviors
2.3. Measurement of Plasma Corticosterone
2.4. Long-Term and Generalized Pain
2.5. Sex Difference
2.6. Involvement of Lysophosphatidic Acid (LPA) Receptor Signaling
2.7. Pharmacotherapy
2.7.1. Morphine
2.7.2. Antidepressants
2.7.3. Pregabalin (PGB)
2.7.4. Donepezil
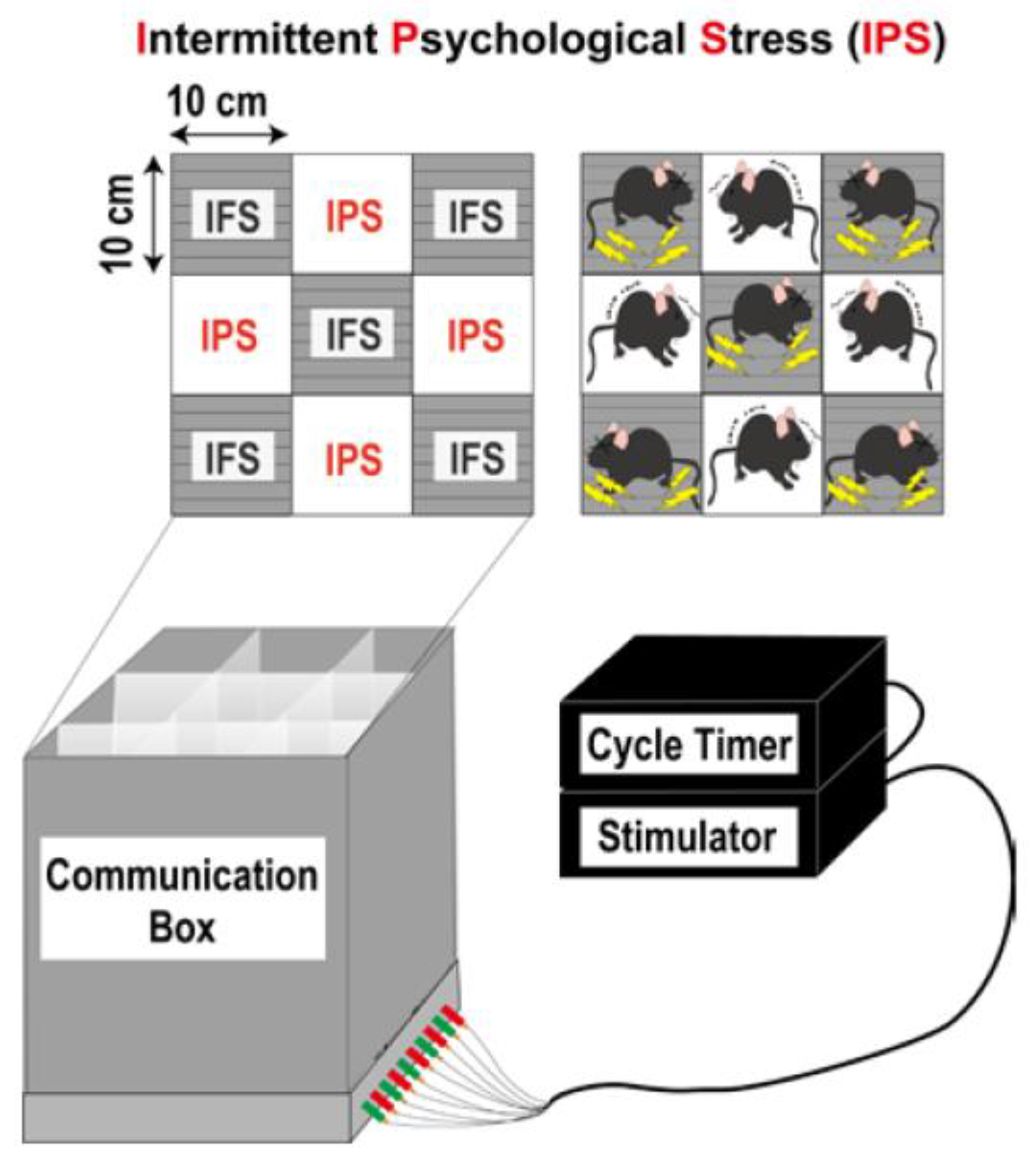
3. Intermittent Psychological Stress (IPS)-Induced Generalized Pain (IPGP) Model
3.1. Protocol [56]
3.2. Behavioral Observation
3.3. Corticosterone Levels
3.4. Long-Term IPGP
3.5. Sex Difference
3.6. Involvement of LPAR1 Signaling
3.7. Monoamine Turnover Rate
3.8. Pharmacotherapy
3.8.1. Lack of Actions of Diclofenac and Morphine
3.8.2. Pregabalin (PGB)
3.8.3. Duloxetine
3.8.4. Mirtazapine
4. Detailed Mechanisms of ICGP and IPGP
4.1. ICGP and IPGP Model Mimic the Patho-Physiological Features of FM
4.2. ICGP and IPGP Model Mimic the Pharmacotherapeutic Features of FM
4.3. Mechanisms Underlying the Loss of Central Morphine Analgesia in the ICGP Model
4.4. Potential Radical Treatments for FM
4.5. Differential Mechanisms between Generalized Pain and Loss of Central Morphine Analgesia
4.6. Relationship to Immunity
4.7. Relationship to Small Fiber Neuropathy
4.8. Limitations and Future Study
4.9. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Bidonde, J.; Fisher, E.; Perrot, S.; Moore, R.A.; Bell, R.F.; Makri, S.; Hauser, W. Effectiveness of non-pharmacological interventions for fibromyalgia and quality of review methods: An overview of Cochrane Reviews. Semin. Arthritis Rheum. 2023, 63, 152248. [Google Scholar] [CrossRef] [PubMed]
- Fitzcharles, M.A.; Cohen, S.P.; Clauw, D.J.; Littlejohn, G.; Usui, C.; Hauser, W. Nociplastic pain: Towards an understanding of prevalent pain conditions. Lancet 2021, 397, 2098–2110. [Google Scholar] [CrossRef] [PubMed]
- Galvez-Sanchez, C.M.; Duschek, S.; Reyes Del Paso, G.A. Psychological impact of fibromyalgia: Current perspectives. Psychol. Res. Behav. Manag. 2019, 12, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Gupta, H.; Girma, B.; Jenkins, J.S.; Kaufman, S.E.; Lee, C.A.; Kaye, A.D. Milnacipran for the Treatment of Fibromyalgia. Health Psychol. Res. 2021, 9, 25532. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Choi, S.E.; Park, D.J.; Lee, S.S. Disentangling Diagnosis and Management of Fibromyalgia. J. Rheum. Dis. 2022, 29, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, F.; Clauw, D.J.; Fitzcharles, M.A.; Goldenberg, D.L.; Katz, R.S.; Mease, P.; Russell, A.S.; Russell, I.J.; Winfield, J.B.; Yunus, M.B. The American College of Rheumatology preliminary diagnostic criteria for fibromyalgia and measurement of symptom severity. Arthritis Care Res. 2010, 62, 600–610. [Google Scholar] [CrossRef]
- Wolfe, F.; Smythe, H.A.; Yunus, M.B.; Bennett, R.M.; Bombardier, C.; Goldenberg, D.L.; Tugwell, P.; Campbell, S.M.; Abeles, M.; Clark, P.; et al. The American College of Rheumatology 1990 Criteria for the Classification of Fibromyalgia. Report of the Multicenter Criteria Committee. Arthritis Rheum. 1990, 33, 160–172. [Google Scholar] [CrossRef]
- Gracely, R.H.; Petzke, F.; Wolf, J.M.; Clauw, D.J. Functional magnetic resonance imaging evidence of augmented pain processing in fibromyalgia. Arthritis Rheum. 2002, 46, 1333–1343. [Google Scholar] [CrossRef]
- Hauser, W.; Ablin, J.; Fitzcharles, M.A.; Littlejohn, G.; Luciano, J.V.; Usui, C.; Walitt, B. Fibromyalgia. Nat. Rev. Dis. Primers 2015, 1, 15022. [Google Scholar] [CrossRef]
- McBeth, J.; Mulvey, M.R. Fibromyalgia: Mechanisms and potential impact of the ACR 2010 classification criteria. Nat. Rev. Rheumatol. 2012, 8, 108–116. [Google Scholar] [CrossRef]
- Wolfe, F.; Clauw, D.J.; Fitzcharles, M.A.; Goldenberg, D.L.; Hauser, W.; Katz, R.L.; Mease, P.J.; Russell, A.S.; Russell, I.J.; Walitt, B. 2016 Revisions to the 2010/2011 fibromyalgia diagnostic criteria. Semin. Arthritis Rheum. 2016, 46, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Aydede, M.; Shriver, A. Recently introduced definition of “nociplastic pain” by the International Association for the Study of Pain needs better formulation. Pain 2018, 159, 1176–1177. [Google Scholar] [CrossRef] [PubMed]
- Minhas, D.; Murphy, A.; Clauw, D.J. Fibromyalgia and centralized pain in the rheumatoid arthritis patient. Curr. Opin. Rheumatol. 2023, 35, 170–174. [Google Scholar] [CrossRef]
- Clauw, D.J.; Arnold, L.M.; McCarberg, B.H. The science of fibromyalgia. Mayo Clin. Proc. 2011, 86, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Russell, I.J.; Larson, A.A. Neurophysiopathogenesis of fibromyalgia syndrome: A unified hypothesis. Rheum. Dis. Clin. N. Am. 2009, 35, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.A.; Clauw, D.J. Understanding fibromyalgia: Lessons from the broader pain research community. J. Pain. 2009, 10, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Tzadok, R.; Ablin, J.N. Current and Emerging Pharmacotherapy for Fibromyalgia. Pain Res. Manag. 2020, 2020, 6541798. [Google Scholar] [CrossRef]
- Calandre, E.P.; Rico-Villademoros, F.; Slim, M. An update on pharmacotherapy for the treatment of fibromyalgia. Expert Opin. Pharmacother. 2015, 16, 1347–1368. [Google Scholar] [CrossRef]
- Carville, S.F.; Arendt-Nielsen, L.; Bliddal, H.; Blotman, F.; Branco, J.C.; Buskila, D.; Da Silva, J.A.; Danneskiold-Samsoe, B.; Dincer, F.; Henriksson, C.; et al. EULAR evidence-based recommendations for the management of fibromyalgia syndrome. Ann. Rheum. Dis. 2008, 67, 536–541. [Google Scholar] [CrossRef]
- Fitzcharles, M.A.; Ste-Marie, P.A.; Goldenberg, D.L.; Pereira, J.X.; Abbey, S.; Choiniere, M.; Ko, G.; Moulin, D.E.; Panopalis, P.; Proulx, J.; et al. 2012 Canadian Guidelines for the diagnosis and management of fibromyalgia syndrome: Executive summary. Pain Res. Manag. 2013, 18, 119–126. [Google Scholar] [CrossRef]
- Hauser, W.; Arnold, B.; Eich, W.; Felde, E.; Flugge, C.; Henningsen, P.; Herrmann, M.; Kollner, V.; Kuhn, E.; Nutzinger, D.; et al. Management of fibromyalgia syndrome—An interdisciplinary evidence-based guideline. GMS Ger. Med. Sci. 2008, 6, Doc14. [Google Scholar] [PubMed]
- Hauser, W.; Urrutia, G.; Tort, S.; Uceyler, N.; Walitt, B. Serotonin and noradrenaline reuptake inhibitors (SNRIs) for fibromyalgia syndrome. Cochrane Database Syst. Rev. 2013, 1, CD010292. [Google Scholar] [CrossRef] [PubMed]
- Quilici, S.; Chancellor, J.; Lothgren, M.; Simon, D.; Said, G.; Le, T.K.; Garcia-Cebrian, A.; Monz, B. Meta-analysis of duloxetine vs. pregabalin and gabapentin in the treatment of diabetic peripheral neuropathic pain. BMC Neurol. 2009, 9, 6. [Google Scholar] [CrossRef]
- Toth, C. Pregabalin: Latest safety evidence and clinical implications for the management of neuropathic pain. Ther. Adv. Drug Saf. 2014, 5, 38–56. [Google Scholar] [CrossRef] [PubMed]
- Arnold, L.M.; Clauw, D.J.; Wohlreich, M.M.; Wang, F.; Ahl, J.; Gaynor, P.J.; Chappell, A.S. Efficacy of duloxetine in patients with fibromyalgia: Pooled analysis of 4 placebo-controlled clinical trials. Prim. Care Companion J. Clin. Psychiatry 2009, 11, 237–244. [Google Scholar] [CrossRef]
- Ilari, S.; Passacatini, L.C.; Malafoglia, V.; Oppedisano, F.; Maiuolo, J.; Gliozzi, M.; Palma, E.; Tomino, C.; Fini, M.; Raffaeli, W.; et al. Tantali fibromyalgic supplicium: Is there any relief with the antidepressant employment? A systematic review. Pharmacol. Res. 2022, 186, 106547. [Google Scholar] [CrossRef]
- Staud, R. In fibromyalgia, amitriptyline is comparable to FDA-approved drugs for symptoms and acceptability. Ann. Intern. Med. 2022, 175, JC118. [Google Scholar] [CrossRef]
- Gilron, I.; Chaparro, L.E.; Tu, D.; Holden, R.R.; Milev, R.; Towheed, T.; DuMerton-Shore, D.; Walker, S. Combination of pregabalin with duloxetine for fibromyalgia: A randomized controlled trial. Pain 2016, 157, 1532–1540. [Google Scholar] [CrossRef]
- Thorpe, J.; Shum, B.; Moore, R.A.; Wiffen, P.J.; Gilron, I. Combination pharmacotherapy for the treatment of fibromyalgia in adults. Cochrane Database Syst. Rev. 2018, 2, CD010585. [Google Scholar] [CrossRef]
- Welsch, P.; Uceyler, N.; Klose, P.; Walitt, B.; Hauser, W. Serotonin and noradrenaline reuptake inhibitors (SNRIs) for fibromyalgia. Cochrane Database Syst. Rev. 2018, 2, CD010292. [Google Scholar] [CrossRef]
- Derry, S.; Wiffen, P.J.; Hauser, W.; Mucke, M.; Tolle, T.R.; Bell, R.F.; Moore, R.A. Oral nonsteroidal anti-inflammatory drugs for fibromyalgia in adults. Cochrane Database Syst. Rev. 2017, 3, CD012332. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, D.L.; Clauw, D.J.; Palmer, R.E.; Clair, A.G. Opioid Use in Fibromyalgia: A Cautionary Tale. Mayo Clin. Proc. 2016, 91, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.E.; Clauw, D.J.; Scott, D.J.; McLean, S.A.; Gracely, R.H.; Zubieta, J.K. Decreased central mu-opioid receptor availability in fibromyalgia. J. Neurosci. 2007, 27, 10000–10006. [Google Scholar] [CrossRef] [PubMed]
- Schrepf, A.; Harper, D.E.; Harte, S.E.; Wang, H.; Ichesco, E.; Hampson, J.P.; Zubieta, J.K.; Clauw, D.J.; Harris, R.E. Endogenous opioidergic dysregulation of pain in fibromyalgia: A PET and fMRI study. Pain 2016, 157, 2217–2225. [Google Scholar] [CrossRef] [PubMed]
- Clauw, D.J. What is the meaning of “small fiber neuropathy” in fibromyalgia? Pain 2015, 156, 2115–2116. [Google Scholar] [CrossRef] [PubMed]
- Doppler, K.; Rittner, H.L.; Deckart, M.; Sommer, C. Reduced dermal nerve fiber diameter in skin biopsies of patients with fibromyalgia. Pain 2015, 156, 2319–2325. [Google Scholar] [CrossRef] [PubMed]
- Levine, T.D.; Saperstein, D.S. Routine use of punch biopsy to diagnose small fiber neuropathy in fibromyalgia patients. Clin. Rheumatol. 2015, 34, 413–417. [Google Scholar] [CrossRef]
- Ramirez, M.; Martinez-Martinez, L.A.; Hernandez-Quintela, E.; Velazco-Casapia, J.; Vargas, A.; Martinez-Lavin, M. Small fiber neuropathy in women with fibromyalgia. An in vivo assessment using corneal confocal bio-microscopy. Semin. Arthritis Rheum. 2015, 45, 214–219. [Google Scholar] [CrossRef]
- Khasar, S.G.; Miao, F.J.; Janig, W.; Levine, J.D. Vagotomy-induced enhancement of mechanical hyperalgesia in the rat is sympathoadrenal-mediated. J. Neurosci. 1998, 18, 3043–3049. [Google Scholar] [CrossRef]
- Sluka, K.A.; Kalra, A.; Moore, S.A. Unilateral intramuscular injections of acidic saline produce a bilateral, long-lasting hyperalgesia. Muscle Nerve 2001, 24, 37–46. [Google Scholar] [CrossRef]
- Nagakura, Y.; Oe, T.; Aoki, T.; Matsuoka, N. Biogenic amine depletion causes chronic muscular pain and tactile allodynia accompanied by depression: A putative animal model of fibromyalgia. Pain 2009, 146, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Khasar, S.G.; Dina, O.A.; Green, P.G.; Levine, J.D. Sound stress-induced long-term enhancement of mechanical hyperalgesia in rats is maintained by sympathoadrenal catecholamines. J. Pain. 2009, 10, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Amit, Z.; Galina, Z.H. Stress-induced analgesia: Adaptive pain suppression. Physiol. Rev. 1986, 66, 1091–1120. [Google Scholar] [CrossRef] [PubMed]
- Butler, R.K.; Finn, D.P. Stress-induced analgesia. Prog. Neurobiol. 2009, 88, 184–202. [Google Scholar] [CrossRef] [PubMed]
- Marek, P.; Panocka, I.; Sadowski, B. Activation of anti- and pro-nociceptive mechanisms by front paw shock in spinal mice: Involvement of humoral factors. Pharmacol. Biochem. Behav. 1986, 24, 791–793. [Google Scholar] [CrossRef]
- Terman, G.W.; Shavit, Y.; Lewis, J.W.; Cannon, J.T.; Liebeskind, J.C. Intrinsic mechanisms of pain inhibition: Activation by stress. Science 1984, 226, 1270–1277. [Google Scholar] [CrossRef] [PubMed]
- Hata, T.; Kita, T.; Itoh, E.; Kawabata, A. The relationship of hyperalgesia in SART (repeated cold)-stressed animals to the autonomic nervous system. J. Auton. Pharmacol. 1988, 8, 45–52. [Google Scholar] [CrossRef]
- Hata, T.; Nishimura, Y.; Kita, T.; Itoh, E.; Kawabata, A. The abnormal open-field behavior of SART-stressed rats and effects of some drugs on it. Jpn. J. Pharmacol. 1988, 48, 479–490. [Google Scholar] [CrossRef]
- Imbe, H.; Iwai-Liao, Y.; Senba, E. Stress-induced hyperalgesia: Animal models and putative mechanisms. Front. Biosci. 2006, 11, 2179–2192. [Google Scholar] [CrossRef]
- Satoh, M.; Kuraishi, Y.; Kawamura, M. Effects of intrathecal antibodies to substance P, calcitonin gene-related peptide and galanin on repeated cold stress-induced hyperalgesia: Comparison with carrageenan-induced hyperalgesia. Pain 1992, 49, 273–278. [Google Scholar] [CrossRef]
- Nishiyori, M.; Ueda, H. Prolonged gabapentin analgesia in an experimental mouse model of fibromyalgia. Mol. Pain 2008, 4, 52. [Google Scholar] [CrossRef] [PubMed]
- Nishiyori, M.; Uchida, H.; Nagai, J.; Araki, K.; Mukae, T.; Kishioka, S.; Ueda, H. Permanent relief from intermittent cold stress-induced fibromyalgia-like abnormal pain by repeated intrathecal administration of antidepressants. Mol. Pain 2011, 7, 69. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Gao, X.; Wang, A.; Wang, Y.; Du, Y.; Li, L.; Li, M.; Li, C.; Jin, X.; Zhao, M. Depression comorbid with hyperalgesia: Different roles of neuroinflammation induced by chronic stress and hypercortisolism. J. Affect. Disord. 2019, 256, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Zenker, N.; Bernstein, D.E. The estimation of small amounts of corticosterone in rat plasma. J. Biol. Chem. 1958, 231, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Neyama, H.; Dozono, N.; Uchida, H.; Ueda, H. Mirtazapine, an alpha2 Antagonist-Type Antidepressant, Reverses Pain and Lack of Morphine Analgesia in Fibromyalgia-Like Mouse Models. J. Pharmacol. Exp. Ther. 2020, 375, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Neyama, H. LPA1 receptor involvement in fibromyalgia-like pain induced by intermittent psychological stress, empathy. Neurobiol. Pain 2017, 1, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Neyama, H.; Dozono, N.; Ueda, H. NR2A-NMDA Receptor Blockade Reverses the Lack of Morphine Analgesia Without Affecting Chronic Pain Status in a Fibromyalgia-Like Mouse Model. J. Pharmacol. Exp. Ther. 2020, 373, 103–112. [Google Scholar] [CrossRef]
- Matsumoto, M.; Xie, W.; Ma, L.; Ueda, H. Pharmacological switch in Abeta-fiber stimulation-induced spinal transmission in mice with partial sciatic nerve injury. Mol. Pain 2008, 4, 25. [Google Scholar] [CrossRef]
- Ueda, H. Peripheral mechanisms of neuropathic pain—Involvement of lysophosphatidic acid receptor-mediated demyelination. Mol. Pain 2008, 4, 11. [Google Scholar] [CrossRef]
- Koga, K.; Furue, H.; Rashid, M.H.; Takaki, A.; Katafuchi, T.; Yoshimura, M. Selective activation of primary afferent fibers evaluated by sine-wave electrical stimulation. Mol. Pain 2005, 1, 13. [Google Scholar] [CrossRef]
- Sophocleous, A.; Idris, A.I. Ovariectomy/Orchiectomy in Rodents. Methods Mol. Biol. 2019, 1914, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Rashid, M.H.; Fujita, R.; Contos, J.J.; Chun, J.; Ueda, H. Initiation of neuropathic pain requires lysophosphatidic acid receptor signaling. Nat. Med. 2004, 10, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H. Lysophosphatidic acid signaling is the definitive mechanism underlying neuropathic pain. Pain 2017, 158 (Suppl. S1), S55–S65. [Google Scholar] [CrossRef]
- Ueda, H. LPA receptor signaling as a therapeutic target for radical treatment of neuropathic pain and fibromyalgia. Pain. Manag. 2020, 10, 43–53. [Google Scholar] [CrossRef]
- Ueda, H. Pathogenic mechanisms of lipid mediator lysophosphatidic acid in chronic pain. Prog. Lipid Res. 2021, 81, 101079. [Google Scholar] [CrossRef] [PubMed]
- Nishiyori, M.; Nagai, J.; Nakazawa, T.; Ueda, H. Absence of morphine analgesia and its underlying descending serotonergic activation in an experimental mouse model of fibromyalgia. Neurosci. Lett. 2010, 472, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Crofford, L.J.; Rowbotham, M.C.; Mease, P.J.; Russell, I.J.; Dworkin, R.H.; Corbin, A.E.; Young, J.P., Jr.; LaMoreaux, L.K.; Martin, S.A.; Sharma, U.; et al. Pregabalin for the treatment of fibromyalgia syndrome: Results of a randomized, double-blind, placebo-controlled trial. Arthritis Rheum. 2005, 52, 1264–1273. [Google Scholar] [CrossRef]
- Hauser, W.; Bernardy, K.; Uceyler, N.; Sommer, C. Treatment of fibromyalgia syndrome with gabapentin and pregabalin—A meta-analysis of randomized controlled trials. Pain 2009, 145, 69–81. [Google Scholar] [CrossRef]
- Fishbain, D.A.; Cutler, R.; Rosomoff, H.L.; Rosomoff, R.S. Evidence-based data from animal and human experimental studies on pain relief with antidepressants: A structured review. Pain Med. 2000, 1, 310–316. [Google Scholar] [CrossRef]
- Millan, M.J. Descending control of pain. Prog. Neurobiol. 2002, 66, 355–474. [Google Scholar] [CrossRef]
- Anttila, S.A.; Leinonen, E.V. A review of the pharmacological and clinical profile of mirtazapine. CNS Drug Rev. 2001, 7, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Nutt, D. Mirtazapine: Pharmacology in relation to adverse effects. Acta Psychiatr. Scand. 1997, 391, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Mukae, T.; Fujita, W.; Ueda, H. P-glycoprotein inhibitors improve effective dose and time of pregabalin to inhibit intermittent cold stress-induced central pain. J. Pharmacol. Sci. 2016, 131, 64–67. [Google Scholar] [CrossRef]
- Boesch, D.; Muller, K.; Pourtier-Manzanedo, A.; Loor, F. Restoration of daunomycin retention in multidrug-resistant P388 cells by submicromolar concentrations of SDZ PSC 833, a nonimmunosuppressive cyclosporin derivative. Exp. Cell Res. 1991, 196, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.A.; Cho, H.Y.; Lee, Y.B. The effect of MDR1 G2677T/A polymorphism on pharmacokinetics of gabapentin in healthy Korean subjects. Arch. Pharmacal Res. 2007, 30, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Su, T.Z.; Feng, M.R.; Weber, M.L. Mediation of highly concentrative uptake of pregabalin by L-type amino acid transport in Chinese hamster ovary and Caco-2 cells. J. Pharmacol. Exp. Ther. 2005, 313, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Clauw, D.J. Fibromyalgia: A clinical review. JAMA 2014, 311, 1547–1555. [Google Scholar] [CrossRef]
- Bonafede, R.P.; Downey, D.C.; Bennett, R.M. An association of fibromyalgia with primary Sjogren’s syndrome: A prospective study of 72 patients. J. Rheumatol. 1995, 22, 133–136. [Google Scholar]
- Mukae, T.; Uchida, H.; Ueda, H. Donepezil reverses intermittent stress-induced generalized chronic pain syndrome in mice. J. Pharmacol. Exp. Ther. 2015, 353, 471–479. [Google Scholar] [CrossRef]
- Banks, W.A. Drug delivery to the brain in Alzheimer’s disease: Consideration of the blood-brain barrier. Adv. Drug Deliv. Rev. 2012, 64, 629–639. [Google Scholar] [CrossRef]
- Murata, Y.; Narisawa, Y.; Shimono, R.; Ohmori, H.; Mori, M.; Ohe, K.; Mine, K.; Enjoji, M. A high fat diet-induced decrease in hippocampal newly-born neurons of male mice is exacerbated by mild psychological stress using a Communication Box. J. Affect. Disord. 2017, 209, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, T.; Liang, Z.; Suzuki, H.; Kawana, S. Inhibitory effects of antipsychotic and anxiolytic agents on stress-induced degranulation of mouse dermal mast cells. Clin. Exp. Dermatol. 2010, 35, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Fields, H.L.; Basbaum, A.I. Brainstem control of spinal pain-transmission neurons. Annu. Rev. Physiol. 1978, 40, 217–248. [Google Scholar] [CrossRef] [PubMed]
- Russell, I.J.; Vaeroy, H.; Javors, M.; Nyberg, F. Cerebrospinal fluid biogenic amine metabolites in fibromyalgia/fibrositis syndrome and rheumatoid arthritis. Arthritis Rheum. 1992, 35, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Millan, M.J.; Gobert, A.; Rivet, J.M.; Adhumeau-Auclair, A.; Cussac, D.; Newman-Tancredi, A.; Dekeyne, A.; Nicolas, J.P.; Lejeune, F. Mirtazapine enhances frontocortical dopaminergic and corticolimbic adrenergic, but not serotonergic, transmission by blockade of alpha2-adrenergic and serotonin2C receptors: A comparison with citalopram. Eur. J. Neurosci. 2000, 12, 1079–1095. [Google Scholar] [CrossRef] [PubMed]
- Jorum, E. Analgesia or hyperalgesia following stress correlates with emotional behavior in rats. Pain 1988, 32, 341–348. [Google Scholar] [CrossRef]
- Cumming, J.; Mays, N.; Gribben, B. Reforming primary health care: Is New Zealand’s primary health care strategy achieving its early goals? Aust. N. Z. Health Policy 2008, 5, 24. [Google Scholar] [CrossRef]
- Inoue, M.; Mishina, M.; Ueda, H. Locus-specific rescue of GluRepsilon1 NMDA receptors in mutant mice identifies the brain regions important for morphine tolerance and dependence. J. Neurosci. 2003, 23, 6529–6536. [Google Scholar] [CrossRef]
- Ueda, H.; Ueda, M. Mechanisms underlying morphine analgesic tolerance and dependence. Front. Biosci. 2009, 14, 5260–5272. [Google Scholar] [CrossRef]
- Rubinstein, M.; Mogil, J.S.; Japon, M.; Chan, E.C.; Allen, R.G.; Low, M.J. Absence of opioid stress-induced analgesia in mice lacking beta-endorphin by site-directed mutagenesis. Proc. Natl. Acad. Sci. USA 1996, 93, 3995–4000. [Google Scholar] [CrossRef]
- Ueda, H. Systems Pathology of Neuropathic Pain and Fibromyalgia. Biol. Pharm. Bull. 2019, 42, 1773–1782. [Google Scholar] [CrossRef] [PubMed]
- Uchida, H.; Nagai, J.; Ueda, H. Lysophosphatidic acid and its receptors LPA1 and LPA3 mediate paclitaxel-induced neuropathic pain in mice. Mol. Pain 2014, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Neyama, H.; Matsushita, Y. Lysophosphatidic Acid Receptor 1- and 3-Mediated Hyperalgesia and Hypoalgesia in Diabetic Neuropathic Pain Models in Mice. Cells 2020, 9, 1906. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Neyama, H.; Nagai, J.; Matsushita, Y.; Tsukahara, T.; Tsukahara, R. Involvement of lysophosphatidic acid-induced astrocyte activation underlying the maintenance of partial sciatic nerve injury-induced neuropathic pain. Pain 2018, 159, 2170–2178. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Neyama, H.; Sasaki, K.; Miyama, C.; Iwamoto, R. Lysophosphatidic acid LPA(1) and LPA(3) receptors play roles in the maintenance of late tissue plasminogen activator-induced central poststroke pain in mice. Neurobiol. Pain 2019, 5, 100020. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.J.; Linker-Israeli, M.; Hallegua, D.; Silverman, S.; Silver, D.; Weisman, M.H. Cytokines play an aetiopathogenetic role in fibromyalgia: A hypothesis and pilot study. Rheumatology 2001, 40, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Bazzichi, L.; Rossi, A.; Massimetti, G.; Giannaccini, G.; Giuliano, T.; De Feo, F.; Ciapparelli, A.; Dell’Osso, L.; Bombardieri, S. Cytokine patterns in fibromyalgia and their correlation with clinical manifestations. Clin. Exp. Rheumatol. 2007, 25, 225–230. [Google Scholar]
- Kadetoff, D.; Lampa, J.; Westman, M.; Andersson, M.; Kosek, E. Evidence of central inflammation in fibromyalgia-increased cerebrospinal fluid interleukin-8 levels. J. Neuroimmunol. 2012, 242, 33–38. [Google Scholar] [CrossRef]
- Slade, G.D.; Conrad, M.S.; Diatchenko, L.; Rashid, N.U.; Zhong, S.; Smith, S.; Rhodes, J.; Medvedev, A.; Makarov, S.; Maixner, W.; et al. Cytokine biomarkers and chronic pain: Association of genes, transcription, and circulating proteins with temporomandibular disorders and widespread palpation tenderness. Pain 2011, 152, 2802–2812. [Google Scholar] [CrossRef]
- Wang, H.; Buchner, M.; Moser, M.T.; Daniel, V.; Schiltenwolf, M. The role of IL-8 in patients with fibromyalgia: A prospective longitudinal study of 6 months. Clin. J. Pain 2009, 25, 1–4. [Google Scholar] [CrossRef]
- Illescas-Montes, R.; Costela-Ruiz, V.J.; Melguizo-Rodriguez, L.; De Luna-Bertos, E.; Ruiz, C.; Ramos-Torrecillas, J. Application of Salivary Biomarkers in the Diagnosis of Fibromyalgia. Diagnostics 2021, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Goebel, A.; Krock, E.; Gentry, C.; Israel, M.R.; Jurczak, A.; Urbina, C.M.; Sandor, K.; Vastani, N.; Maurer, M.; Cuhadar, U.; et al. Passive transfer of fibromyalgia symptoms from patients to mice. J. Clin. Investig. 2021, 131, e144201. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Guo, T.Z.; Li, W.W.; Birklein, F.; Escolano, F.L.; Herrnberger, M.; Clark, J.D.; Kingery, W.S. C5a complement and cytokine signaling mediate the pronociceptive effects of complex regional pain syndrome patient IgM in fracture mice. Pain 2021, 162, 1400–1415. [Google Scholar] [CrossRef] [PubMed]
- Capellino, S.; Claus, M.; Watzl, C. Regulation of natural killer cell activity by glucocorticoids, serotonin, dopamine, and epinephrine. Cell. Mol. Immunol. 2020, 17, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Lassen, J.; Sturner, K.H.; Gierthmuhlen, J.; Dargvainiene, J.; Kixmuller, D.; Leypoldt, F.; Baron, R.; Hullemann, P. Protective role of natural killer cells in neuropathic pain conditions. Pain 2021, 162, 2366–2375. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Drury, G.L.; Parisien, M.; Ozdag Acarli, A.N.; Al-Aubodah, T.A.; Nijnik, A.; Wen, X.; Tugarinov, N.; Verner, M.; Klares, R., 3rd; et al. Unbiased immune profiling reveals a natural killer cell-peripheral nerve axis in fibromyalgia. Pain 2022, 163, e821–e836. [Google Scholar] [CrossRef]
- Greisen, J.; Hokland, M.; Grofte, T.; Hansen, P.O.; Jensen, T.S.; Vilstrup, H.; Tonnesen, E. Acute pain induces an instant increase in natural killer cell cytotoxicity in humans and this response is abolished by local anaesthesia. Br. J. Anaesth. 1999, 83, 235–240. [Google Scholar] [CrossRef]
- Jang, Y.; Yeom, M.Y.; Kang, E.S.; Kang, J.W.; Song, H.K. The antinociceptive effect of dexmedetomidine modulates spleen cell immunity in mice. Int. J. Med. Sci. 2014, 11, 226–233. [Google Scholar] [CrossRef]
- Schedlowski, M.; Hosch, W.; Oberbeck, R.; Benschop, R.J.; Jacobs, R.; Raab, H.R.; Schmidt, R.E. Catecholamines modulate human NK cell circulation and function via spleen-independent beta 2-adrenergic mechanisms. J. Immunol. 1996, 156, 93–99. [Google Scholar] [CrossRef]
- Sharify, A.; Mahmoudi, M.; Izad, M.H.; Hosseini, M.J.; Sharify, M. Effect of acute pain on splenic NK cell activity, lymphocyte proliferation and cytokine production activities. Immunopharmacol. Immunotoxicol. 2007, 29, 465–476. [Google Scholar] [CrossRef]
- Landis, C.A.; Lentz, M.J.; Tsuji, J.; Buchwald, D.; Shaver, J.L. Pain, psychological variables, sleep quality, and natural killer cell activity in midlife women with and without fibromyalgia. Brain Behav. Immun. 2004, 18, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Vitale, C.; Chiossone, L.; Cantoni, C.; Morreale, G.; Cottalasso, F.; Moretti, S.; Pistorio, A.; Haupt, R.; Lanino, E.; Dini, G.; et al. The corticosteroid-induced inhibitory effect on NK cell function reflects down-regulation and/or dysfunction of triggering receptors involved in natural cytotoxicity. Eur. J. Immunol. 2004, 34, 3028–3038. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Knudsen, E.; Wang, L.; Maghazachi, A.A. Lysophosphatidic acid induces human natural killer cell chemotaxis and intracellular calcium mobilization. Eur. J. Immunol. 2003, 33, 2083–2089. [Google Scholar] [CrossRef] [PubMed]
- Maghazachi, A.A. G protein-coupled receptors in natural killer cells. J. Leukoc. Biol. 2003, 74, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Lagadari, M.; Truta-Feles, K.; Lehmann, K.; Berod, L.; Ziemer, M.; Idzko, M.; Barz, D.; Kamradt, T.; Maghazachi, A.A.; Norgauer, J. Lysophosphatidic acid inhibits the cytotoxic activity of NK cells: Involvement of Gs protein-mediated signaling. Int. Immunol. 2009, 21, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Dozono, N.; Tanaka, K.; Kaneko, S.; Neyama, H.; Uchida, H. Allodynia by Splenocytes From Mice With Acid-Induced Fibromyalgia-Like Generalized Pain and Its Sexual Dimorphic Regulation by Brain Microglia. Front. Neurosci. 2020, 14, 600166. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.K.; Liu, I.Y.; Chang, Y.T.; Chen, Y.C.; Chen, C.C.; Yen, C.T.; Shin, H.S.; Chen, C.C. Ca(v)3.2 T-type Ca2+ channel-dependent activation of ERK in paraventricular thalamus modulates acid-induced chronic muscle pain. J. Neurosci. 2010, 30, 10360–10368. [Google Scholar] [CrossRef] [PubMed]
- Caro, X.J.; Winter, E.F. Evidence of abnormal epidermal nerve fiber density in fibromyalgia: Clinical and immunologic implications. Arthritis Rheumatol. 2014, 66, 1945–1954. [Google Scholar] [CrossRef]
- Uceyler, N.; Zeller, D.; Kahn, A.K.; Kewenig, S.; Kittel-Schneider, S.; Schmid, A.; Casanova-Molla, J.; Reiners, K.; Sommer, C. Small fibre pathology in patients with fibromyalgia syndrome. Brain 2013, 136, 1857–1867. [Google Scholar] [CrossRef]
- Oaklander, A.L. What is the meaning of “small-fiber polyneuropathy” in fibromyalgia? An alternate answer. Pain 2016, 157, 1366–1367. [Google Scholar] [CrossRef]
- Gemignani, F.; Bellanova, M.F.; Saccani, E.; Pavesi, G. Non-length-dependent small fiber neuropathy: Not a matter of stockings and gloves. Muscle Nerve 2022, 65, 10–28. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Vas, P.; Rayman, G. Small Fiber Neuropathy in Diabetes Polyneuropathy: Is It Time to Change? J. Diabetes Sci. Technol. 2022, 16, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, S.; Boulton, A.J.; Dyck, P.J.; Freeman, R.; Horowitz, M.; Kempler, P.; Lauria, G.; Malik, R.A.; Spallone, V.; Vinik, A.; et al. Diabetic neuropathies: Update on definitions, diagnostic criteria, estimation of severity, and treatments. Diabetes Care 2010, 33, 2285–2293. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Pathophysiology | ICGP | IPGP | ||
|---|---|---|---|---|
| Mechanical hyperalgesia | ++ | PWT (g)@P1–12 (a) | ++ | PWT@P1, P8 (c) |
| Thermal hyperalgesia | ++ | PWL (sec)@P15 (a) | ++ | PWL@P0–19 (c) |
| Electrical hyperalgesia | ++ − | 250, 2000 Hz@P13, 12 (h) 5 Hz@P13 (h) | ||
| Depression-like behaviors | − | Tail suspension (b) | − | Tail suspension Forced swimming (c) |
| LPAR1-KO | Lost | PWL@P6 (c) | Lost | PWL@P1 (c) |
| Sex difference (after gonadectomy) | Female >> Male (a) | Female >> Male (c) | ||
| Pharmacotherapy | ICGP | IPGP | ||
| Diclofenac analgesia (i.p.) | − | PWL > 10 mg/kg (c) | ||
| Morphine analgesia (s.c.) | − | PWL > 3 mg/kg (d) | − | PWL > 1 mg/kg (c) |
| Morphine analgesia (i.c.v.) | − − | PWL > 0.3 nmol (d,e) 250, 2000 Hz > 0.3 nmol (e) | − | PWL > 0.3 nmol (c) |
| Morphine analgesia (i.t.) | ++ ++ | PWL@P5 1 nmol (d) 5 Hz@P5 1 nmol (e) | ||
| Pregabalin (i.p.) | ++ | PWL 1 mg/kg (f) | ++ | PWL 1, 3 mg/kg (c) |
| Pregabalin (i.c.v.) | +++ | PWL 1 μg (f) | +++ | PWL 0.3 μg (c) |
| Pregabalin (i.t.) | + | PWL 10 μg (f) | + | PWL 3 μg (c) |
| Duloxetine (i.p.) | ++ | PWL 30 mg/kg (c) | ||
| Duloxetine (i.c.v.) | − | PWL > 3 μg (c) | ||
| Duloxetine (i.t.) | ++ | PWL 1 μg (c) | ||
| Milnacipran (i.t.) | ++ | PWL 0.1 μg (b) | ||
| Mirtazapine (i.p.) | ++ | PWL 1 mg/kg (g) | ++ | PWL, PWT 1 mg/kg (c) |
| Mirtazapine (i.c.v.) | +++ | PWL 1 μg (g) | +++ | PWL 1 μg (c) |
| Mirtazapine (i.t.) | − | PWL > 1 μg (g) | − | PWL > 1 μg (c) |
| Donepezil (i.p.) | +++ | PWL 10, 100 μg/kg (h) | ||
| Donepezil (i.c.v.) | +++ | PWL 10 μg (h) | ||
| Donepezil (i.t.) | − | PWL > 10 μg (h) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ueda, H.; Neyama, H. Fibromyalgia Animal Models Using Intermittent Cold and Psychological Stress. Biomedicines 2024, 12, 56. https://doi.org/10.3390/biomedicines12010056
Ueda H, Neyama H. Fibromyalgia Animal Models Using Intermittent Cold and Psychological Stress. Biomedicines. 2024; 12(1):56. https://doi.org/10.3390/biomedicines12010056
Chicago/Turabian StyleUeda, Hiroshi, and Hiroyuki Neyama. 2024. "Fibromyalgia Animal Models Using Intermittent Cold and Psychological Stress" Biomedicines 12, no. 1: 56. https://doi.org/10.3390/biomedicines12010056