
Exogenous OCT4 and SOX2 Contribution to In Vitro Reprogramming in Cattle

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bovine Fetal Fibroblast (bFF) Isolation and Experimental Design
2.2. Generation of Fibroblasts Expressing Exogenous OCT4 and SOX2
2.3. Flow Cytometry Analysis
2.4. Gene Expression of Imprinted and Pluripotency Genes
2.5. Allele-Specific Methylation Analyses of the DMR at the H19/IGF2 Locus
2.6. In Vitro Induced Reprogramming into Pluripotency
2.7. Somatic Cell Nuclear Transfer
2.8. Statistical Analysis
3. Results
3.1. bFF Expressing Exogenous OCT4 and SOX2
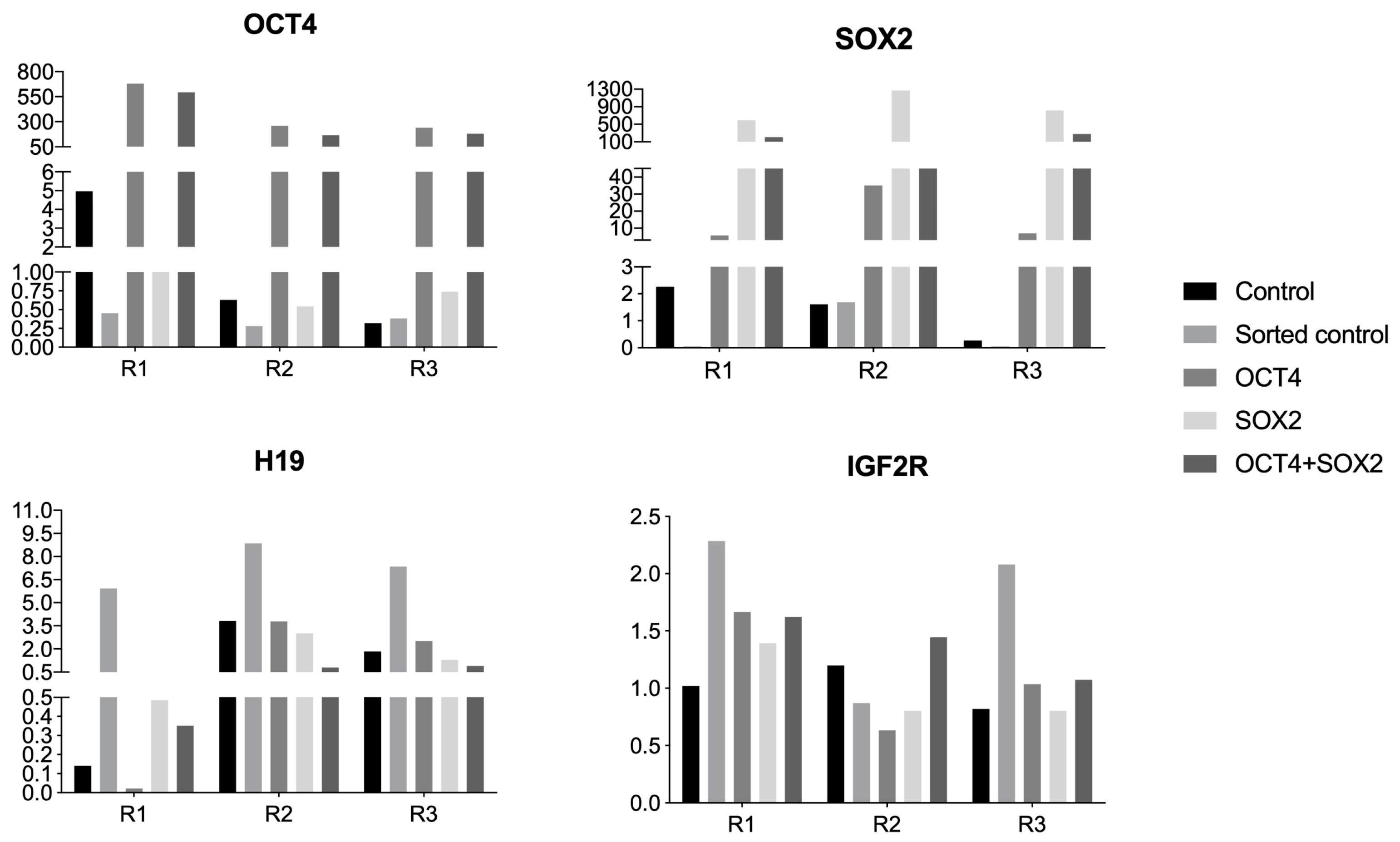
3.2. Quantitative Gene Expression Analyzes of Imprinted Genes or Genes Related to Pluripotency
3.3. Allele-Specific Methylation Analyses of the DMR at the H19/IGF2 Locus
3.4. Pluripotency Induction (iPSC Production)
3.5. Somatic Cell Nuclear Transfer Using hOCT4 and hSOX2 Overexpressing Donor Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brezina, P.R.; Ning, N.; Mitchell, E.; Zacur, H.A.; Baramki, T.A.; Zhao, Y. Recent Advances in Assisted Reproductive Technology. Curr. Obstet. Gynecol. Rep. 2012, 1, 166–173. [Google Scholar] [CrossRef]
- Herrick, J.R. Assisted Reproductive Technologies for Endangered Species Conservation: Developing Sophisticated Protocols with Limited Access to Animals with Unique Reproductive Mechanisms. Biol. Reprod. 2019, 100, 1158–1170. [Google Scholar] [CrossRef]
- Bird, A. DNA Methylation Patterns and Epigenetic Memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Bortvin, A.; Eggan, K.; Skaletsky, H.; Akutsu, H.; Berry, D.L.; Yanagimachi, R.; Page, D.C.; Jaenisch, R. Incomplete Reactivation of Oct4-Related Genes in Mouse Embryos Cloned from Somatic Nuclei. Dev. Camb. Engl. 2003, 130, 1673–1680. [Google Scholar]
- Eilertsen, K.J.; Power, R.A.; Harkins, L.L.; Misica, P. Targeting Cellular Memory to Reprogram the Epigenome, Restore Potential, and Improve Somatic Cell Nuclear Transfer. Anim. Reprod. Sci. 2007, 98, 129–146. [Google Scholar] [CrossRef]
- Santos, F.; Dean, W. Epigenetic Reprogramming during Early Development in Mammals. Reprod. Camb. Engl. 2004, 127, 643–651. [Google Scholar] [CrossRef]
- Smith, L.C.; Therrien, J.; Filion, F.; Bressan, F.; Meirelles, F.V. Epigenetic Consequences of Artificial Reproductive Technologies to the Bovine Imprinted Genes SNRPN, H19/IGF2, and IGF2R. Front. Genet. 2015, 6, 58. [Google Scholar] [CrossRef]
- Hansen, P.J. Implications of Assisted Reproductive Technologies for Pregnancy Outcomes in Mammals. Annu. Rev. Anim. Biosci. 2020, 8, 395–413. [Google Scholar] [CrossRef]
- Smith, L.C.; Suzuki, J.; Goff, A.K.; Filion, F.; Therrien, J.; Murphy, B.D.; Kohan-Ghadr, H.R.; Lefebvre, R.; Brisville, A.C.; Buczinski, S.; et al. Developmental and Epigenetic Anomalies in Cloned Cattle. Reprod. Domest. Anim. Zuchthyg. 2012, 47 (Suppl. S4), 107–114. [Google Scholar] [CrossRef]
- Brown, K.W.; Villar, A.J.; Bickmore, W.; Clayton-Smith, J.; Catchpoole, D.; Maher, E.R.; Reik, W. Imprinting Mutation in the Beckwith-Wiedemann Syndrome Leads to Biallelic IGF2 Expression through an H19-Independent Pathway. Hum. Mol. Genet. 1996, 5, 2027–2032. [Google Scholar] [CrossRef]
- Bartholdi, D.; Krajewska-Walasek, M.; Ounap, K.; Gaspar, H.; Chrzanowska, K.H.; Ilyana, H.; Kayserili, H.; Lurie, I.W.; Schinzel, A.; Baumer, A. Epigenetic Mutations of the Imprinted IGF2-H19 Domain in Silver-Russell Syndrome (SRS): Results from a Large Cohort of Patients with SRS and SRS-like Phenotypes. J. Med. Genet. 2009, 46, 192–197. [Google Scholar] [CrossRef]
- Runte, M.; Kroisel, P.M.; Gillessen-Kaesbach, G.; Varon, R.; Horn, D.; Cohen, M.Y.; Wagstaff, J.; Horsthemke, B.; Buiting, K. SNURF-SNRPN and UBE3A Transcript Levels in Patients with Angelman Syndrome. Hum. Genet. 2004, 114, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.L.; Leff, S.E. Maternal Imprinting of Human SNRPN, a Gene Deleted in Prader-Willi Syndrome. Nat. Genet. 1994, 6, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Amor, D.J.; Halliday, J. A Review of Known Imprinting Syndromes and Their Association with Assisted Reproduction Technologies. Hum. Reprod. Oxf. Engl. 2008, 23, 2826–2834. [Google Scholar] [CrossRef] [PubMed]
- Young, L.E.; Sinclair, K.D.; Wilmut, I. Large Offspring Syndrome in Cattle and Sheep. Rev. Reprod. 1998, 3, 155–163. [Google Scholar] [CrossRef]
- Ceelen, M.; Vermeiden, J.P. Health of Human and Livestock Conceived by Assisted Reproduction. Twin Res. Off. J. Int. Soc. Twin Stud. 2001, 4, 412–416. [Google Scholar] [CrossRef]
- Chen, Z.; Robbins, K.M.; Wells, K.D.; Rivera, R.M. Large Offspring Syndrome: A Bovine Model for the Human Loss-of-Imprinting Overgrowth Syndrome Beckwith-Wiedemann. Epigenetics Off. J. DNA Methylation Soc. 2013, 8, 591–601. [Google Scholar] [CrossRef]
- Nichols, J.; Zevnik, B.; Anastassiadis, K.; Niwa, H.; Klewe-Nebenius, D.; Chambers, I.; Schöler, H.; Smith, A. Formation of Pluripotent Stem Cells in the Mammalian Embryo Depends on the POU Transcription Factor Oct4. Cell 1998, 95, 379–391. [Google Scholar] [CrossRef]
- Mitsui, K.; Tokuzawa, Y.; Itoh, H.; Segawa, K.; Murakami, M.; Takahashi, K.; Maruyama, M.; Maeda, M.; Yamanaka, S. The Homeoprotein Nanog Is Required for Maintenance of Pluripotency in Mouse Epiblast and ES Cells. Cell 2003, 113, 631–642. [Google Scholar] [CrossRef]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core Transcriptional Regulatory Circuitry in Human Embryonic Stem Cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef]
- Hammachi, F.; Morrison, G.M.; Sharov, A.A.; Livigni, A.; Narayan, S.; Papapetrou, E.P.; O’Malley, J.; Kaji, K.; Ko, M.S.H.; Ptashne, M.; et al. Transcriptional Activation by Oct4 Is Sufficient for the Maintenance and Induction of Pluripotency. Cell Rep. 2012, 1, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Rizzino, A.; Wuebben, E.L. Sox2/Oct4: A Delicately Balanced Partnership in Pluripotent Stem Cells and Embryogenesis. Biochim. Biophys. Acta 2016, 1859, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Daley, G.Q. Molecular Basis of Pluripotency. Hum. Mol. Genet. 2008, 17, R23–R27. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, K.M.; Prather, R.S. Somatic Cell Nuclear Transfer Efficiency: How Can It Be Improved through Nuclear Remodeling and Reprogramming? Mol. Reprod. Dev. 2010, 77, 1001–1015. [Google Scholar] [CrossRef]
- Reik, W.; Constância, M.; Fowden, A.; Anderson, N.; Dean, W.; Ferguson-Smith, A.; Tycko, B.; Sibley, C. Regulation of Supply and Demand for Maternal Nutrients in Mammals by Imprinted Genes. J. Physiol. 2003, 547, 35–44. [Google Scholar] [CrossRef]
- Nelissen, E.C.M.; Dumoulin, J.C.M.; Busato, F.; Ponger, L.; Eijssen, L.M.; Evers, J.L.H.; Tost, J.; van Montfoort, A.P.A. Altered Gene Expression in Human Placentas after IVF/ICSI. Hum. Reprod. Oxf. Engl. 2014, 29, 2821–2831. [Google Scholar] [CrossRef]
- Constância, M.; Hemberger, M.; Hughes, J.; Dean, W.; Ferguson-Smith, A.; Fundele, R.; Stewart, F.; Kelsey, G.; Fowden, A.; Sibley, C.; et al. Placental-Specific IGF-II Is a Major Modulator of Placental and Fetal Growth. Nature 2002, 417, 945–948. [Google Scholar] [CrossRef]
- Zimmerman, D.L.; Boddy, C.S.; Schoenherr, C.S. Oct4/Sox2 Binding Sites Contribute to Maintaining Hypomethylation of the Maternal Igf2/H19 Imprinting Control Region. PLoS ONE 2013, 8, e81962. [Google Scholar] [CrossRef]
- Abi Habib, W.; Azzi, S.; Brioude, F.; Steunou, V.; Thibaud, N.; Das Neves, C.; Le Jule, M.; Chantot-Bastaraud, S.; Keren, B.; Lyonnet, S.; et al. Extensive Investigation of the IGF2/H19 Imprinting Control Region Reveals Novel OCT4/SOX2 Binding Site Defects Associated with Specific Methylation Patterns in Beckwith-Wiedemann Syndrome. Hum. Mol. Genet. 2014, 23, 5763–5773. [Google Scholar] [CrossRef]
- Pessoa, L.V.F.; Bressan, F.F.; Chiaratti, M.R.; Pires, P.R.L.; Perecin, F.; Smith, L.C.; Meirelles, F.V. Mitochondrial DNA Dynamics during in Vitro Culture and Pluripotency Induction of a Bovine Rho0 Cell Line. Genet. Mol. Res. 2015, 14, 14093–14104. [Google Scholar] [CrossRef]
- Papapetrou, E.P.; Tomishima, M.J.; Chambers, S.M.; Mica, Y.; Reed, E.; Menon, J.; Tabar, V.; Mo, Q.; Studer, L.; Sadelain, M. Stoichiometric and Temporal Requirements of Oct4, Sox2, Klf4, and c-Myc Expression for Efficient Human IPSC Induction and Differentiation. Proc. Natl. Acad. Sci. USA 2009, 106, 12759–12764. [Google Scholar] [CrossRef] [PubMed]
- Pessoa, L.V.F.; Pires, P.R.L.; Collado, M.; Pieri, N.C.G.; Recchia, K.; Souza, A.F.; Perecin, F.; Da Silveira, J.C.; Andrade, A.F.C.; Ambrosio, C.E.; et al. Generation and MiRNA Characterization of Equine Induced Pluripotent Stem Cells Derived from Fetal and Adult Multipotent Tissues. Stem Cells Int. 2019, 2019, 1393791. [Google Scholar] [CrossRef] [PubMed]
- Bressan, F.F.; Bassanezze, V.; de Figueiredo Pessôa, L.V.; Sacramento, C.B.; Malta, T.M.; Kashima, S.; Fantinato Neto, P.; Strefezzi, R.D.F.; Pieri, N.C.G.; Krieger, J.E.; et al. Generation of Induced Pluripotent Stem Cells from Large Domestic Animals. Stem Cell Res. Ther. 2020, 11, 247. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Therrien, J.; Filion, F.; Lefebvre, R.; Goff, A.K.; Perecin, F.; Meirelles, F.V.; Smith, L.C. Loss of Methylation at H19 DMD Is Associated with Biallelic Expression and Reduced Development in Cattle Derived by Somatic Cell Nuclear Transfer. Biol. Reprod. 2011, 84, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.D.S.; Bressan, F.F.; Zecchin, K.G.; Vercesi, A.E.; Mesquita, L.G.; Merighe, G.K.F.; King, W.A.; Ohashi, O.M.; Pimentel, J.R.V.; Perecin, F.; et al. Serum-starved apoptotic fibroblasts reduce blastocyst production but enable development to term after SCNT in cattle. Cloning Stem Cells 2009, 11, 565–573. [Google Scholar] [CrossRef]
- Bressan, F.F.; dos Santos Miranda, M.; Perecin, F.; De Bem, T.H.; Pereira, F.T.V.; Russo-Carbolante, E.M.; Alves, D.; Strauss, B.; Bajgelman, M.; Krieger, J.E.; et al. Improved production of genetically modified fetuses with homogeneous transgene expression after transgene integration site analysis and recloning in cattle. Cell. Reprogram. 2011, 13, 29–36. [Google Scholar] [CrossRef]
- Pan, S.; Chen, W.; Liu, X.; Xiao, J.; Wang, Y.; Liu, J.; Du, Y.; Wang, Y.; Zhang, Y. Application of a Novel Population of Multipotent Stem Cells Derived from Skin Fibroblasts as Donor Cells in Bovine SCNT. PLoS ONE 2015, 10, e0114423. [Google Scholar] [CrossRef]
- Rodríguez-Alvarez, L.; Manriquez, J.; Velasquez, A.; Castro, F.O. Constitutive Expression of the Embryonic Stem Cell Marker OCT4 in Bovine Somatic Donor Cells Influences Blastocysts Rate and Quality after Nucleus Transfer. Vitro Cell. Dev. Biol.-Anim. 2013, 49, 657–667. [Google Scholar] [CrossRef]
- Ruvinsky, A. Basics of Gametic Imprinting. J. Anim. Sci. 1999, 77 (Suppl. S2), 228–237. [Google Scholar] [CrossRef]
- Reik, W.; Dean, W.; Walter, J. Epigenetic Reprogramming in Mammalian Development. Science 2001, 293, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Urdinguio, R.G.; Sanchez-Mut, J.V.; Esteller, M. Epigenetic Mechanisms in Neurological Diseases: Genes, Syndromes, and Therapies. Lancet Neurol. 2009, 8, 1056–1072. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Hirano, K.; Nagata, S.; Tada, T. Sox2 Expression Effects on Direct Reprogramming Efficiency as Determined by Alternative Somatic Cell Fate. Stem Cell Res. 2011, 6, 177–186. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | 5′–3′ Sequence |
|---|---|
| ACTB_FWD | GCGGACAGGATGCAGAAA |
| ACTB_REV | ACGGAGTACTTGCGCTCAG |
| C2ORF29_FWD | ACTGAGCCTGACCATGCGATC |
| C2ORF29_REV | GGCTGGAGTGAGGCCAATATG |
| H19_FWD | AGTGGGAGGGGCATTGGACT |
| H19_REV | GACCATATCATATCCCTCTGTGC- |
| SOX2_FWD | ATGGGCTCGGTGGTGAAGT |
| SOX2_REV | TGGTAGTGCTGGGACATGTGA |
| OCT4_FWD | GCAAACGATCAAGCAGTGACTAC |
| OCT4_REV | GGCGCCAGAGGAGAGGATACG |
| hOCT4% | hSOX2% | hOCT4 + hSOX2% | |
|---|---|---|---|
| bFF1 | 79.8 | 10.2 | 1.3 |
| bFF2 | 22.7 | 4.2 | 0.4 |
| bFF3 | 18.7 | 3.9 | 0.2 |
| Average | 40.4 | 6.1 | 0.63 |
| Non-Sorted Control | Sorted Control | OCT4+ | SOX2+ | OCT4 + SOX2 | |
|---|---|---|---|---|---|
| Conversion rate | 98.03% | 98.23% | 95.09% | 98.89% | 98.32% |
| Non-Sorted Control (%DMR; %CTCF) | Sorted Control (%DMR, %CTCF) | OCT4+ (%DMR, %CTCF) | SOX2+ (%DMR, %CTCF) | OCT4 + SOX2 (%DMR, %CTCF) | |
|---|---|---|---|---|---|
| Maternal allele | 21.42; 33.33 | 0; 0 | 21.24; 6.67 | 2.4; 5.55 | 5.95; 11.11 |
| Paternal allele | 100; 100 | 92.85; 100 | 100; 100 | 96.42; 100 | 92.3 |
| Non-Sorted Control | Sorted Control | OCT4+ | SOX2+ | OCT4 + SOX2 | |
|---|---|---|---|---|---|
| R1 | 0.00035 (7) | 0 | 0 | 0 | 0 |
| R2 | 0.00005 (1) | 0 | 0 | 0 | 0 |
| R3 | 0 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machado, L.S.; Borges, C.M.; de Lima, M.A.; Sangalli, J.R.; Therrien, J.; Pessôa, L.V.d.F.; Fantinato Neto, P.; Perecin, F.; Smith, L.C.; Meirelles, F.V.; et al. Exogenous OCT4 and SOX2 Contribution to In Vitro Reprogramming in Cattle. Biomedicines 2023, 11, 2577. https://doi.org/10.3390/biomedicines11092577
Machado LS, Borges CM, de Lima MA, Sangalli JR, Therrien J, Pessôa LVdF, Fantinato Neto P, Perecin F, Smith LC, Meirelles FV, et al. Exogenous OCT4 and SOX2 Contribution to In Vitro Reprogramming in Cattle. Biomedicines. 2023; 11(9):2577. https://doi.org/10.3390/biomedicines11092577
Chicago/Turabian StyleMachado, Lucas Simões, Camila Martins Borges, Marina Amaro de Lima, Juliano Rodrigues Sangalli, Jacinthe Therrien, Laís Vicari de Figueiredo Pessôa, Paulo Fantinato Neto, Felipe Perecin, Lawrence Charles Smith, Flavio Vieira Meirelles, and et al. 2023. "Exogenous OCT4 and SOX2 Contribution to In Vitro Reprogramming in Cattle" Biomedicines 11, no. 9: 2577. https://doi.org/10.3390/biomedicines11092577