
Connexin 37, 40, 43 and Pannexin 1 Expression in the Gastric Mucosa of Patients with Systemic Sclerosis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Tissue Procurement and Processing
2.3. Double Immunofluorescence Staining
2.4. Immunofluorescence Signal Quantification
2.5. Statistical Analysis
3. Results
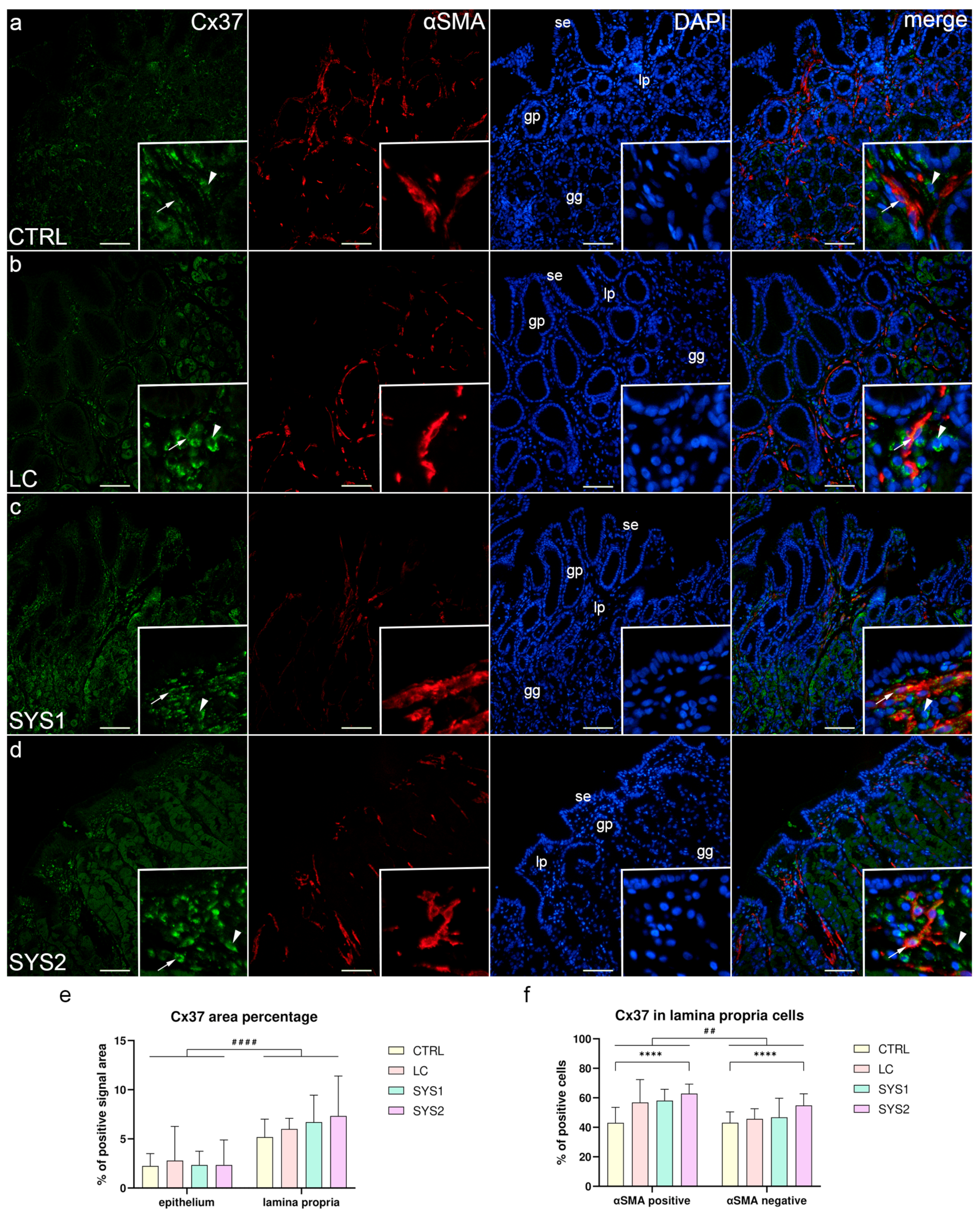
3.1. Double Immunofluorescence Staining for Cx37 and αSMA
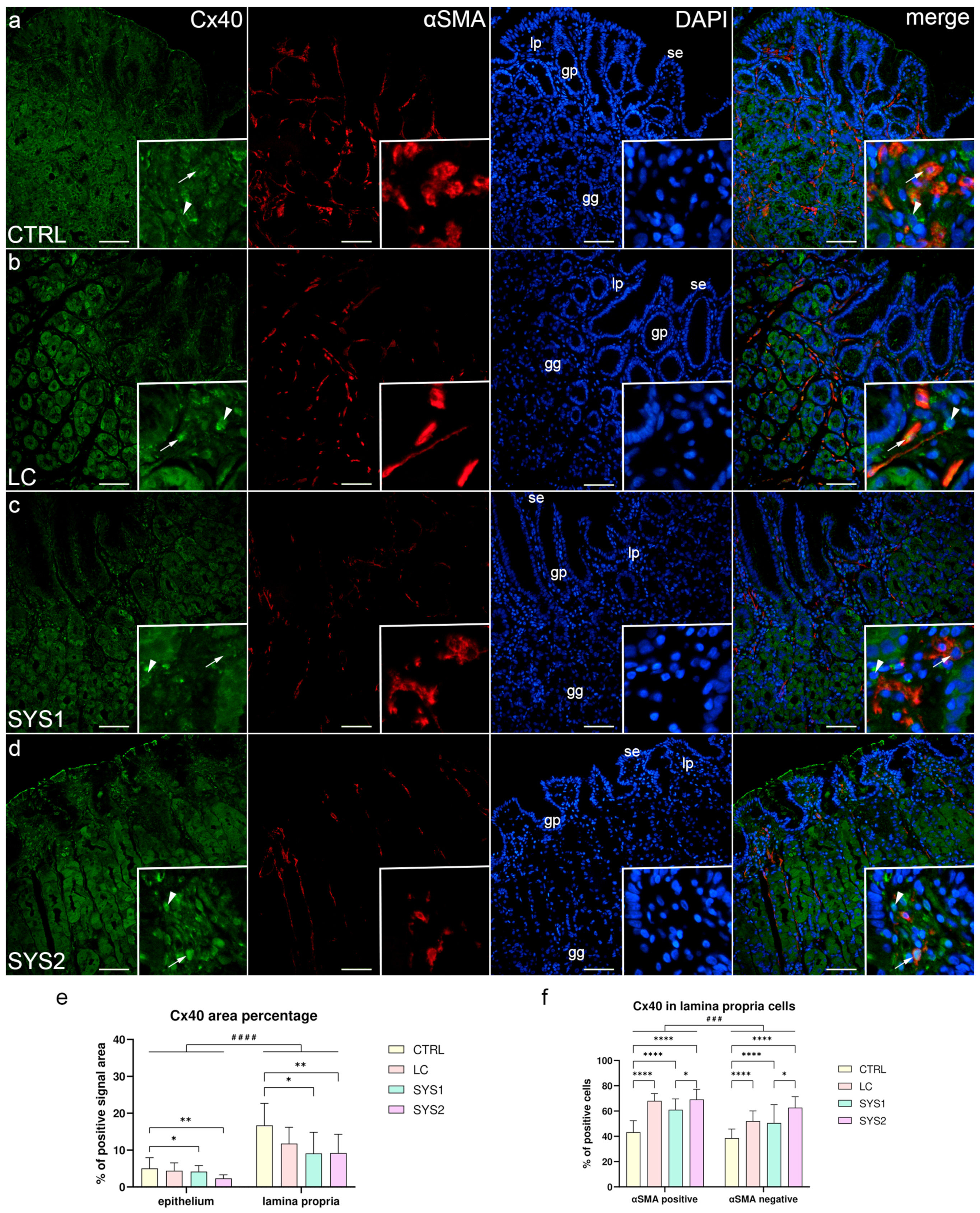
3.2. Double Immunofluorescence Staining for Cx40 and αSMA
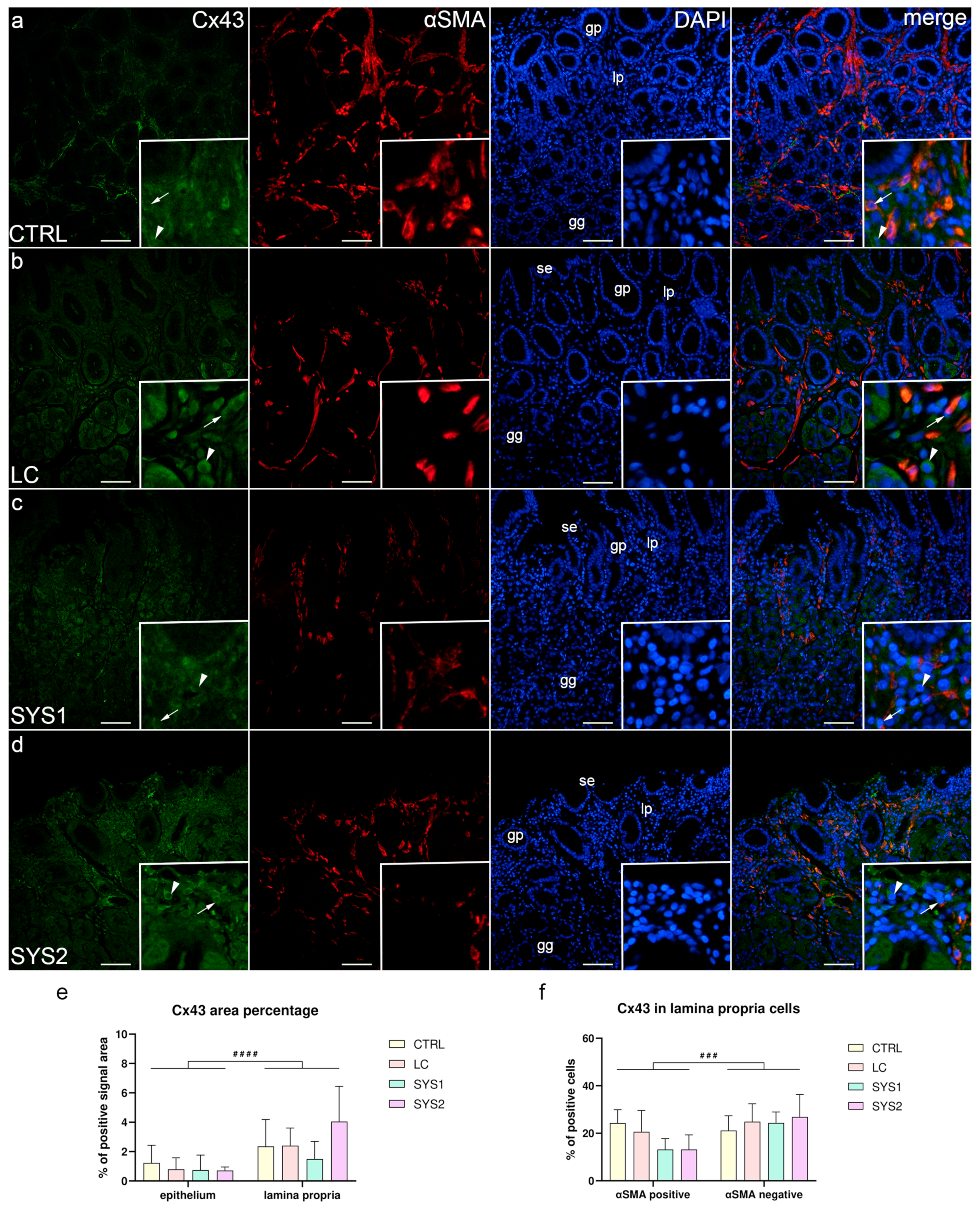
3.3. Double Immunofluorescence Staining for Cx43 and αSMA
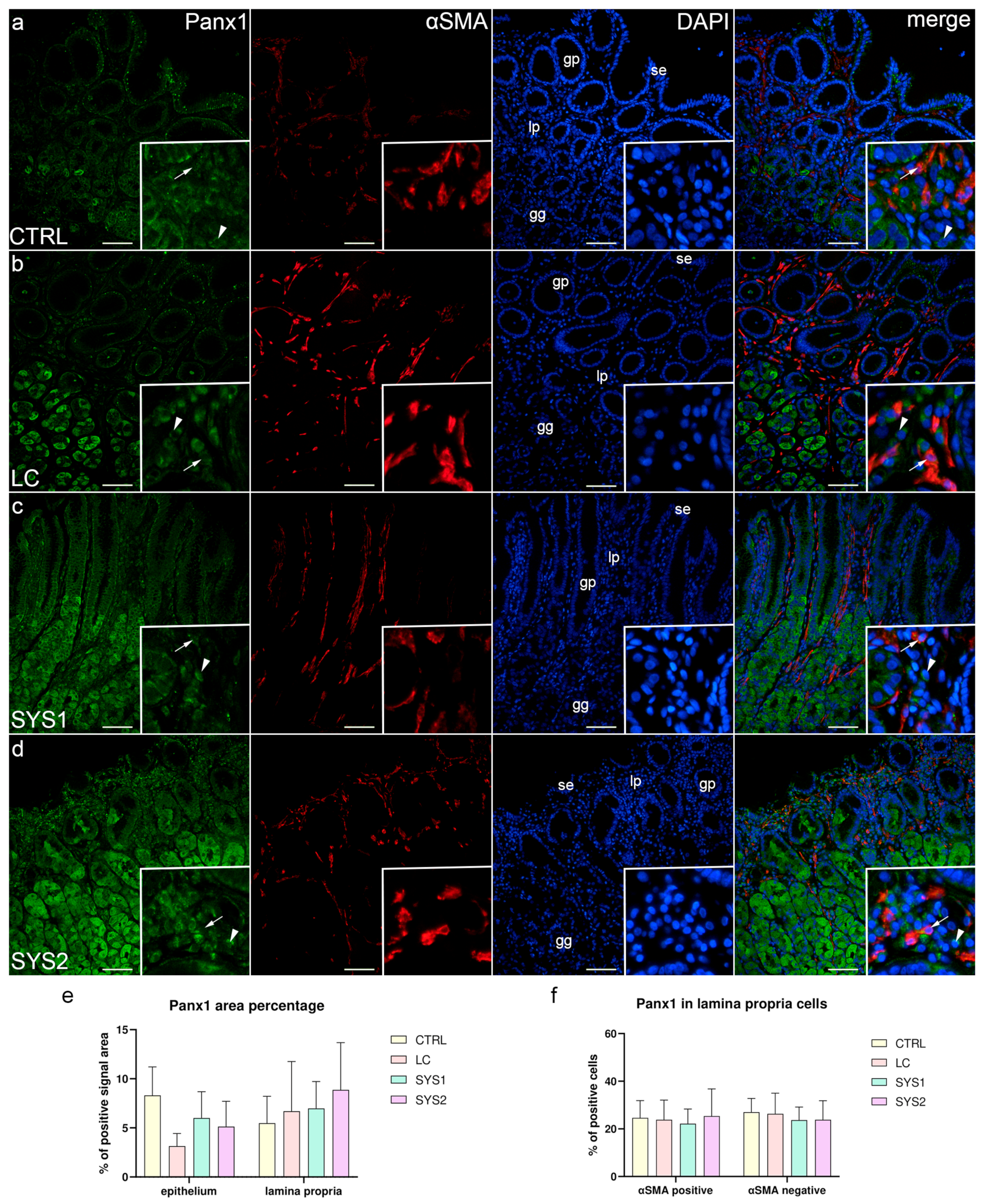
3.4. Double Immunofluorescence Staining for Panx1 and αSMA
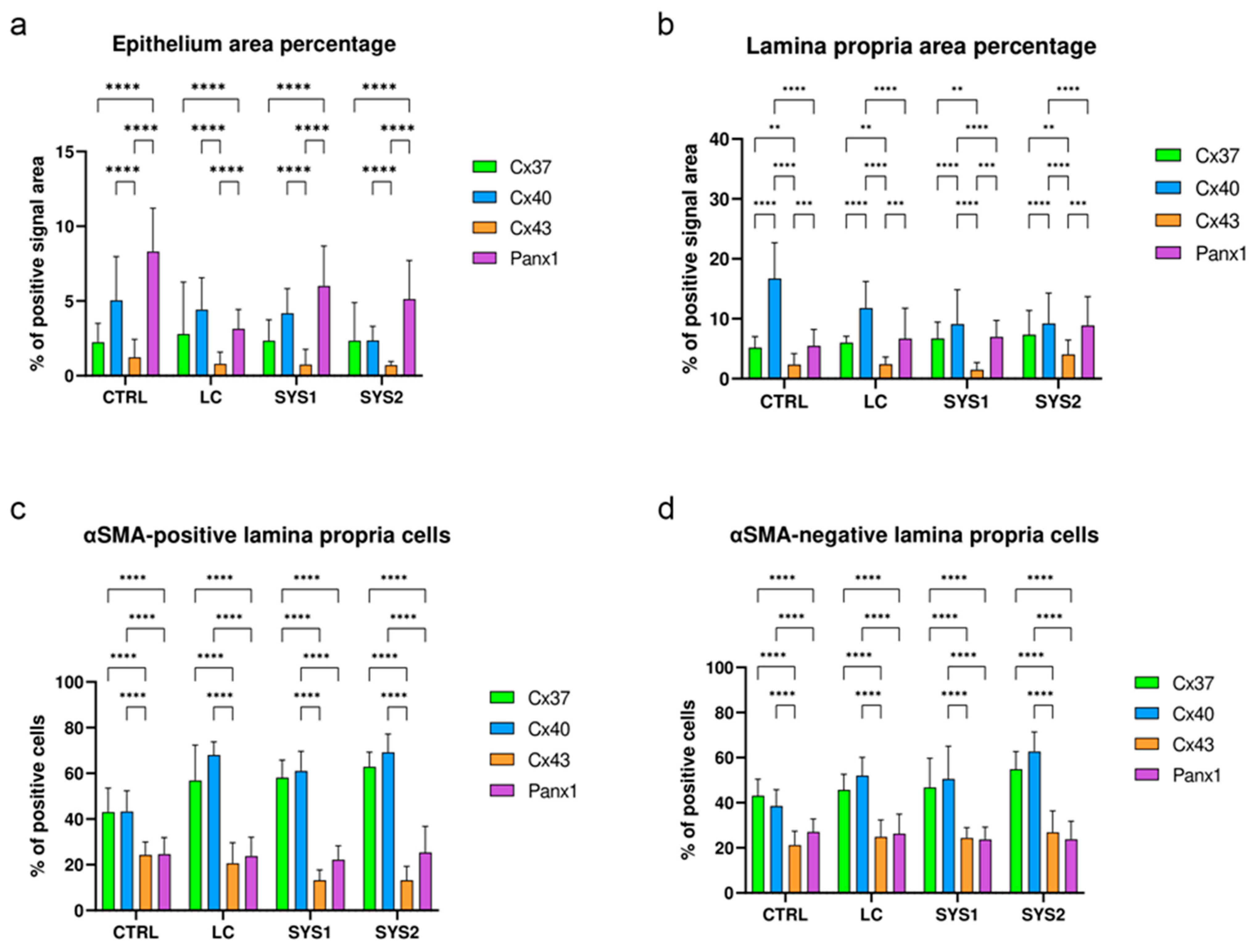
3.5. Comparison of Cx37, Cx40, Cx43, and Panx1 Protein Expressions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Furst, D.E.; Clements, P.J. Hypothesis for the pathogenesis of systemic sclerosis. J. Rheumatol. Suppl. 1997, 48, 53–57. [Google Scholar]
- LeRoy, E.C.; Black, C.; Fleischmajer, R.; Jablonska, S.; Krieg, T.; Medsger, T.A., Jr.; Rowell, N.; Wollheim, F. Scleroderma (systemic sclerosis): Classification, subsets and pathogenesis. J. Rheumatol. 1988, 15, 202–205. [Google Scholar] [PubMed]
- Chandran, G.; Smith, M.; Ahern, M.J.; Roberts-Thomson, P.J. A study of scleroderma in South Australia: Prevalence, subset characteristics and nailfold capillaroscopy. Aust. N. Z. J. Med. 1995, 25, 688–694. [Google Scholar] [CrossRef]
- Cockrill, T.; del Junco, D.J.; Arnett, F.C.; Assassi, S.; Tan, F.K.; McNearney, T.; Fischbach, M.; Perry, M.; Mayes, M.D. Separate influences of birth order and gravidity/parity on the development of systemic sclerosis. Arthritis Care Res. 2010, 62, 418–424. [Google Scholar] [CrossRef]
- Marie, I.; Gehanno, J.F.; Bubenheim, M.; Duval-Modeste, A.B.; Joly, P.; Dominique, S.; Bravard, P.; Noel, D.; Cailleux, A.F.; Weber, J.; et al. Prospective study to evaluate the association between systemic sclerosis and occupational exposure and review of the literature. Autoimmun. Rev. 2014, 13, 151–156. [Google Scholar] [CrossRef]
- Mayes, M.D.; Lacey, J.V., Jr.; Beebe-Dimmer, J.; Gillespie, B.W.; Cooper, B.; Laing, T.J.; Schottenfeld, D. Prevalence, incidence, survival, and disease characteristics of systemic sclerosis in a large US population. Arthritis Rheum. 2003, 48, 2246–2255. [Google Scholar] [CrossRef] [PubMed]
- Steen, V.D.; Medsger, T.A., Jr. Epidemiology and natural history of systemic sclerosis. Rheum. Dis. Clin. N. Am. 1990, 16, 1–10. [Google Scholar] [CrossRef]
- Thoua, N.M.; Bunce, C.; Brough, G.; Forbes, A.; Emmanuel, A.V.; Denton, C.P. Assessment of gastrointestinal symptoms in patients with systemic sclerosis in a UK tertiary referral centre. Rheumatology 2010, 49, 1770–1775. [Google Scholar] [CrossRef]
- Sallam, H.; McNearney, T.A.; Chen, J.D. Systematic review: Pathophysiology and management of gastrointestinal dysmotility in systemic sclerosis (scleroderma). Aliment. Pharmacol. Ther. 2006, 23, 691–712. [Google Scholar] [CrossRef]
- Forbes, A.; Marie, I. Gastrointestinal complications: The most frequent internal complications of systemic sclerosis. Rheumatology 2009, 48 (Suppl. S3), iii36–iii39. [Google Scholar] [CrossRef]
- LeRoy, E.C. A brief overview of the pathogenesis of scleroderma (systemic sclerosis). Ann. Rheum. Dis. 1992, 51, 286–288. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A. Cutaneous tissue repair: Basic biologic considerations. I. J. Am. Acad. Dermatol. 1985, 13, 701–725. [Google Scholar] [CrossRef]
- Border, W.A.; Ruoslahti, E. Transforming growth factor-beta in disease: The dark side of tissue repair. J. Clin. Investig. 1992, 90, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ihn, H.; Sato, S.; Fujimoto, M.; Kikuchi, K.; Kadono, T.; Tamaki, K.; Takehara, K. Circulating intercellular adhesion molecule-1 in the sera of patients with systemic sclerosis: Enhancement by inflammatory cytokines. Br. J. Rheumatol. 1997, 36, 1270–1275. [Google Scholar] [CrossRef]
- Jelaska, A.; Arakawa, M.; Broketa, G.; Korn, J.H. Heterogeneity of collagen synthesis in normal and systemic sclerosis skin fibroblasts. Increased proportion of high collagen-producing cells in systemic sclerosis fibroblasts. Arthritis Rheum. 1996, 39, 1338–1346. [Google Scholar] [CrossRef]
- Kirk, T.Z.; Mark, M.E.; Chua, C.C.; Chua, B.H.; Mayes, M.D. Myofibroblasts from scleroderma skin synthesize elevated levels of collagen and tissue inhibitor of metalloproteinase (TIMP-1) with two forms of TIMP-1. J. Biol. Chem. 1995, 270, 3423–3428. [Google Scholar] [CrossRef] [PubMed]
- Leroy, E.C.; Smith, E.A.; Kahaleh, M.B.; Trojanowska, M.; Silver, R.M. A strategy for determining the pathogenesis of systemic sclerosis. Is transforming growth factor beta the answer? Arthritis Rheum. 1989, 32, 817–825. [Google Scholar] [CrossRef]
- Kulozik, M.; Hogg, A.; Lankat-Buttgereit, B.; Krieg, T. Co-localization of transforming growth factor beta 2 with alpha 1(I) procollagen mRNA in tissue sections of patients with systemic sclerosis. J. Clin. Investig. 1990, 86, 917–922. [Google Scholar] [CrossRef]
- Shestopalov, V.I.; Panchin, Y. Pannexins and gap junction protein diversity. Cell. Mol. Life Sci. CMLS 2008, 65, 376–394. [Google Scholar] [CrossRef]
- Harris, A.L. Emerging issues of connexin channels: Biophysics fills the gap. Q. Rev. Biophys. 2001, 34, 325–472. [Google Scholar] [CrossRef]
- Penuela, S.; Bhalla, R.; Gong, X.Q.; Cowan, K.N.; Celetti, S.J.; Cowan, B.J.; Bai, D.; Shao, Q.; Laird, D.W. Pannexin 1 and pannexin 3 are glycoproteins that exhibit many distinct characteristics from the connexin family of gap junction proteins. J. Cell Sci. 2007, 120, 3772–3783. [Google Scholar] [CrossRef]
- Scemes, E.; Spray, D.C.; Meda, P. Connexins, pannexins, innexins: Novel roles of “hemi-channels”. Pflug. Arch. Eur. J. Physiol. 2009, 457, 1207–1226. [Google Scholar] [CrossRef] [PubMed]
- World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [CrossRef]
- van den Hoogen, F.; Khanna, D.; Fransen, J.; Johnson, S.R.; Baron, M.; Tyndall, A.; Matucci-Cerinic, M.; Naden, R.P.; Medsger, T.A., Jr.; Carreira, P.E.; et al. 2013 classification criteria for systemic sclerosis: An American college of rheumatology/European league against rheumatism collaborative initiative. Ann. Rheum. Dis. 2013, 72, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Khanna, D.; Furst, D.E.; Clements, P.J.; Allanore, Y.; Baron, M.; Czirjak, L.; Distler, O.; Foeldvari, I.; Kuwana, M.; Matucci-Cerinic, M.; et al. Standardization of the modified Rodnan skin score for use in clinical trials of systemic sclerosis. J. Scleroderma Relat. Disord. 2017, 2, 11–18. [Google Scholar] [CrossRef]
- Medsger, T.A., Jr.; Bombardieri, S.; Czirjak, L.; Scorza, R.; Della Rossa, A.; Bencivelli, W. Assessment of disease severity and prognosis. Clin. Exp. Rheumatol. 2003, 21, S42–S46. [Google Scholar]
- Boric, K.; Mardesic, S.; Martinovic Kaliterna, D.; Radic, M.; Tadin Hadjina, I.; Vukojevic, K.; Kosovic, I.; Solic, I.; Zekic Tomas, S.; Saraga-Babic, M. Expression of apoptotic and proliferation factors in gastric mucosa of patients with systemic sclerosis correlates with form of the disease. Sci. Rep. 2019, 9, 18461. [Google Scholar] [CrossRef] [PubMed]
- Ogorevc, M.; Kosovic, I.; Filipovic, N.; Bocina, I.; Juric, M.; Benzon, B.; Mardesic, S.; Vukojevic, K.; Saraga, M.; Kablar, B.; et al. Differences in Immunohistochemical and Ultrastructural Features between Podocytes and Parietal Epithelial Cells (PECs) Are Observed in Developing, Healthy Postnatal, and Pathologically Changed Human Kidneys. Int. J. Mol. Sci. 2022, 23, 7501. [Google Scholar] [CrossRef]
- Manetti, M.; Neumann, E.; Milia, A.F.; Tarner, I.H.; Bechi, P.; Matucci-Cerinic, M.; Ibba-Manneschi, L.; Muller-Ladner, U. Severe fibrosis and increased expression of fibrogenic cytokines in the gastric wall of systemic sclerosis patients. Arthritis Rheum. 2007, 56, 3442–3447. [Google Scholar] [CrossRef]
- Manetti, M.; Neumann, E.; Muller, A.; Schmeiser, T.; Saar, P.; Milia, A.F.; Endlicher, E.; Roeb, E.; Messerini, L.; Matucci-Cerinic, M.; et al. Endothelial/lymphocyte activation leads to prominent CD4+ T cell infiltration in the gastric mucosa of patients with systemic sclerosis. Arthritis Rheum. 2008, 58, 2866–2873. [Google Scholar] [CrossRef]
- Taroni, J.N.; Martyanov, V.; Huang, C.C.; Mahoney, J.M.; Hirano, I.; Shetuni, B.; Yang, G.Y.; Brenner, D.; Jung, B.; Wood, T.A.; et al. Molecular characterization of systemic sclerosis esophageal pathology identifies inflammatory and proliferative signatures. Arthritis Res. Ther. 2015, 17, 194. [Google Scholar] [CrossRef] [PubMed]
- Thoua, N.M.; Derrett-Smith, E.C.; Khan, K.; Dooley, A.; Shi-Wen, X.; Denton, C.P. Gut fibrosis with altered colonic contractility in a mouse model of scleroderma. Rheumatology 2012, 51, 1989–1998. [Google Scholar] [CrossRef]
- Maes, M.; Cogliati, B.; Crespo Yanguas, S.; Willebrords, J.; Vinken, M. Roles of connexins and pannexins in digestive homeostasis. Cell. Mol. Life Sci. CMLS 2015, 72, 2809–2821. [Google Scholar] [CrossRef]
- Nishitani, A.; Hirota, S.; Nishida, T.; Isozaki, K.; Hashimoto, K.; Nakagomi, N.; Matsuda, H. Differential expression of connexin 43 in gastrointestinal stromal tumours of gastric and small intestinal origin. J. Pathol. 2005, 206, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Koval, M.; Billaud, M.; Straub, A.C.; Johnstone, S.R.; Zarbock, A.; Duling, B.R.; Isakson, B.E. Spontaneous lung dysfunction and fibrosis in mice lacking connexin 40 and endothelial cell connexin 43. Am. J. Pathol. 2011, 178, 2536–2546. [Google Scholar] [CrossRef]
- Cogliati, B.; Mennecier, G.; Willebrords, J.; Da Silva, T.C.; Maes, M.; Pereira, I.V.A.; Crespo-Yanguas, S.; Hernandez-Blazquez, F.J.; Dagli, M.L.Z.; Vinken, M. Connexins, Pannexins, and Their Channels in Fibroproliferative Diseases. J. Membr. Biol. 2016, 249, 199–213. [Google Scholar] [CrossRef]
- Lu, F.; Gao, J.; Ogawa, R.; Hyakusoku, H. Variations in gap junctional intercellular communication and connexin expression in fibroblasts derived from keloid and hypertrophic scars. Plast. Reconstr. Surg. 2007, 119, 844–851. [Google Scholar] [CrossRef]
- Asazuma-Nakamura, Y.; Dai, P.; Harada, Y.; Jiang, Y.; Hamaoka, K.; Takamatsu, T. Cx43 contributes to TGF-beta signaling to regulate differentiation of cardiac fibroblasts into myofibroblasts. Exp. Cell Res. 2009, 315, 1190–1199. [Google Scholar] [CrossRef]
- Dolmatova, E.; Spagnol, G.; Boassa, D.; Baum, J.R.; Keith, K.; Ambrosi, C.; Kontaridis, M.I.; Sorgen, P.L.; Sosinsky, G.E.; Duffy, H.S. Cardiomyocyte ATP release through pannexin 1 aids in early fibroblast activation. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H1208–H1218. [Google Scholar] [CrossRef]
- Chekeni, F.B.; Elliott, M.R.; Sandilos, J.K.; Walk, S.F.; Kinchen, J.M.; Lazarowski, E.R.; Armstrong, A.J.; Penuela, S.; Laird, D.W.; Salvesen, G.S.; et al. Pannexin 1 channels mediate ’find-me’ signal release and membrane permeability during apoptosis. Nature 2010, 467, 863–867. [Google Scholar] [CrossRef]
- Qu, Y.; Misaghi, S.; Newton, K.; Gilmour, L.L.; Louie, S.; Cupp, J.E.; Dubyak, G.R.; Hackos, D.; Dixit, V.M. Pannexin-1 is required for ATP release during apoptosis but not for inflammasome activation. J. Immunol. 2011, 186, 6553–6561. [Google Scholar] [CrossRef] [PubMed]
- Stellato, M.; Dewenter, M.; Rudnik, M.; Hukara, A.; Ozsoy, C.; Renoux, F.; Pachera, E.; Gantenbein, F.; Seebeck, P.; Uhtjaerv, S.; et al. The AP-1 transcription factor Fosl-2 drives cardiac fibrosis and arrhythmias under immunofibrotic conditions. Commun. Biol. 2023, 6, 161. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.F.; Daniel, E.E. Gap junctions in gastrointestinal muscle contain multiple connexins. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G533–G543. [Google Scholar] [CrossRef]
- Seki, K.; Komuro, T. Distribution of interstitial cells of Cajal and gap junction protein, Cx 43 in the stomach of wild-type and W/Wv mutant mice. Anat. Embryol. 2002, 206, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Manetti, M.; Milia, A.F.; Benelli, G.; Messerini, L.; Matucci-Cerinic, M.; Ibba-Manneschi, L. The gastric wall in systemic sclerosis patients: A morphological study. Ital. J. Anat. Embryol. 2010, 115, 115–121. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | SSc Patients (n = 15) | Healthy Volunteers (n = 5) |
|---|---|---|
| Age (years) a | 54.3 ± 13.6 | 54.9 ± 11.2 |
| Female sex | 15 (100%) | 5 (100%) |
| Caucasian race | 15 (100%) | 5 (100%) |
| Disease duration b | 13 (1–35) | N/A |
| Antinuclear antibody positivity | 13 (86.7%) | 0 (0%) |
| Anti-centromere antibody positivity | 4 (26.7%) | 0 (0%) |
| Anti-topoisomerase I (Scl 70) antibody positivity | 9 (60%) | 0 (0%) |
| Modified Rodnan skin score b | 15 (4–26) | 0 (0–0) |
| Diffusing capacity for carbon monoxide (DLCO) a | 58.4 ± 22.2 | N/A |
| Presence of interstitial lung disease | 8 (53.3%) | 0 (0%) |
| Presence of gastrointestinal tract lesions | 10 (66.7%) | 0 (0%) |
| Presence of heart involvement | 3 (20%) | 0 (0%) |
| Presence of peripheral vascular involvement | 12 (80%) | 0 (0%) |
| Presence of digital ulcers | 3 (20%) | 0 (0%) |
| Presence of skeletal muscle involvement | 10 (66%) | 0 (0%) |
| Presence of joint and tendon involvement | 12 (80%) | 0 (0%) |
| Antibodies | Host | Code No. | Dilution | Source | |
|---|---|---|---|---|---|
| Primary | Anti-Cx37/GJA4 | Rabbit | ab181701 | 1:300 | Abcam (Cambridge, UK) |
| Anti-Cx40/GJA5 | Rabbit | ab213688 | 1:50 | Abcam (Cambridge, UK) | |
| Anti-Cx43/GJA1 | Goat | ab87645 | 1:100 | Abcam (Cambridge, UK) | |
| Anti-pannexin 1 | Rabbit | ABN242 | 1:300 | Merck KGaA (Darmstadt, Germany) | |
| Anit-Smooth Muscle Actin | Mouse | M0851 | 1:300 | Dako (Glostrup, Denmark) | |
| Secondary | Alexa Fluor® 488 AffiniPure Anti-Goat IgG (H + L) | Donkey | 705-545-003 | 1:400 | Jackson Immuno Research Laboratories, Inc., (Baltimore, PA, USA) |
| Alexa Fluor® 488 AffiniPure Anti-Rabbit IgG (H + L) | Donkey | 711-545-152 | 1:400 | Jackson Immuno Research Laboratories, Inc., (Baltimore, PA, USA) | |
| Rhodamine Red™-X (RRX) AffiniPure Anti-Mouse IgG (H + L) | Donkey | 715-295-151 | 1:400 | Jackson Immuno Research Laboratories, Inc., (Baltimore, PA, USA) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavic, B.; Ogorevc, M.; Boric, K.; Vukovic, D.; Saraga-Babic, M.; Mardesic, S. Connexin 37, 40, 43 and Pannexin 1 Expression in the Gastric Mucosa of Patients with Systemic Sclerosis. Biomedicines 2023, 11, 2487. https://doi.org/10.3390/biomedicines11092487
Pavic B, Ogorevc M, Boric K, Vukovic D, Saraga-Babic M, Mardesic S. Connexin 37, 40, 43 and Pannexin 1 Expression in the Gastric Mucosa of Patients with Systemic Sclerosis. Biomedicines. 2023; 11(9):2487. https://doi.org/10.3390/biomedicines11092487
Chicago/Turabian StylePavic, Berna, Marin Ogorevc, Katarina Boric, Dubravka Vukovic, Mirna Saraga-Babic, and Snjezana Mardesic. 2023. "Connexin 37, 40, 43 and Pannexin 1 Expression in the Gastric Mucosa of Patients with Systemic Sclerosis" Biomedicines 11, no. 9: 2487. https://doi.org/10.3390/biomedicines11092487