
Indoleamine 2,3-Dioxygenase (IDO) Activity: A Perspective Biomarker for Laboratory Determination in Tumor Immunotherapy
Abstract
:1. Introduction
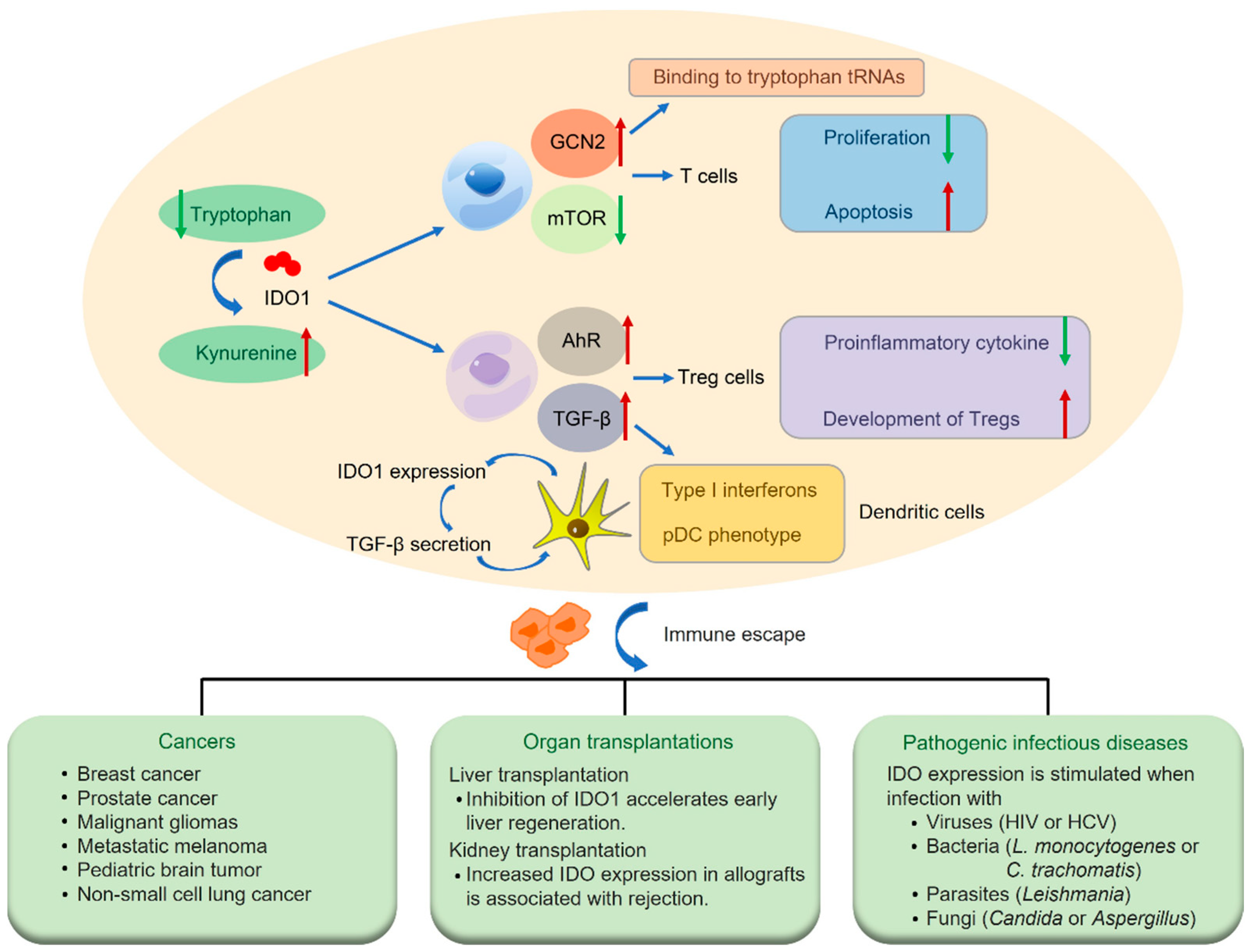
2. Immune Regulation of IDO Pathway
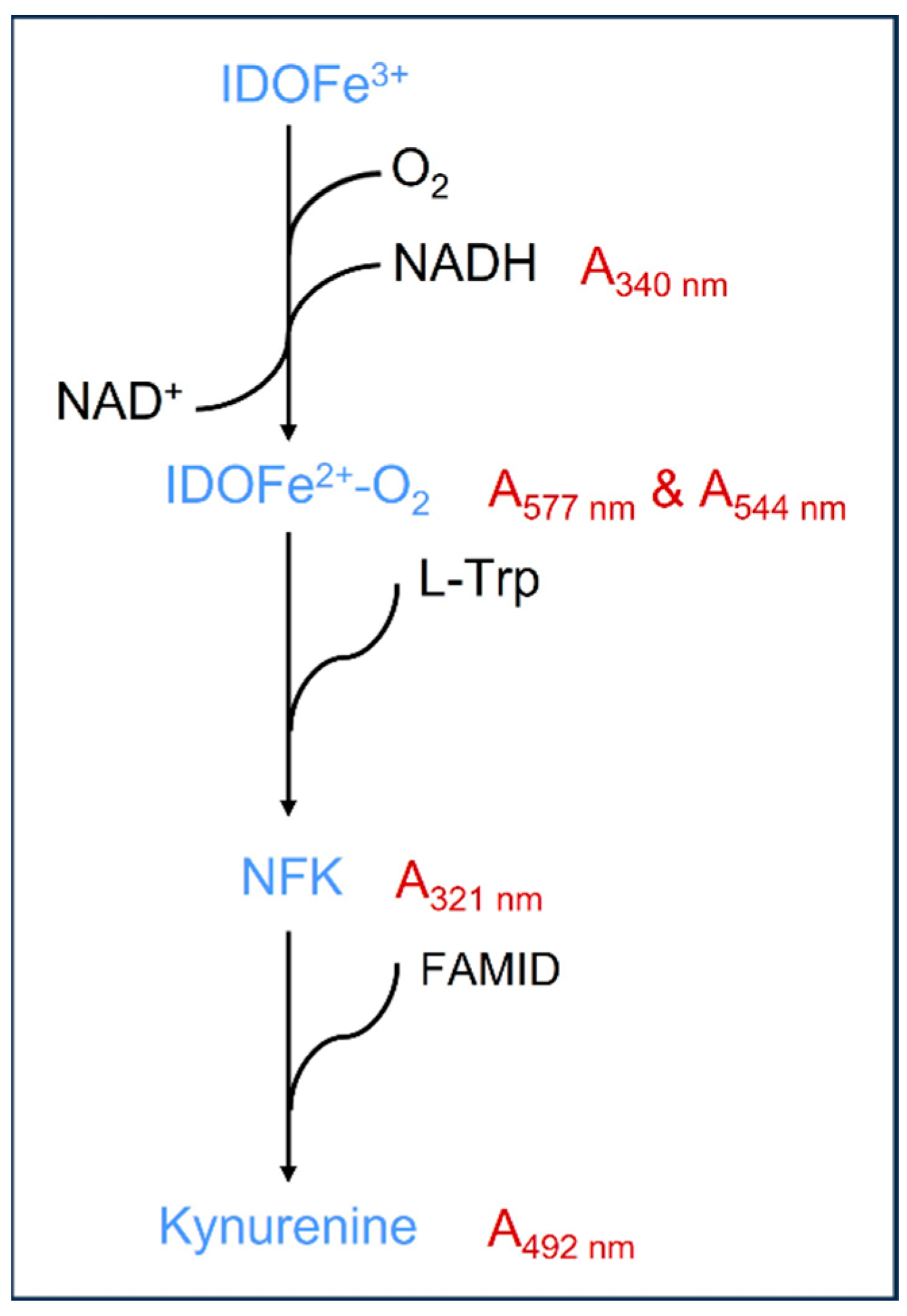
3. Biochemical Properties of IDO in the KP
4. A Perspective Biomarker in a Variety of Human Diseases
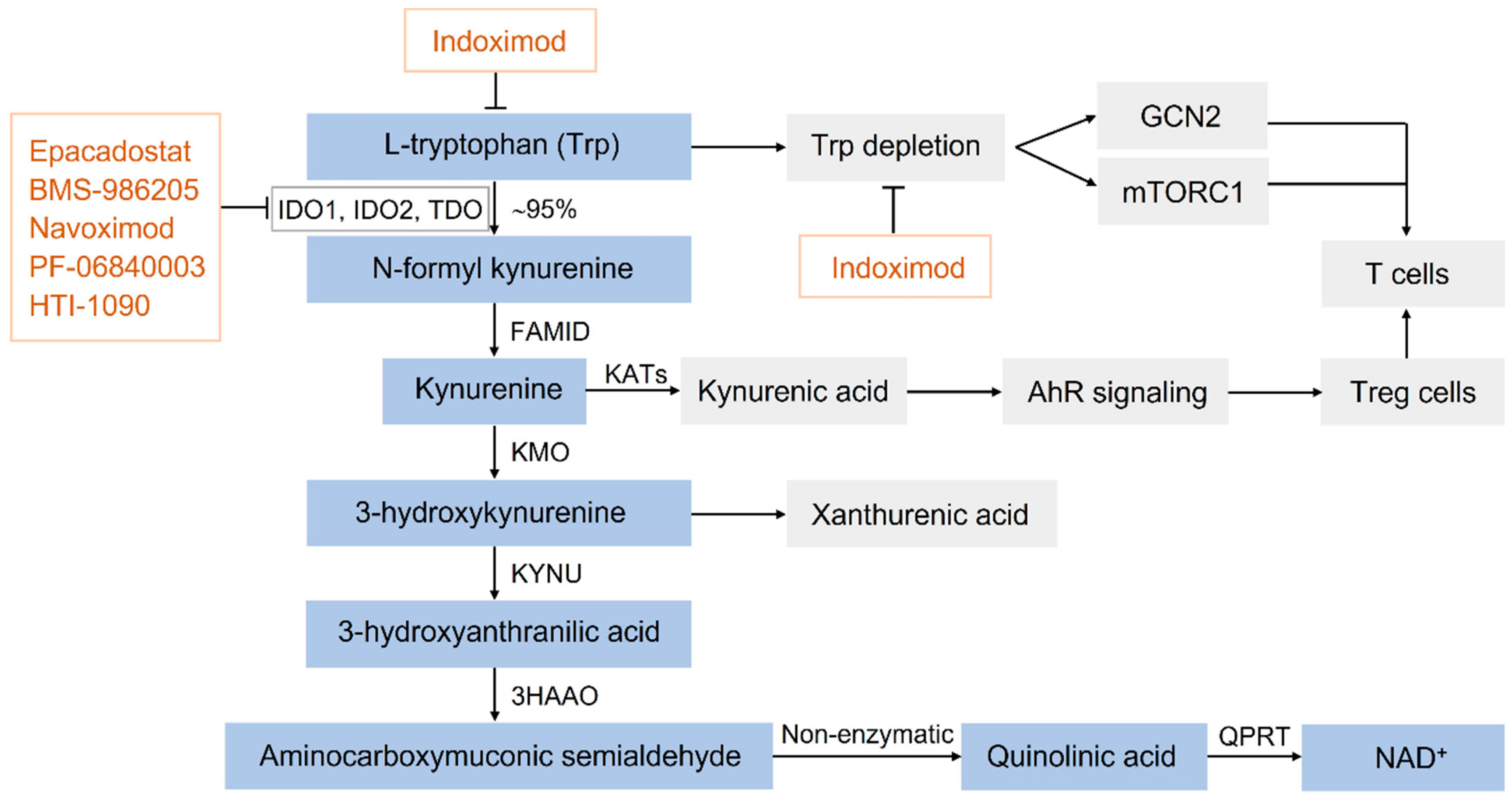
5. IDO Inhibitors in Clinical Trials for Cancer Immunotherapy
6. Applications of IDO Activity Assays
6.1. IDO Enzyme Activity Assays Based on Different Intermediates
6.2. IDO Activity Assays in Inhibitors In Vitro
6.3. IDO Activity Assays in Human Plasma/Serum
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barik, S. The Uniqueness of Tryptophan in Biology: Properties, Metabolism, Interactions and Localization in Proteins. Int. J. Mol. Sci. 2020, 21, 8776. [Google Scholar] [CrossRef]
- Badawy, A.A. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. IJTR 2017, 10, 1178646917691938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badawy, A.A. Kynurenine pathway and human systems. Exp. Gerontol. 2020, 129, 110770. [Google Scholar] [CrossRef]
- González Esquivel, D.; Ramírez-Ortega, D.; Pineda, B.; Castro, N.; Ríos, C.; Pérez de la Cruz, V. Kynurenine pathway metabolites and enzymes involved in redox reactions. Neuropharmacology 2017, 112, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.S.; Gupta, S.; Barton, S.A.; Vander Wall, R.; Tan, V.; Heng, B.; Phillips, J.K.; Guillemin, G.J. Temporal Profile of Kynurenine Pathway Metabolites in a Rodent Model of Autosomal Recessive Polycystic Kidney Disease. Int. J. Tryptophan Res. IJTR 2022, 15, 11786469221126063. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Ramprasath, T.; Wang, H.; Zou, M.H. Abnormal kynurenine pathway of tryptophan catabolism in cardiovascular diseases. Cell. Mol. Life Sci. CMLS 2017, 74, 2899–2916. [Google Scholar] [CrossRef] [PubMed]
- Platten, M.; Nollen, E.A.A.; Röhrig, U.F.; Fallarino, F.; Opitz, C.A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 2019, 18, 379–401. [Google Scholar] [CrossRef]
- Too, L.K.; Li, K.M.; Suarna, C.; Maghzal, G.J.; Stocker, R.; McGregor, I.S.; Hunt, N.H. Deletion of TDO2, IDO-1 and IDO-2 differentially affects mouse behavior and cognitive function. Behav. Brain Res. 2016, 312, 102–117. [Google Scholar] [CrossRef]
- Peyraud, F.; Guegan, J.P.; Bodet, D.; Cousin, S.; Bessede, A.; Italiano, A. Targeting Tryptophan Catabolism in Cancer Immunotherapy Era: Challenges and Perspectives. Front. Immunol. 2022, 13, 807271. [Google Scholar] [CrossRef]
- Dostal, C.R.; Carson Sulzer, M.; Kelley, K.W.; Freund, G.G.; McCusker, R.H. Glial and tissue-specific regulation of Kynurenine Pathway dioxygenases by acute stress of mice. Neurobiol. Stress 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Bakmiwewa, S.M.; Fatokun, A.A.; Tran, A.; Payne, R.J.; Hunt, N.H.; Ball, H.J. Identification of selective inhibitors of indoleamine 2,3-dioxygenase 2. Bioorganic Med. Chem. Lett. 2012, 22, 7641–7646. [Google Scholar] [CrossRef]
- Théate, I.; van Baren, N.; Pilotte, L.; Moulin, P.; Larrieu, P.; Renauld, J.C.; Hervé, C.; Gutierrez-Roelens, I.; Marbaix, E.; Sempoux, C.; et al. Extensive profiling of the expression of the indoleamine 2,3-dioxygenase 1 protein in normal and tumoral human tissues. Cancer Immunol. Res. 2015, 3, 161–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvajal-Hausdorf, D.E.; Mani, N.; Velcheti, V.; Schalper, K.A.; Rimm, D.L. Objective measurement and clinical significance of IDO1 protein in hormone receptor-positive breast cancer. J. Immunother. Cancer 2017, 5, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlo, L.M.; Mandik-Nayak, L. IDO2: A Pathogenic Mediator of Inflammatory Autoimmunity. Clin. Med. Insights. Pathol. 2016, 9, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlova, A.; Frédérick, R. Current state on tryptophan 2,3-dioxygenase inhibitors: A patent review. Expert. Opin. Ther. Pat. 2019, 29, 11–23. [Google Scholar] [CrossRef]
- Meininger, D.; Zalameda, L.; Liu, Y.; Stepan, L.P.; Borges, L.; McCarter, J.D.; Sutherland, C.L. Purification and kinetic characterization of human indoleamine 2,3-dioxygenases 1 and 2 (IDO1 and IDO2) and discovery of selective IDO1 inhibitors. Biochim. Et. Biophys. Acta 2011, 1814, 1947–1954. [Google Scholar] [CrossRef]
- Takkenkamp, T.J.; Jalving, M.; Hoogwater, F.J.H.; Walenkamp, A.M.E. The immune tumour microenvironment of neuroendocrine tumours and its implications for immune checkpoint inhibitors. Endocr. Relat. Cancer 2020, 27, R329–R343. [Google Scholar] [CrossRef]
- Botticelli, A.; Cerbelli, B.; Lionetto, L.; Zizzari, I.; Salati, M.; Pisano, A.; Federica, M.; Simmaco, M.; Nuti, M.; Marchetti, P. Can IDO activity predict primary resistance to anti-PD-1 treatment in NSCLC? J. Transl. Med. 2018, 16, 219. [Google Scholar] [CrossRef] [Green Version]
- Botticelli, A.; Mezi, S.; Pomati, G.; Cerbelli, B.; Cerbelli, E.; Roberto, M.; Giusti, R.; Cortellini, A.; Lionetto, L.; Scagnoli, S.; et al. Tryptophan Catabolism as Immune Mechanism of Primary Resistance to Anti-PD-1. Front. Immunol. 2020, 11, 1243. [Google Scholar] [CrossRef]
- Wu, C.; Spector, S.A.; Theodoropoulos, G.; Nguyen, D.J.M.; Kim, E.Y.; Garcia, A.; Savaraj, N.; Lim, D.C.; Paul, A.; Feun, L.G.; et al. Dual inhibition of IDO1/TDO2 enhances anti-tumor immunity in platinum-resistant non-small cell lung cancer. Cancer Metab. 2023, 11, 7. [Google Scholar] [CrossRef]
- Mandarano, M.; Orecchini, E.; Bellezza, G.; Vannucci, J.; Ludovini, V.; Baglivo, S.; Tofanetti, F.R.; Chiari, R.; Loreti, E.; Puma, F.; et al. Kynurenine/Tryptophan Ratio as a Potential Blood-Based Biomarker in Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2021, 22, 4403. [Google Scholar] [CrossRef] [PubMed]
- Takikawa, O. Biochemical and medical aspects of the indoleamine 2,3-dioxygenase-initiated L-tryptophan metabolism. Biochem. Biophys. Res. Commun. 2005, 338, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Lancellotti, S.; Novarese, L.; De Cristofaro, R. Biochemical properties of indoleamine 2,3-dioxygenase: From structure to optimized design of inhibitors. Curr. Med. Chem. 2011, 18, 2205–2214. [Google Scholar] [CrossRef]
- Najfeld, V.; Menninger, J.; Muhleman, D.; Comings, D.E.; Gupta, S.L. Localization of indoleamine 2,3-dioxygenase gene (INDO) to chromosome 8p12-->p11 by fluorescent in situ hybridization. Cytogenet. Cell Genet. 1993, 64, 231–232. [Google Scholar] [CrossRef] [PubMed]
- Puccetti, P.; Grohmann, U. IDO and regulatory T cells: A role for reverse signalling and non-canonical NF-kappaB activation. Nat. Rev. Immunol. 2007, 7, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Azimnasab-Sorkhabi, P.; Soltani-Asl, M.; Yoshinaga, T.T.; Zaidan Dagli, M.L.; Massoco, C.O.; Kfoury Junior, J.R. Indoleamine-2,3 dioxygenase: A fate-changer of the tumor microenvironment. Mol. Biol. Rep. 2023, 50, 6133–6145. [Google Scholar] [CrossRef]
- Munn, D.H.; Mellor, A.L. IDO in the Tumor Microenvironment: Inflammation, Counter-Regulation, and Tolerance. Trends Immunol. 2016, 37, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Liu, D.; Song, P.; Zou, M.H. Tryptophan-kynurenine pathway is dysregulated in inflammation, and immune activation. Front. Biosci. (Landmark Ed.) 2015, 20, 1116–1143. [Google Scholar] [CrossRef] [Green Version]
- Heitger, A. Regulation of expression and function of IDO in human dendritic cells. Curr. Med. Chem. 2011, 18, 2222–2233. [Google Scholar] [CrossRef]
- Stone, T.W.; Williams, R.O. Modulation of T cells by tryptophan metabolites in the kynurenine pathway. Trends Pharmacol. Sci. 2023, 44, 442–456. [Google Scholar] [CrossRef]
- Wu, H.; Gong, J.; Liu, Y. Indoleamine 2, 3-dioxygenase regulation of immune response (Review). Mol. Med. Rep. 2018, 17, 4867–4873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, A. Role of indoleamine 2,3-dioxygenase 1 (IDO1) and kynurenine pathway in the regulation of the aging process. Ageing Res. Rev. 2022, 75, 101573. [Google Scholar] [CrossRef]
- Zhai, L.; Bell, A.; Ladomersky, E.; Lauing, K.L.; Bollu, L.; Sosman, J.A.; Zhang, B.; Wu, J.D.; Miller, S.D.; Meeks, J.J.; et al. Immunosuppressive IDO in Cancer: Mechanisms of Action, Animal Models, and Targeting Strategies. Front. Immunol. 2020, 11, 1185. [Google Scholar] [CrossRef] [PubMed]
- Löb, S.; Königsrainer, A. Role of IDO in organ transplantation: Promises and difficulties. Int. Rev. Immunol. 2009, 28, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Hainz, U.; Jürgens, B.; Heitger, A. The role of indoleamine 2,3-dioxygenase in transplantation. Transpl. Int. Off. J. Eur. Soc. Organ. Transplant. 2007, 20, 118–127. [Google Scholar] [CrossRef]
- Barth, H.; Raghuraman, S. Persistent infectious diseases say—IDO. Role of indoleamine-2,3-dioxygenase in disease pathogenesis and implications for therapy. Crit. Rev. Microbiol. 2014, 40, 360–368. [Google Scholar] [CrossRef]
- Suzuki, T.; Kawamichi, H.; Imai, K. A myoglobin evolved from indoleamine 2,3-dioxygenase, a tryptophan-degrading enzyme. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1998, 121, 117–128. [Google Scholar] [CrossRef]
- Suzuki, T.; Imai, K. Evolution of myoglobin. Cell. Mol. Life Sci. CMLS 1998, 54, 979–1004. [Google Scholar] [CrossRef]
- Suzuki, T.; Yokouchi, K.; Kawamichi, H.; Yamamoto, Y.; Uda, K.; Yuasa, H.J. Comparison of the sequences of Turbo and Sulculus indoleamine dioxygenase-like myoglobin genes. Gene 2003, 308, 89–94. [Google Scholar] [CrossRef]
- Hirata, F.; Ohnishi, T.; Hayaishi, O. Indoleamine 2,3-dioxygenase. Characterization and properties of enzyme. O2- complex. J. Biol. Chem. 1977, 252, 4637–4642. [Google Scholar] [CrossRef]
- Sono, M. Spectroscopic and equilibrium properties of the indoleamine 2,3-dioxygenase-tryptophan-O2 ternary complex and of analogous enzyme derivatives. Tryptophan binding to ferrous enzyme adducts with dioxygen, nitric oxide, and carbon monoxide. Biochemistry 1986, 25, 6089–6097. [Google Scholar] [CrossRef]
- Rosell, F.I.; Kuo, H.H.; Mauk, A.G. NADH oxidase activity of indoleamine 2,3-dioxygenase. J. Biol. Chem. 2011, 286, 29273–29283. [Google Scholar] [CrossRef] [Green Version]
- Grant, R.S. Indoleamine 2,3-Dioxygenase Activity Increases NAD+ Production in IFN-γ-Stimulated Human Primary Mononuclear Cells. Int. J. Tryptophan Res. IJTR 2018, 11, 1178646917751636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehraj, V.; Routy, J.P. Tryptophan Catabolism in Chronic Viral Infections: Handling Uninvited Guests. Int. J. Tryptophan Res. IJTR 2015, 8, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Mbongue, J.C.; Nicholas, D.A.; Torrez, T.W.; Kim, N.S.; Firek, A.F.; Langridge, W.H. The Role of Indoleamine 2, 3-Dioxygenase in Immune Suppression and Autoimmunity. Vaccines 2015, 3, 703–729. [Google Scholar] [CrossRef] [Green Version]
- Myint, A.M.; Kim, Y.K. Network beyond IDO in psychiatric disorders: Revisiting neurodegeneration hypothesis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 48, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiang, T.; Yang, Y.; Deng, W.; Lu, H.; Wang, S.; Liu, R.; Chang, M.; Wu, S.; Gao, Y.; et al. Postpartum hepatitis and host immunity in pregnant women with chronic HBV infection. Front. Immunol. 2022, 13, 1112234. [Google Scholar] [CrossRef] [PubMed]
- Lashgari, N.A.; Roudsari, N.M.; Shayan, M.; Niazi Shahraki, F.; Hosseini, Y.; Momtaz, S.; Abdolghaffari, A.H. IDO/Kynurenine; novel insight for treatment of inflammatory diseases. Cytokine 2023, 166, 156206. [Google Scholar] [CrossRef]
- Pan, B.; Zhang, F.; Sun, J.; Chen, D.; Huang, W.; Zhang, H.; Cao, C.; Wan, X. Correlation of Indoleamine-2,3-Dioxygenase and Chronic Kidney Disease: A Pilot Study. J. Immunol. Res. 2021, 2021, 8132569. [Google Scholar] [CrossRef]
- Torosyan, R.; Huang, S.; Bommi, P.V.; Tiwari, R.; An, S.Y.; Schonfeld, M.; Rajendran, G.; Kavanaugh, M.A.; Gibbs, B.; Truax, A.D.; et al. Hypoxic preconditioning protects against ischemic kidney injury through the IDO1/kynurenine pathway. Cell Rep. 2021, 36, 109547. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, K.; Shinde, R.; Liu, H.; Gnana-Prakasam, J.P.; Veeranan-Karmegam, R.; Huang, L.; Ravishankar, B.; Bradley, J.; Kvirkvelia, N.; McMenamin, M.; et al. Amino acid metabolism inhibits antibody-driven kidney injury by inducing autophagy. J. Immunol. 2015, 194, 5713–5724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, K.; Li, D.; Xu, F.; Guo, H.; Feng, F.; Ding, Y.; Wan, X.; Sun, N.; Zhang, Y.; Fan, J.; et al. IDO1 as a new immune biomarker for diabetic nephropathy and its correlation with immune cell infiltration. Int. Immunopharmacol. 2021, 94, 107446. [Google Scholar] [CrossRef]
- Zhou, Q.; Shi, Y.; Chen, C.; Wu, F.; Chen, Z. A narrative review of the roles of indoleamine 2,3-dioxygenase and tryptophan-2,3-dioxygenase in liver diseases. Ann. Transl. Med. 2021, 9, 174. [Google Scholar] [CrossRef]
- Xu, L.; Ling, J.; Su, C.; Su, Y.W.; Xu, Y.; Jiang, Z. Emerging Roles on Immunological Effect of Indoleamine 2,3-Dioxygenase in Liver Injuries. Front. Med. 2021, 8, 756435. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kubota, N.; Crouchet, E.; Koneru, B.; Marquez, C.A.; Jajoriya, A.K.; Panda, G.; Qian, T.; Zhu, S.; Goossens, N.; et al. Molecular signatures of long-term hepatocellular carcinoma risk in nonalcoholic fatty liver disease. Sci. Transl. Med. 2022, 14, eabo4474. [Google Scholar] [CrossRef] [PubMed]
- Yoshio, S.; Sugiyama, M.; Shoji, H.; Mano, Y.; Mita, E.; Okamoto, T.; Matsuura, Y.; Okuno, A.; Takikawa, O.; Mizokami, M.; et al. Indoleamine-2,3-dioxygenase as an effector and an indicator of protective immune responses in patients with acute hepatitis B. Hepatol. 2016, 63, 83–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepiller, Q.; Soulier, E.; Li, Q.; Lambotin, M.; Barths, J.; Fuchs, D.; Stoll-Keller, F.; Liang, T.J.; Barth, H. Antiviral and Immunoregulatory Effects of Indoleamine-2,3-Dioxygenase in Hepatitis C Virus Infection. J. Innate Immun. 2015, 7, 530–544. [Google Scholar] [CrossRef]
- Chunna, X.; Xiao, L.; Yu, T.; Lei, Z.; Xiufang, W. IDO expression in lung tissue and its relationship with IFN-γ in asthmatic mice. Chin. Pediatr. Integr. Tradit. West. Med. 2021, 13, 103–107. (In Chinese) [Google Scholar]
- Makala, L.H.; Baban, B.; Lemos, H.; El-Awady, A.R.; Chandler, P.R.; Hou, D.Y.; Munn, D.H.; Mellor, A.L. Leishmania major attenuates host immunity by stimulating local indoleamine 2,3-dioxygenase expression. J. Infect. Dis. 2011, 203, 715–725. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.J.; Ma, P.Y.; Liu, H.; Cao, J.P.; Li, H.M.; Zheng, M.H. Progress of researches on the involvement of indoleamine 2, 3-dioxygenase in regulation of parasite-host immune interactions. Chin. J. Schistosomiasis Control 2020, 33, 209–212. [Google Scholar] [CrossRef]
- Dai, W.; Pan, H.; Kwok, O.; Dubey, J.P. Human indoleamine 2,3-dioxygenase inhibits Toxoplasma gondii growth in fibroblast cells. J. Interferon Res. 1994, 14, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Raitala, A.; Karjalainen, J.; Oja, S.S.; Kosunen, T.U.; Hurme, M. Helicobacter pylori-induced indoleamine 2,3-dioxygenase activity in vivo is regulated by TGFB1 and CTLA4 polymorphisms. Mol. Immunol. 2007, 44, 1011–1014. [Google Scholar] [CrossRef] [PubMed]
- Dolšak, A.; Gobec, S.; Sova, M. Indoleamine and tryptophan 2,3-dioxygenases as important future therapeutic targets. Pharmacol. Ther. 2021, 221, 107746. [Google Scholar] [CrossRef]
- Le Naour, J.; Galluzzi, L.; Zitvogel, L.; Kroemer, G.; Vacchelli, E. Trial watch: IDO inhibitors in cancer therapy. Oncoimmunology 2020, 9, 1777625. [Google Scholar] [CrossRef]
- Brincks, E.L.; Adams, J.; Wang, L.; Turner, B.; Marcinowicz, A.; Ke, J.; Essmann, M.; Mautino, L.M.; Allen, C.V.; Kumar, S.; et al. Indoximod opposes the immunosuppressive effects mediated by IDO and TDO via modulation of AhR function and activation of mTORC1. Oncotarget 2020, 11, 2438–2461. [Google Scholar] [CrossRef] [PubMed]
- Metz, R.; Rust, S.; Duhadaway, J.B.; Mautino, M.R.; Munn, D.H.; Vahanian, N.N.; Link, C.J.; Prendergast, G.C. IDO inhibits a tryptophan sufficiency signal that stimulates mTOR: A novel IDO effector pathway targeted by D-1-methyl-tryptophan. Oncoimmunology 2012, 1, 1460–1468. [Google Scholar] [CrossRef] [Green Version]
- Soliman, H.H.; Minton, S.E.; Han, H.S.; Ismail-Khan, R.; Neuger, A.; Khambati, F.; Noyes, D.; Lush, R.; Chiappori, A.A.; Roberts, J.D.; et al. A phase I study of indoximod in patients with advanced malignancies. Oncotarget 2016, 7, 22928–22938. [Google Scholar] [CrossRef] [Green Version]
- Fox, E.; Oliver, T.; Rowe, M.; Thomas, S.; Zakharia, Y.; Gilman, P.B.; Muller, A.J.; Prendergast, G.C. Indoximod: An Immunometabolic Adjuvant That Empowers T Cell Activity in Cancer. Front. Oncol. 2018, 8, 370. [Google Scholar] [CrossRef] [Green Version]
- Labadie, B.W.; Bao, R.; Luke, J.J. Reimagining IDO Pathway Inhibition in Cancer Immunotherapy via Downstream Focus on the Tryptophan-Kynurenine-Aryl Hydrocarbon Axis. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019, 25, 1462–1471. [Google Scholar] [CrossRef] [Green Version]
- Yue, E.W.; Sparks, R.; Polam, P.; Modi, D.; Douty, B.; Wayland, B.; Glass, B.; Takvorian, A.; Glenn, J.; Zhu, W.; et al. INCB24360 (Epacadostat), a Highly Potent and Selective Indoleamine-2,3-dioxygenase 1 (IDO1) Inhibitor for Immuno-oncology. ACS Med. Chem. Lett. 2017, 8, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Odunsi, K.; Qian, F.; Lugade, A.A.; Yu, H.; Geller, M.A.; Fling, S.P.; Kaiser, J.C.; Lacroix, A.M.; D’Amico, L.; Ramchurren, N.; et al. Metabolic adaptation of ovarian tumors in patients treated with an IDO1 inhibitor constrains antitumor immune responses. Sci. Transl. Med. 2022, 14, eabg8402. [Google Scholar] [CrossRef] [PubMed]
- Long, G.V.; Dummer, R.; Hamid, O.; Gajewski, T.F.; Caglevic, C.; Dalle, S.; Arance, A.; Carlino, M.S.; Grob, J.J.; Kim, T.M.; et al. Epacadostat plus pembrolizumab versus placebo plus pembrolizumab in patients with unresectable or metastatic melanoma (ECHO-301/KEYNOTE-252): A phase 3, randomised, double-blind study. Lancet. Oncol. 2019, 20, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Cherney, E.C.; Zhang, L.; Nara, S.; Zhu, X.; Gullo-Brown, J.; Maley, D.; Lin, T.A.; Hunt, J.T.; Huang, C.; Yang, Z.; et al. Discovery and Preclinical Evaluation of BMS-986242, a Potent, Selective Inhibitor of Indoleamine-2,3-dioxygenase 1. ACS Med. Chem. Lett. 2021, 12, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Balog, A.; Lin, T.A.; Maley, D.; Gullo-Brown, J.; Kandoussi, E.H.; Zeng, J.; Hunt, J.T. Preclinical Characterization of Linrodostat Mesylate, a Novel, Potent, and Selective Oral Indoleamine 2,3-Dioxygenase 1 Inhibitor. Mol. Cancer Ther. 2021, 20, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Sonpavde, G.; Necchi, A.; Gupta, S.; Steinberg, G.D.; Gschwend, J.E.; Van Der Heijden, M.S.; Garzon, N.; Ibrahim, M.; Raybold, B.; Liaw, D.; et al. ENERGIZE: A Phase III study of neoadjuvant chemotherapy alone or with nivolumab with/without linrodostat mesylate for muscle-invasive bladder cancer. Future Oncol. 2020, 16, 4359–4368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Suchomel, J.; Yanez, E.; Yost, E.; Liang, X.; Zhu, R.; Le, H.; Siebers, N.; Joas, L.; Morley, R.; et al. Investigation of the absolute bioavailability and human mass balance of navoximod, a novel IDO1 inhibitor. Br. J. Clin. Pharmacol. 2019, 85, 1751–1760. [Google Scholar] [CrossRef]
- Cheong, J.E.; Ekkati, A.; Sun, L. A patent review of IDO1 inhibitors for cancer. Expert. Opin. Ther. Pat. 2018, 28, 317–330. [Google Scholar] [CrossRef]
- Kumar, S.; Waldo, J.P.; Jaipuri, F.A.; Marcinowicz, A.; Van Allen, C.; Adams, J.; Kesharwani, T.; Zhang, X.; Metz, R.; Oh, A.J.; et al. Discovery of Clinical Candidate (1R,4r)-4-((R)-2-((S)-6-Fluoro-5H-imidazo[5,1-a]isoindol-5-yl)-1-hydroxyethyl)cyclohexan-1-ol (Navoximod), a Potent and Selective Inhibitor of Indoleamine 2,3-Dioxygenase 1. J. Med. Chem. 2019, 62, 6705–6733. [Google Scholar] [CrossRef]
- Gomes, B.; Driessens, G.; Bartlett, D.; Cai, D.; Cauwenberghs, S.; Crosignani, S.; Dalvie, D.; Denies, S.; Dillon, C.P.; Fantin, V.R.; et al. Characterization of the Selective Indoleamine 2,3-Dioxygenase-1 (IDO1) Catalytic Inhibitor EOS200271/PF-06840003 Supports IDO1 as a Critical Resistance Mechanism to PD-(L)1 Blockade Therapy. Mol. Cancer Ther. 2018, 17, 2530–2542. [Google Scholar] [CrossRef] [Green Version]
- Reardon, D.A.; Desjardins, A.; Rixe, O.; Cloughesy, T.; Alekar, S.; Williams, J.H.; Li, R.; Taylor, C.T.; Lassman, A.B. A phase 1 study of PF-06840003, an oral indoleamine 2,3-dioxygenase 1 (IDO1) inhibitor in patients with recurrent malignant glioma. Investig. New Drugs 2020, 38, 1784–1795. [Google Scholar] [CrossRef]
- Muller, A.J.; Mondal, A.; Dey, S.; Prendergast, G.C. IDO1 and inflammatory neovascularization: Bringing new blood to tumor-promoting inflammation. Front. Oncol. 2023, 13, 1165298. [Google Scholar] [CrossRef] [PubMed]
- Fellendorf, F.T.; Bonkat, N.; Dalkner, N.; Schönthaler, E.M.D.; Manchia, M.; Fuchs, D.; Reininghaus, E.Z. Indoleamine 2,3-dioxygenase (IDO)-activity in Severe Psychiatric Disorders: A Systemic Review. Curr. Top. Med. Chem. 2022, 22, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Nomiyama, S.; Hirata, F.; Hayaishi, O. Indoleamine 2,3-dioxygenase. Purification and some properties. J. Biol. Chem. 1978, 253, 4700–4706. [Google Scholar] [CrossRef]
- Sono, M.; Taniguchi, T.; Watanabe, Y.; Hayaishi, O. Indoleamine 2,3-dioxygenase. Equilibrium studies of the tryptophan binding to the ferric, ferrous, and CO-bound enzymes. J. Biol. Chem. 1980, 255, 1339–1345. [Google Scholar] [CrossRef]
- Hara, T.; Yamakura, F.; Takikawa, O.; Hiramatsu, R.; Kawabe, T.; Isobe, K.; Nagase, F. Diazotization of kynurenine by acidified nitrite secreted from indoleamine 2,3-dioxygenase-expressing myeloid dendritic cells. J. Immunol. Methods 2008, 332, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.; Guillemin, G. The Plasma [Kynurenine]/[Tryptophan] Ratio and Indoleamine 2,3-Dioxygenase: Time for Appraisal. Int. J. Tryptophan Res. IJTR 2019, 12, 1178646919868978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerejo, M.; Andrade, G.; Roca, C.; Sousa, J.; Rodrigues, C.; Pinheiro, R.; Chatterjee, S.; Vieira, H.; Calado, P. A powerful yeast-based screening assay for the identification of inhibitors of indoleamine 2,3-dioxygenase. J. Biomol. Screen. 2012, 17, 1362–1371. [Google Scholar] [CrossRef] [Green Version]
- Lanser, L.; Kink, P.; Egger, E.M.; Willenbacher, W.; Fuchs, D.; Weiss, G.; Kurz, K. Inflammation-Induced Tryptophan Breakdown is Related With Anemia, Fatigue, and Depression in Cancer. Front. Immunol. 2020, 11, 249. [Google Scholar] [CrossRef] [Green Version]
- Andersen, M.H. Tumor microenvironment antigens. Semin. Immunopathol. 2023, 45, 253–264. [Google Scholar] [CrossRef]
- Bello, C.; Heinisch, P.P.; Mihalj, M.; Carrel, T.; Luedi, M.M. Indoleamine-2,3-Dioxygenase as a Perioperative Marker of the Immune System. Front. Physiol. 2021, 12, 766511. [Google Scholar] [CrossRef]
- Passarelli, A.; Pisano, C.; Cecere, S.C.; Di Napoli, M.; Rossetti, S.; Tambaro, R.; Ventriglia, J.; Gherardi, F.; Iannacone, E.; Venanzio, S.S.; et al. Targeting immunometabolism mediated by the IDO1 Pathway: A new mechanism of immune resistance in endometrial cancer. Front. Immunol. 2022, 13, 953115. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.H. The T-win® technology: Immune-modulating vaccines. Semin. Immunopathol. 2019, 41, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Ala, M. The footprint of kynurenine pathway in every cancer: A new target for chemotherapy. Eur. J. Pharmacol. 2021, 896, 173921. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Flaherty, K.; Goff, S. Emerging Strategies in Systemic Therapy for the Treatment of Melanoma. Am. Soc. Clin. Oncol. Educ. Book. Am. Soc. Clin. Oncol. Annu. Meet. 2018, 38, 751–758. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Inhibitor | Phase | Medication | Tumor Type | ID | Status |
|---|---|---|---|---|---|
| Indoximod (1-D-MT) | I | Combination with idarubicin and cytarabine | Acute myeloid leukemia | NCT02835729 | 2019-12 completed |
| II | Combination with docetaxel or paclitaxel | Metastatic breast cancer | NCT01792050 | 2017-07 completed | |
| I/II | Combination with ipilimumab, pembrolizumab and nivolumab | Metastatic melanoma | NCT02073123 | 2019-07 completed | |
| II | Combination with sipuleucel-T | Metastatic prostate cancer | NCT01560923 | 2018-12 completed | |
| I | Combination with ibrutinib, cyclophosphamide and etoposide | Pediatric brain cancer | NCT05106296 | Recruiting | |
| I/II | Combination with temozolomide and bevacizumab | Newly diagnosed glioblastoma | NCT02052648 | 2019-06 completed | |
| I | Single agent | Advanced malignancy | NCT00567931 | 2012-09 completed | |
| I | Combination with temozolomide | Pediatric brain tumor | NCT02502708 | 2020-02 completed | |
| I/II | Combination with p53 DC vaccine | Metastatic breast cancer | NCT01042535 | 2018-02 completed | |
| I | Combination with docetaxel | Metastatic solid tumor | NCT01191216 | 2013-08 completed | |
| II | Combination with partial radiation | Progressive brain cancer | NCT04049669 | Recruiting | |
| Epacadostat (INCB024360) | II | Combination with pembrolizumab | Gastrointestinal stromal tumor | NCT03291054 | 2020-08 completed |
| I | Combination with sirolimus | Advanced malignancy | NCT03217669 | Active, not recruiting | |
| II | Combination with pembrolizumab | Gastroesophageal junction or gastric cancer | NCT03196232 | 2018-05 completed | |
| II | Single agent | Myelodysplastic syndrome | NCT01822691 | 2015-02 completed | |
| II | Combination with pembrolizumab | Locally advanced/metastatic sarcoma | NCT03414229 | Active, not recruiting | |
| II | Combination with pembrolizumab | ovarian clear cell carcinoma | NCT03602586 | Active, not recruiting | |
| I | Single agent | Advanced malignancy | NCT01195311 | 2013-07 completed | |
| I/II | Combination with durvalumab | Advanced solid tumor | NCT02318277 | 2020-10 completed | |
| III | Combination with pembrolizumab | Cisplatin-ineligible urothelial carcinoma | NCT03361865 | 2020-08 completed | |
| II | Combination with pembrolizumab | Metastatic non-small cell lung cancer | NCT03322540 | 2020-11 completed | |
| III | Combination with pembrolizumab | Metastatic urothelial carcinoma | NCT03374488 | 2020-07 completed | |
| I/II | Combination with MK-3475 | High colorectal cancer, Endometrial cancer, Head and neck cancer and Hepatocellular carcinoma | NCT02178722 | 2020-11 completed | |
| I/II | Combination with intralesional SD101 | Advanced/refractory solid tumors and lymphoma | NCT03322384 | 2020-04 completed | |
| I/II | Combination with nivolumab | Advanced solid tumors and lymphomas; squamous cell carcinoma of head and neck and non-small cell lung cancer | NCT02327078 | 2020-06 completed | |
| II | Combination with MELITAC 12.1 peptide vaccine | Stage III-IV melanoma | NCT01961115 | 2017-05 completed | |
| III | Combination with pembrolizumab | Locally advanced/metastatic renal cell carcinoma | NCT03260894 | Active, not recruiting | |
| III | Combination with pembrolizumab | Unresectable or metastatic melanoma | NCT02752074 | 2019-08 completed | |
| II | Combination with retifanlimab, INCAGN02385 and INCAGN02390 | Neoadjuvant urothelial carcinoma | NCT04586244 | Recruiting | |
| I/II | Single agent | Locally advanced rectal cancer | NCT03516708 | Recruiting | |
| I/II | Combination with DPX-Survivac and cyclophosphamide | Recurrent ovarian cancer | NCT02785250 | Active, not recruiting | |
| I | Single agent | Epithelial ovarian, fallopian tube, or primary peritoneal cancer | NCT02042430 | Active, not recruiting | |
| II | Combination with pembrolizumab | Metastatic pancreas cancer | NCT03006302 | Active, not recruiting | |
| II | Combination with pembrolizumab and platinum-based chemotherapy | Metastatic non-small cell lung cancer | NCT03322566 | 2020-10 completed | |
| II | Combination with retifanlimab and pemigatinib | Advanced or metastatic endometrial cancer | NCT04463771 | Recruiting | |
| III | Combination with pembrolizumab and cetuximab | Recurrent or metastatic head and neck squamous cell carcinoma | NCT03358472 | Active, not recruiting | |
| BMS-986205 | II | Combination with nivolumab | Endometrial cancer | NCT04106414 | Active, not recruiting |
| I/II | Combination with nivolumab | Liver cancer | NCT03695250 | Active, not recruiting | |
| I/II | Combination with nivolumab | Advanced malignant solid tumor | NCT03792750 | 2020-12 completed | |
| III | Combination with nivolumab | Advanced melanoma | NCT03329846 | 2020-07 completed | |
| II | Combination with nivolumab | Squamous cell cancer of the head and neck | NCT03854032 | Active, not recruiting | |
| I | Combination with nivolumab | Newly diagnosed glioblastoma | NCT04047706 | Active, not recruiting | |
| I/II | Combination with nivolumab, relatlimab and ipilimumab | Advanced tumor | NCT03459222 | Active, not recruiting | |
| II | Combination with nivolumab, relatlimab and ipilimumab | Advanced renal cell carcinoma | NCT02996110 | 2021-11 completed | |
| I | Combination with nivolumab | Advanced malignant tumor | NCT03192943 | 2018-12 completed | |
| I/II | Combination with nivolumab, and ipilimumab | Advanced malignant tumor | NCT02658890 | 2021-10 completed | |
| II | Combination with nivolumab, relatlimab and ipilimumab | Advanced gastric cancer | NCT02935634 | 2022-05 completed | |
| III | Combination with nivolumab, gemcitabine and cisplatin | Muscle invasive bladder cancer | NCT03661320 | Active, not recruiting | |
| I | Combination with nivolumab, relatlimab and cabiralizumab | Solid tumor | NCT03335540 | 2021-08 completed | |
| Navoximod (GDC-0919, NLG-919) | I | Single agent | Advanced solid tumor | NCT02048709 | 2016-02 completed |
| I | Combination with atezolizumab | Locally advanced or metastatic solid tumors | NCT02471846 | 2019-10 completed | |
| PF-06840003 | I | Single agent | Malignant gliomas | NCT02764151 | Terminated |
| HTI-1090 (SHR9146) | I | Single agent | Advanced solid tumor | NCT03208959 | 2019-01 completed |
| I | Combination with SHR-1210 and apatinib | Advanced solid tumor | NCT03491631 | Active, not recruiting |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.; Zhang, J. Indoleamine 2,3-Dioxygenase (IDO) Activity: A Perspective Biomarker for Laboratory Determination in Tumor Immunotherapy. Biomedicines 2023, 11, 1988. https://doi.org/10.3390/biomedicines11071988
Yang P, Zhang J. Indoleamine 2,3-Dioxygenase (IDO) Activity: A Perspective Biomarker for Laboratory Determination in Tumor Immunotherapy. Biomedicines. 2023; 11(7):1988. https://doi.org/10.3390/biomedicines11071988
Chicago/Turabian StyleYang, Pengbo, and Junhua Zhang. 2023. "Indoleamine 2,3-Dioxygenase (IDO) Activity: A Perspective Biomarker for Laboratory Determination in Tumor Immunotherapy" Biomedicines 11, no. 7: 1988. https://doi.org/10.3390/biomedicines11071988