
Thalamocortical Mechanisms Underlying Real and Imagined Acupuncture

Abstract
:1. Introduction
2. Material and Methods
2.1. Participants
2.2. Experiment Procedures
2.2.1. Session 1 Training Session
2.2.2. Sessions 2–5 Intervention Sessions
2.3. Pain Threshold Measurements
2.4. MRI Data Acquisition
2.5. MRI Data Pre-Processing
2.6. Seed-Based Functional Connectivity Analysis
3. Results
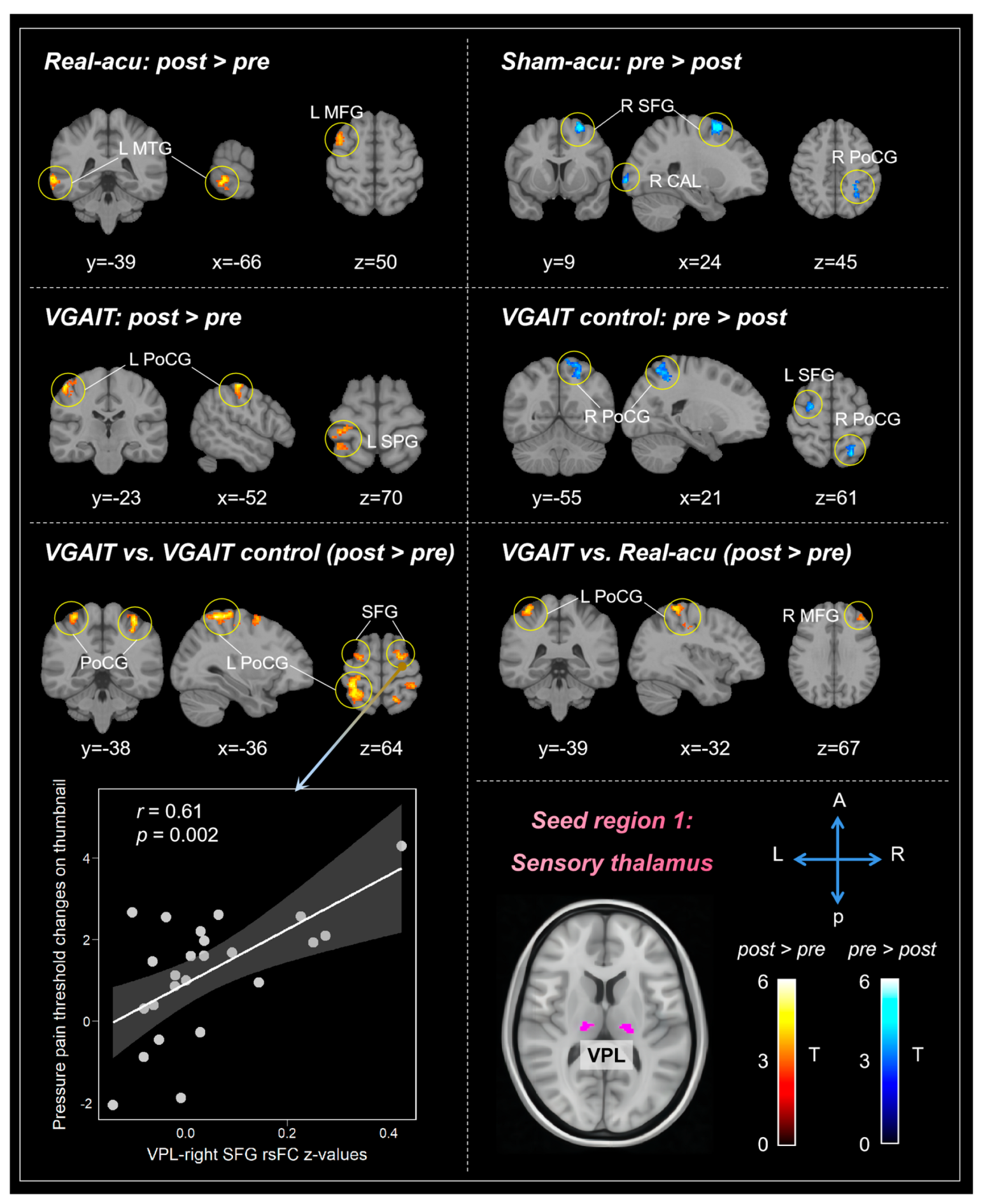
3.1. Seed-Based rsFC Results
3.1.1. Sensory Thalamus (VPL)
3.1.2. Limbic Thalamus (MD)
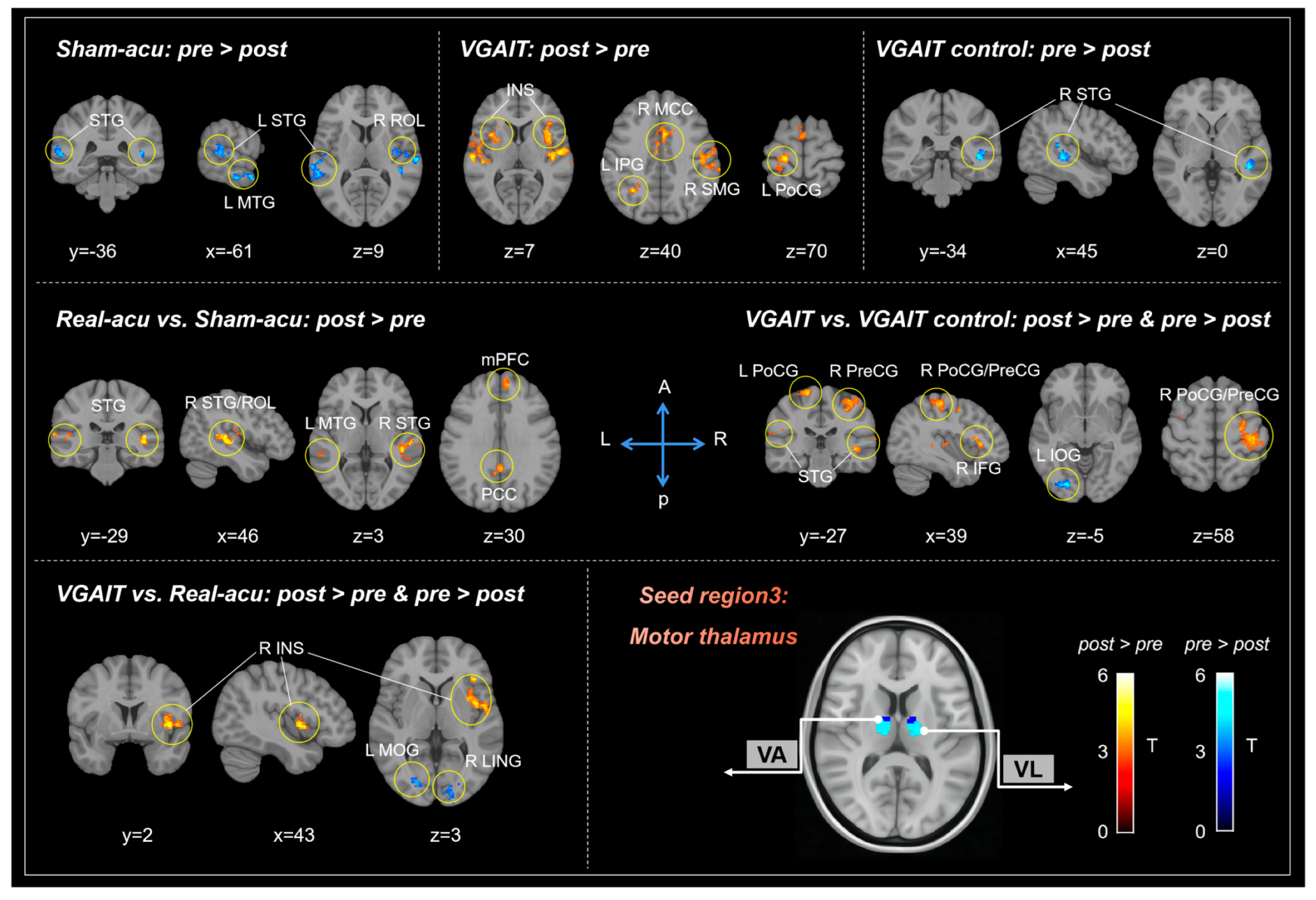
3.1.3. Motor Thalamus (Mthal)
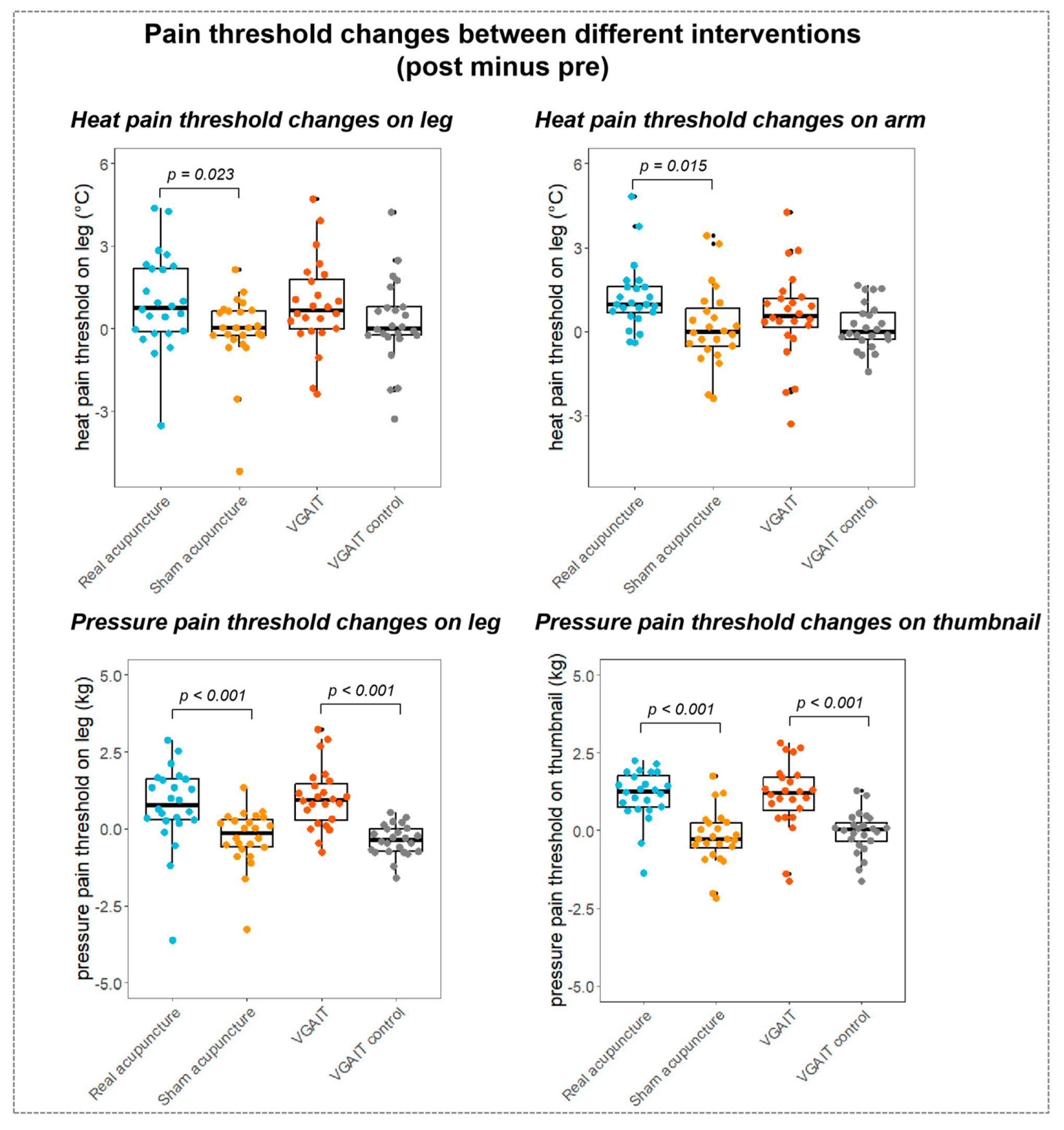
3.2. Pain Threshold Results
3.3. Associations between rsFC Differences and Pain Threshold Changes
3.3.1. Correlation between VPL-Based rsFC and Pain Threshold Changes
3.3.2. Correlation between MD-Based rsFC and Pain Threshold Changes
3.3.3. Correlation between Mthal-Based rsFC and Pain Threshold Changes
4. Discussion
4.1. The Modulation Effects of Acupuncture on Thalamocortical Circuits
4.2. The Modulation Effects of VGAIT on Thalamocortical Circuits
4.3. Comparisons of Modulation Effects between Acupuncture and VGAIT on Thalamocortical Circuits
4.4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lavand’Homme, P. The Progression from Acute to Chronic Pain. Curr. Opin. Anaesthesiol. 2011, 24, 545–550. [Google Scholar] [CrossRef]
- Michaelides, A.; Zis, P. Depression, Anxiety and Acute Pain: Links and Management Challenges. Postgrad. Med. J. 2019, 131, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Pozek, J.P.J.; Beausang, D.; Baratta, J.L.; Viscusi, E.R. The Acute to Chronic Pain Transition: Can Chronic Pain Be Prevented? Med. Clin. N. Am. 2016, 100, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Shanthanna, H.; Nelson, A.M.; Kissoon, N.; Narouze, S. The COVID-19 Pandemic and Its Consequences for Chronic Pain: A Narrative Review. Anaesthesia 2022, 77, 1039–1050. [Google Scholar] [CrossRef]
- Nahin, R.L.; Boineau, R.; Khalsa, P.S.; Stussman, B.J.; Weber, W.J. Evidence-Based Evaluation of Complementary Health Approaches for Pain Management in the United States. Mayo Clin. Proc. 2016, 91, 1292–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.B.; Roman, D.L. Novel Treatments for Chronic Pain: Moving beyond Opioids. Transl. Res. 2021, 234, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Dowell, D.; Ragan, K.R.; Jones, C.M.; Baldwin, G.T.; Chou, R. CDC Clinical Practice Guideline for Prescribing Opioids for Pain—United States, 2022. MMWR Recomm. Rep. 2022, 71, 1–95. [Google Scholar] [CrossRef] [PubMed]
- Qaseem, A.; McLean, R.M.; O’Gurek, D.; Batur, P.; Lin, K.; Kansagara, D.L. Nonpharmacologic and Pharmacologic Management of Acute Pain from Non-Low Back, Musculoskeletal Injuries in Adults: A Clinical Guideline from the American College of Physicians and American Academy of Family Physicians. Ann. Intern. Med. 2020, 173, 739–748. [Google Scholar] [CrossRef]
- Kong, J.; Eshel, M.N. Applying the Power of the Mind in Acupuncture Treatment of Pain. Med. Acupunct. 2020, 32, 367–372. [Google Scholar] [CrossRef]
- Cao, J.; Tu, Y.; Orr, S.P.; Lang, C.; Park, J.; Vangel, M.; Chen, L.; Gollub, R.; Kong, J. Analgesic Effects Evoked by Real and Imagined Acupuncture: A Neuroimaging Study. Cereb. Cortex 2019, 29, 3220–3231. [Google Scholar] [CrossRef]
- Cao, J.; Orr, S.P.; Wilson, G.; Kong, J. Imagined and Actual Acupuncture Effects on Chronic Low Back Pain: A Preliminary Study. Neural Plast. 2020, 2020, 8579743. [Google Scholar] [CrossRef]
- Jones, E.G. Thalamocortical Dysrhythmia and Chronic Pain. Pain 2010, 150, 4–5. [Google Scholar] [CrossRef]
- Tu, Y.; Fu, Z.; Mao, C.; Falahpour, M.; Gollub, R.L.; Park, J.; Wilson, G.; Napadow, V.; Gerber, J.; Chan, S.T.; et al. Distinct Thalamocortical Network Dynamics Are Associated with the Pathophysiology of Chronic Low Back Pain. Nat. Commun. 2020, 11, 3948. [Google Scholar] [CrossRef]
- Mao, C.P.; Wilson, G.; Cao, J.; Meshberg, N.; Huang, Y.; Kong, J. Abnormal Anatomical and Functional Connectivity of the Thalamo-Sensorimotor Circuit in Chronic Low Back Pain: Resting-State Functional Magnetic Resonance Imaging and Diffusion Tensor Imaging Study. Neuroscience 2022, 487, 143–154. [Google Scholar] [CrossRef]
- Ahn, S.; Prim, J.H.; Alexander, M.L.; McCulloch, K.L.; Fröhlich, F. Identifying and Engaging Neuronal Oscillations by Transcranial Alternating Current Stimulation in Patients with Chronic Low Back Pain: A Randomized, Crossover, Double-Blind, Sham-Controlled Pilot Study. J. Pain Off. J. Am. Pain Soc. 2019, 20, 277.e1. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, Y.; Li, H.; Yan, Z.; Zhang, Y.; Liu, X.; Hou, X.; Chen, W.; Tu, Y.; Hodges, S.; et al. Transcutaneous Auricular Vagus Nerve Stimulation (TaVNS) for Migraine: An FMRI Study. Reg. Anesth. Pain. Med. 2021, 46, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Biella, G.; Sotgiu, M.L.; Pellegata, G.; Paulesu, E.; Castiglioni, I.; Fazio, F. Acupuncture Produces Central Activations in Pain Regions. Neuroimage 2001, 14, 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, F.; Wang, J.Y. Modulation of Central Nociceptive Coding by Acupoint Stimulation. Neurochem. Res. 2008, 33, 1950–1955. [Google Scholar] [CrossRef]
- Volz, M.S.; Suarez-Contreras, V.; Portilla, A.L.S.; Illigens, B.; Bermpohl, F.; Fregni, F. Movement Observation-Induced Modulation of Pain Perception and Motor Cortex Excitability. Clin. Neurophysiol. 2015, 126, 1204–1211. [Google Scholar] [CrossRef]
- Müller, K.; Bacht, K.; Prochnow, D.; Schramm, S.; Seitz, R.J. Activation of Thalamus in Motor Imagery Results from Gating by Hypnosis. Neuroimage 2013, 66, 361–367. [Google Scholar] [CrossRef]
- Groh, A.; Krieger, P.; Mease, R.A.; Henderson, L. Acute and Chronic Pain Processing in the Thalamocortical System of Humans and Animal Models. Neuroscience 2018, 387, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.S.; Zhang, Y.; Halassa, M.M.; Feng, G. Thalamic Subnetworks as Units of Function. Nat. Neurosci. 2022, 25, 140. [Google Scholar] [CrossRef] [PubMed]
- Behrens, T.E.J.; Johansen-Berg, H.; Woolrich, M.W.; Smith, S.M.; Wheeler-Kingshott, C.A.M.; Boulby, P.A.; Barker, G.J.; Sillery, E.L.; Sheehan, K.; Ciccarelli, O.; et al. Non-Invasive Mapping of Connections between Human Thalamus and Cortex Using Diffusion Imaging. Nat. Neurosci. 2003, 6, 750–757. [Google Scholar] [CrossRef]
- Grodd, W.; Kumar, V.J.; Schüz, A.; Lindig, T.; Scheffler, K. The Anterior and Medial Thalamic Nuclei and the Human Limbic System: Tracing the Structural Connectivity Using Diffusion-Weighted Imaging. Sci. Rep. 2020, 10, 10957. [Google Scholar] [CrossRef]
- O’Connell, N.E.; Marston, L.; Spencer, S.; DeSouza, L.H.; Wand, B.M. Non-Invasive Brain Stimulation Techniques for Chronic Pain. Cochrane Database Syst. Rev. 2018, 4, CD008208. [Google Scholar] [CrossRef] [PubMed]
- Knotkova, H.; Hamani, C.; Sivanesan, E.; Le Beuffe, M.F.E.; Moon, J.Y.; Cohen, S.P.; Huntoon, M.A. Neuromodulation for Chronic Pain. Lancet 2021, 397, 2111–2124. [Google Scholar] [CrossRef]
- Coombes, S.A.; Misra, G. Pain and Motor Processing in the Human Cerebellum. Pain 2016, 157, 117–127. [Google Scholar] [CrossRef]
- Gaidica, M.; Hurst, A.; Cyr, C.; Leventhal, D.K. Distinct Populations of Motor Thalamic Neurons Encode Action Initiation, Action Selection, and Movement Vigor. J. Neurosci. 2018, 38, 6563–6573. [Google Scholar] [CrossRef]
- Kaelin-Lang, A.; Luft, A.R.; Sawaki, L.; Burstein, A.H.; Sohn, Y.H.; Cohen, L.G. Modulation of Human Corticomotor Excitability by Somatosensory Input. J. Physiol. 2002, 540, 623. [Google Scholar] [CrossRef]
- Yew, L.L.; Cui, S.L.; Fook-Chong, S. The Effect of Acupuncture on Motor Cortex Excitability and Plasticity. Neurosci. Lett. 2005, 384, 145–149. [Google Scholar] [CrossRef]
- Cao, J.; Tu, Y.; Wilson, G.; Orr, S.P.; Kong, J. Characterizing the Analgesic Effects of Real and Imagined Acupuncture Using Functional and Structure MRI. Neuroimage 2020, 221, 117176. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Tu, Y.; Orr, S.P.; Wilson, G.; Kong, J. Modulatory Effects of Actual and Imagined Acupuncture on the Functional Connectivity of the Periaqueductal Gray and Ventral Tegmental Area. Psychosom. Med. 2021, 83, 870–879. [Google Scholar] [CrossRef]
- Kong, J.; Gollub, R.L.; Rosman, I.S.; Megan Webb, J.; Vangel, M.G.; Kirsch, I.; Kaptchuk, T.J. Brain Activity Associated with Expectancy-Enhanced Placebo Analgesia as Measured by Functional Magnetic Resonance Imaging. J. Neurosci. 2006, 26, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Streitberger, K.; Kleinhenz, J. Introducing a Placebo Needle into Acupuncture Research. Lancet 1998, 352, 364–365. [Google Scholar] [CrossRef]
- Coronado, R.A.; Kindler, L.L.; Valencia, C.; George, S.Z. Thermal and Pressure Pain Sensitivity in Patients with Unilateral Shoulder Pain: Comparison of Involved and Uninvolved Sides. J. Orthop. Sports Phys. Ther. 2011, 41, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.; Spaeth, R.; Cook, A.; Kirsch, I.; Claggett, B.; Vangel, M.; Gollub, R.L.; Smoller, J.W.; Kaptchuk, T.J. Are All Placebo Effects Equal? Placebo Pills, Sham Acupuncture, Cue Conditioning and Their Association. PLoS ONE 2013, 8, e67485. [Google Scholar] [CrossRef] [Green Version]
- Schabrun, S.M.; Jones, E.; Elgueta Cancino, E.L.; Hodges, P.W. Targeting Chronic Recurrent Low Back Pain from the Top-down and the Bottom-up: A Combined Transcranial Direct Current Stimulation and Peripheral Electrical Stimulation Intervention. Brain Stimul. 2014, 7, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Zhang, B.; Cao, J.; Wilson, G.; Zhang, Z.; Kong, J. Identifying Inter-Individual Differences in Pain Threshold Using Brain Connectome: A Test-Retest Reproducible Study. Neuroimage 2019, 202, 116049. [Google Scholar] [CrossRef]
- Rolls, E.T.; Huang, C.C.; Lin, C.P.; Feng, J.; Joliot, M. Automated Anatomical Labelling Atlas 3. Neuroimage 2020, 206, 116189. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Nierhaus, T.; Pach, D.; Witt, C.M.; Yi, M. Default Mode Network as a Neural Substrate of Acupuncture: Evidence, Challenges and Strategy. Front. Neurosci. 2019, 13, 100. [Google Scholar] [CrossRef] [Green Version]
- Alves, P.N.; Foulon, C.; Karolis, V.; Bzdok, D.; Margulies, D.S.; Volle, E.; Thiebaut de Schotten, M. An Improved Neuroanatomical Model of the Default-Mode Network Reconciles Previous Neuroimaging and Neuropathological Findings. Commun. Biol. 2019, 2, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, B.J.; Davey, C.G.; Savage, H.S.; Jamieson, A.J.; Leonards, C.A.; Moffat, B.A.; Glarin, R.K.; Steward, T. Dynamic Subcortical Modulators of Human Default Mode Network Function. Cerebral Cortex 2022, 32, 4345. [Google Scholar] [CrossRef]
- Argaman, Y.; Kisler, L.B.; Granovsky, Y.; Coghill, R.C.; Sprecher, E.; Manor, D.; Weissman-Fogel, I. The Endogenous Analgesia Signature in the Resting Brain of Healthy Adults and Migraineurs. J. Pain. 2020, 21, 905–918. [Google Scholar] [CrossRef]
- Zhang, P.; Jiang, Y.; Liu, G.; Han, J.; Wang, J.; Ma, L.; Hu, W.; Zhang, J. Altered Brain Functional Network Dynamics in Classic Trigeminal Neuralgia: A Resting-State Functional Magnetic Resonance Imaging Study. J. Headache Pain 2021, 22, 147. [Google Scholar] [CrossRef] [PubMed]
- Coppola, G.; Di Renzo, A.; Tinelli, E.; Petolicchio, B.; Parisi, V.; Serrao, M.; Porcaro, C.; Fiorelli, M.; Caramia, F.; Schoenen, J.; et al. Thalamo-Cortical Networks in Subtypes of Migraine with Aura Patients. J. Headache Pain 2021, 22, 58. [Google Scholar] [CrossRef] [PubMed]
- Ichesco, E.; Puiu, T.; Hampson, J.P.; Kairys, A.E.; Clauw, D.J.; Harte, S.E.; Peltier, S.J.; Harris, R.E.; Schmidt-Wilcke, T. Altered FMRI Resting-State Connectivity in Individuals with Fibromyalgia on Acute Pain Stimulation. Eur. J. Pain 2016, 20, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Riegner, G.; Posey, G.; Oliva, V.; Jung, Y.; Mobley, W.; Zeidan, F. Disentangling Self from Pain: Mindfulness Meditation-Induced Pain Relief Is Driven by Thalamic-Default Mode Network Decoupling. Pain 2023, 164, 280–291. [Google Scholar] [CrossRef]
- Ong, W.Y.; Stohler, C.S.; Herr, D.R. Role of the Prefrontal Cortex in Pain Processing. Mol. Neurobiol. 2019, 56, 1137. [Google Scholar] [CrossRef] [Green Version]
- Favilla, S.; Huber, A.; Pagnoni, G.; Lui, F.; Facchin, P.; Cocchi, M.; Baraldi, P.; Porro, C.A. Ranking Brain Areas Encoding the Perceived Level of Pain from FMRI Data. Neuroimage 2014, 90, 153–162. [Google Scholar] [CrossRef]
- Balducci, T.; Garza-Villarreal, E.A.; Valencia, A.; Aleman, A.; van Tol, M.J. Abnormal Functional Neurocircuitry Underpinning Emotional Processing in Fibromyalgia. Eur. Arch. Psychiatry Clin. Neurosci. 2023. [Google Scholar] [CrossRef]
- Kamping, S.; Andoh, J.; Bomba, I.C.; Diers, M.; Diesch, E.; Flor, H. Contextual Modulation of Pain in Masochists: Involvement of the Parietal Operculum and Insula. Pain 2016, 157, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, J.; White, N.S.; Kwong, K.K.; Vangel, M.G.; Rosman, I.S.; Gracely, R.H.; Gollub, R.L. Using FMRI to Dissociate Sensory Encoding from Cognitive Evaluation of Heat Pain Intensity. Hum. Brain Mapp. 2006, 27, 715–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.G.; Schloesser, D.; Arensdorf, A.M.; Simmons, J.M.; Cui, C.; Valentino, R.; Gnadt, J.W.; Nielsen, L.; Hillaire-Clarke, C.S.; Spruance, V.; et al. The Emerging Science of Interoception: Sensing, Integrating, Interpreting, and Regulating Signals within the Self. Trends Neurosci. 2021, 44, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Khalsa, S.S.; Adolphs, R.; Cameron, O.G.; Critchley, H.D.; Davenport, P.W.; Feinstein, J.S.; Feusner, J.D.; Garfinkel, S.N.; Lane, R.D.; Mehling, W.E.; et al. Interoception and Mental Health: A Roadmap. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 501–513. [Google Scholar] [CrossRef]
- Berntson, G.G.; Khalsa, S.S. Neural Circuits of Interoception. Trends Neurosci. 2021, 44, 17–28. [Google Scholar] [CrossRef]
- Chen, Z.; Xiao, L.; Liu, H.; Zhang, Q.; Wang, Q.; Lv, Y.; Zhai, Y.; Zhang, J.; Dong, S.; Wei, X.; et al. Altered Thalamo-Cortical Functional Connectivity in Patients with Vestibular Migraine: A Resting-State FMRI Study. Neuroradiology 2022, 64, 119–127. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.; Zeng, M.; He, J.; Qi, G.; Zhang, S.; Liu, R. Abnormal Whole Brain Functional Connectivity Pattern Homogeneity and Couplings in Migraine Without Aura. Front. Hum. Neurosci. 2020, 14, 551. [Google Scholar] [CrossRef]
- Chen, B.; Guo, Q.; Zhang, Q.; Di, Z.; Zhang, Q. Revealing the Central Mechanism of Acupuncture for Primary Dysmenorrhea Based on Neuroimaging: A Narrative Review. Pain. Res. Manag. 2023, 2023, 8307249. [Google Scholar] [CrossRef]
- Flodin, P.; Martinsen, S.; Löfgren, M.; Bileviciute-Ljungar, I.; Kosek, E.; Fransson, P. Fibromyalgia Is Associated with Decreased Connectivity between Pain- and Sensorimotor Brain Areas. Brain Connect. 2014, 4, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Weng, H.Y.; Feldman, J.L.; Leggio, L.; Napadow, V.; Park, J.; Price, C.J. Interventions and Manipulations of Interoception. Trends Neurosci. 2021, 44, 52. [Google Scholar] [CrossRef] [PubMed]
- Zeidan, F.; Emerson, N.M.; Farris, S.R.; Ray, J.N.; Jung, Y.; McHaffie, J.G.; Coghill, R.C. Mindfulness Meditation-Based Pain Relief Employs Different Neural Mechanisms Than Placebo and Sham Mindfulness Meditation-Induced Analgesia. J. Neurosci. 2015, 35, 15307. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, S.S.; Oliveira, L.R.; DeSantana, J.M. Correlations between Brain Changes and Pain Management after Cognitive and Meditative Therapies: A Systematic Review of Neuroimaging Studies. Complement. Ther. Med. 2018, 39, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zhang, Y.Q. A New Perspective on the Anterior Cingulate Cortex and Affective Pain. Neurosci. Biobehav. Rev. 2018, 90, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, P.N.; Peng, Y.B.; Boyette-Davis, J.A.; Uhelski, M.L. The Anterior Cingulate Cortex and Pain Processing. Front. Integr. Neurosci. 2014, 8, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, A.D.B. Significance of the Insula for the Evolution of Human Awareness of Feelings from the Body. Ann. N. Y Acad. Sci. 2011, 1225, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q.; Nomi, J.S.; Hébert-Seropian, B.; Ghaziri, J.; Boucher, O. Structure and Function of the Human Insula. J. Clin. Neurophysiol. 2017, 34, 300. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.T.; Lu, P.L. Thalamus and Pain. Acta Anaesthesiol. Taiwan 2013, 51, 73–80. [Google Scholar] [CrossRef]
- Haggard, P.; Iannetti, G.D.; Longo, M.R. Spatial Sensory Organization and Body Representation in Pain Perception. Curr. Biol. 2013, 23, R164–R176. [Google Scholar] [CrossRef] [Green Version]
- Preston, C.; Newport, R. Analgesic Effects of Multisensory Illusions in Osteoarthritis. Rheumatology 2011, 50, 2314–2315. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Lin, C.P.; Liu, H.L.; Hsu, Y.Y.; Lim, K.E.; Hung, D.; Decety, J. Expertise Modulates the Perception of Pain in Others. Curr. Biol. 2007, 17, 1708–1713. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Ye, C.; Wang, H.; Mao, Y.; Zhang, W.; Liu, A.; Yang, C.L.; Li, T.; Hayashi, L.; Zhao, W.; et al. Sound Induces Analgesia through Corticothalamic Circuits. Science 2022, 377, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.R.; Iannetti, G.D.; Mancini, F.; Driver, J.; Haggard, P. Linking Pain and the Body: Neural Correlates of Visually Induced Analgesia. J. Neurosci. 2012, 32, 2601–2607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, Z.; Gangadharan, V.; Liu, S.; Körber, C.; Tan, L.L.; Li, H.; Oswald, M.J.; Kang, J.; Martin-Cortecero, J.; Männich, D.; et al. Layer-Specific Pain Relief Pathways Originating from Primary Motor Cortex. Science 2022, 378, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Hamani, C.; Fonoff, E.T.; Parravano, D.C.; Silva, V.A.; Galhardoni, R.; Monaco, B.A.; Navarro, J.; Yeng, L.T.; Teixeira, M.J.; De Andrade, D.C. Motor Cortex Stimulation for Chronic Neuropathic Pain: Results of a Double-Blind Randomized Study. Brain 2021, 144, 2994–3004. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.M.; Graven-Nielsen, T.; De Andrade, D.C. New Updates on Transcranial Magnetic Stimulation in Chronic Pain. Curr. Opin. Support. Palliat. Care 2022, 16, 65–70. [Google Scholar] [CrossRef]
- Gunduz, M.E.; Pacheco-Barrios, K.; Bonin Pinto, C.; Duarte, D.; Vélez, F.G.S.; Gianlorenco, A.C.L.; Teixeira, P.E.P.; Giannoni-Luza, S.; Crandell, D.; Battistella, L.R.; et al. Effects of Combined and Alone Transcranial Motor Cortex Stimulation and Mirror Therapy in Phantom Limb Pain: A Randomized Factorial Trial. Neurorehabil Neural Repair. 2021, 35, 704–716. [Google Scholar] [CrossRef]
- Kim, J.; Ryu, S.B.; Lee, S.E.; Shin, J.; Jung, H.H.; Kim, S.J.; Kim, K.H.; Chang, J.W. Motor Cortex Stimulation and Neuropathic Pain: How Does Motor Cortex Stimulation Affect Pain-Signaling Pathways? J. Neurosurg. 2016, 124, 866–876. [Google Scholar] [CrossRef] [Green Version]
- Eccleston, C. Chronic Pain as Embodied Defence: Implications for Current and Future Psychological Treatments. Pain 2018, 159 (Suppl. S1), S17–S23. [Google Scholar] [CrossRef]
- Herbert, B.M.; Pollatos, O. The Body in the Mind: On the Relationship Between Interoception and Embodiment. Top. Cogn. Sci. 2012, 4, 692–704. [Google Scholar] [CrossRef]
- Pollatos, O.; Füstös, J.; Critchley, H.D. On the Generalised Embodiment of Pain: How Interoceptive Sensitivity Modulates Cutaneous Pain Perception. Pain. 2012, 153, 1680–1686. [Google Scholar] [CrossRef]
- Asma, S.T. Imagination: A New Foundation for the Science of Mind. Biol. Theory 2022, 17, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; Van Essen, D.C.; Raichle, M.E. The Human Brain Is Intrinsically Organized into Dynamic, Anticorrelated Functional Networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar] [CrossRef] [Green Version]
- Di, X.; Biswal, B.B. Modulatory Interactions between the Default Mode Network and Task Positive Networks in Resting-State. PeerJ 2014, 2, e367. [Google Scholar] [CrossRef] [Green Version]
- Limanowski, J.; Lopes, P.; Keck, J.; Baudisch, P.; Friston, K.; Blankenburg, F. Action-Dependent Processing of Touch in the Human Parietal Operculum and Posterior Insula. Cerebral Cortex 2020, 30, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.E.; Kosslyn, S.M. Cognitive Psychology: Mind and Brain; Prentice Hall: New Jersey, NJ, USA, 2007; pp. 194–199. [Google Scholar]
- Berger, C.C.; Henrik Ehrsson, H. The Fusion of Mental Imagery and Sensation in the Temporal Association Cortex. J. Neurosci. 2014, 34, 13684–13692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Song, Q.; Zhang, R.; Zhou, Y.; Kong, Y. Enhanced Temporal Coupling between Thalamus and Dorsolateral Prefrontal Cortex Mediates Chronic Low Back Pain and Depression. Neural Plast. 2021, 2021, 7498714. [Google Scholar] [CrossRef]
- Angst, M.S.; Tingle, M.; Phillips, N.G.; Carvalho, B. Determining Heat and Mechanical Pain Threshold in Inflamed Skin of Human Subjects. J. Vis. Exp. 2009, 1092. [Google Scholar] [CrossRef]
- Beissner, F.; Brandau, A.; Henke, C.; Felden, L.; Baumgärtner, U.; Treede, R.D.; Oertel, B.G.; Lötsch, J. Quick Discrimination of A(Delta) and C Fiber Mediated Pain Based on Three Verbal Descriptors. PLoS ONE 2010, 5, e12944. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Contrasts (n = 24) | Brain Regions a | Cluster Size | Peak z-Value | Peak MNI Coordinates | |||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Real-acu: post vs. pre | post > pre | L Middle temporal gyrus | 165 | 4.02 | −68 | −38 | −6 |
| L Middle frontal gyrus | 114 | 3.51 | −38 | 14 | 56 | ||
| pre > post | No regions survive the threshold | ||||||
| Sham-acu: post vs. pre | post > pre | No regions survive the threshold | |||||
| pre > post | R Superior frontal gyrus | 203 | 4.25 | 26 | 8 | 62 | |
| R Calcarine | 197 | 3.93 | 20 | −104 | −2 | ||
| R Postcentral gyrus | 169 | 3.67 | 30 | −42 | 46 | ||
| VGAIT: post vs. pre | post > pre | L Postcentral gyrus | 170 | 4.18 | −54 | −22 | 58 |
| L Superior parietal gyrus | 151 | 3.58 | −32 | −44 | 68 | ||
| pre > post | No regions survive the threshold | ||||||
| VGAIT-control: post vs. pre | post > pre | No regions survive the threshold | |||||
| pre > post | R Postcentral gyrus | 249 | 4.07 | 14 | −52 | 68 | |
| L Superior frontal gyrus | 151 | 3.87 | −24 | 0 | 66 | ||
| Real-acu vs. Sham-acu | No regions survive the threshold | ||||||
| VGAIT vs. VGAIT control | post > pre | L Postcentral gyrus | 898 | 4.45 | −32 | −48 | 66 |
| R Postcentral gyrus | 814 | 4.36 | 36 | −38 | 50 | ||
| L Superior frontal gyrus | 187 | 3.73 | −24 | −4 | 66 | ||
| R Superior frontal gyrus | 136 | 4.04 | 20 | −2 | 72 | ||
| R Superior frontal gyrus | 107 | 4.31 | 28 | −12 | 48 | ||
| pre > post | No regions survive the threshold | ||||||
| VGAIT vs. Real-acu | post > pre | L Postcentral gyrus | 369 | 4.04 | −30 | −42 | 68 |
| R Middle frontal gyrus | 128 | 3.25 | 44 | 40 | 34 | ||
| pre > post | No regions survive the threshold | ||||||
| Contrasts (n = 24) | Brain Regions a | Cluster Size | Peak z-Value | Peak MNI Coordinates | |||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Real-acu: post vs. pre | post > pre | R Middle temporal gyrus | 364 | 4.00 | 62 | −30 | −12 |
| L Angular gyrus | 224 | 3.75 | −50 | −70 | 42 | ||
| R Angular gyrus | 216 | 3.54 | 40 | −62 | 42 | ||
| R Middle frontal gyrus | 139 | 4.12 | 34 | 24 | 54 | ||
| R Inferior frontal gyrus | 139 | 4.18 | 48 | 36 | 10 | ||
| R Cerebellar Lobule VI | 98 | 3.66 | 8 | −82 | −16 | ||
| R Lingual gyrus | 96 | 3.61 | 16 | −74 | −6 | ||
| L Middle occipital gyrus | 87 | 4.47 | −24 | −86 | 6 | ||
| L Middle temporal gyrus | 78 | 4.08 | −66 | −24 | −8 | ||
| pre > post | L Precuneus | 405 | 4.24 | −12 | −50 | 66 | |
| Sham-acu: post vs. pre | post > pre | No regions survive the threshold | |||||
| pre > post | R Posterior cingulate cortex | 389 | 4.13 | 6 | −38 | 30 | |
| Bil Cerebellum lobule IX | 180 | 5.05 | 2 | −46 | −42 | ||
| L Middle temporal gyrus | 121 | 3.63 | −56 | −18 | −24 | ||
| R Middle temporal gyrus | 111 | 3.64 | 68 | −12 | −12 | ||
| VGAIT: post vs. pre | post > pre | R Anterior insula | 388 | 3.97 | 38 | 4 | 2 |
| Bil Supplementary motor area | 233 | 3.54 | 2 | 22 | 50 | ||
| R Superior temporal pole | 208 | 4.42 | 46 | 20 | −18 | ||
| L Anterior insula | 192 | 4.05 | −24 | 22 | 6 | ||
| L Anterior insula | 153 | 4.06 | −34 | 4 | 8 | ||
| L Middle frontal gyrus | 143 | 3.83 | −40 | 54 | 8 | ||
| R Middle frontal gyrus | 114 | 3.45 | 42 | 36 | 14 | ||
| L Superior temporal gyrus | 106 | 3.33 | −52 | −24 | 4 | ||
| pre > post | No regions survive the threshold | ||||||
| VGAIT control: post vs. pre | post > pre | R Cerebellum lobule VI | 181 | 4.44 | 30 | −46 | −28 |
| pre > post | R Superior temporal pole | 152 | 4.85 | 54 | 16 | −22 | |
| Real-acu vs. Sham-acu | post > pre | R Lingual gyrus | 678 | 4.36 | 12 | −100 | −14 |
| R Middle temporal gyrus | 510 | 4.62 | 66 | −24 | −12 | ||
| L Lingual gyrus | 301 | 3.94 | −26 | −90 | −20 | ||
| L Angular gyrus | 200 | 4.01 | −50 | −70 | 42 | ||
| R Angular gyrus | 193 | 4.43 | 40 | −60 | 42 | ||
| R Middle frontal gyrus | 121 | 4.14 | 38 | 14 | 62 | ||
| Bil Posterior cingulate gyrus | 111 | 3.42 | −2 | −52 | 28 | ||
| L Parahippocampal gyrus | 110 | 4.34 | −34 | −6 | −26 | ||
| pre > post | L Precuneus | 323 | 4.49 | −10 | −48 | 72 | |
| R Middle cingulate gyrus | 299 | 5.06 | 6 | 12 | 42 | ||
| R Middle frontal gyrus | 104 | 3.95 | 34 | 50 | 30 | ||
| R Postcentral gyrus | 103 | 3.57 | 20 | −38 | 74 | ||
| VGAIT vs. VGAIT control | post > pre | R Superior temporal pole | 609 | 4.67 | 46 | 20 | −20 |
| Bil Supplementary motor area/Anterior cingulate cortex | 508 | 4.53 | 0 | 20 | 52 | ||
| R Middle frontal gyrus | 244 | 3.99 | 44 | 50 | 14 | ||
| R Precentral gyrus | 112 | 4.24 | 28 | −10 | 50 | ||
| L Middle frontal gyrus | 105 | 3.48 | −38 | 52 | 6 | ||
| pre > post | No regions survive the threshold | ||||||
| VGAIT vs. Real-acu | post > pre | Bil Anterior cingulate cortex | 393 | 3.71 | 0 | 32 | 24 |
| R Anterior insula | 293 | 4.25 | 38 | 6 | 2 | ||
| L Anterior insula | 211 | 4.45 | −36 | 4 | 6 | ||
| L Supramarginal gyrus | 113 | 3.51 | −56 | −28 | 26 | ||
| L Middle frontal gyrus | 105 | 3.57 | −34 | 42 | 38 | ||
| pre > post | No regions survive the threshold | ||||||
| Contrasts (n = 24) | Brain Regions a | Cluster Size | Peak z-Value | Peak MNI Coordinates | |||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Real-acu: post vs. pre | No regions survive the threshold | ||||||
| Sham-acu: post vs. pre | post > pre | No regions survive the threshold | |||||
| pre > post | L Superior temporal gyrus | 281 | 3.76 | −68 | −30 | 8 | |
| R Superior temporal gyrus | 231 | 4.16 | 44 | −32 | 6 | ||
| R Rolandic operculum | 172 | 4.03 | 52 | −16 | 14 | ||
| R Middle temporal gyrus | 153 | 4.98 | 48 | −12 | −18 | ||
| L Middle temporal gyrus | 139 | 4.28 | −60 | 2 | −22 | ||
| VGAIT: post vs. pre | post > pre | R Insula | 1305 | 4.51 | 44 | 2 | 4 |
| R Middle cingulate cortex | 881 | 4.55 | 6 | 14 | 36 | ||
| L Superior temporal gyrus | 406 | 4.18 | −62 | −18 | 10 | ||
| R Supramarginal gyrus | 233 | 4.12 | 50 | −24 | 30 | ||
| L Insula | 192 | 4.38 | −30 | 2 | 12 | ||
| L Postcentral gyrus | 146 | 3.80 | −20 | −26 | 72 | ||
| L Inferior parietal gyrus | 121 | 3.62 | −32 | −54 | 40 | ||
| pre > post | No regions survive the threshold | ||||||
| VGAIT-control: post vs. pre | post > pre | No regions survive the threshold | |||||
| pre > post | R Superior temporal gyrus | 149 | 4.12 | 44 | −36 | 0 | |
| Real-acu vs. Sham-acu | post > pre | R Superior temporal gyrus | 231 | 4.38 | 48 | −30 | 8 |
| L Superior temporal gyrus | 225 | 3.83 | −50 | −40 | 22 | ||
| L Middle temporal gyrus | 135 | 3.86 | −60 | 2 | −24 | ||
| R Rolandic operculum | 128 | 4.05 | 52 | −16 | 16 | ||
| Bil Posterior cingulate cortex | 114 | 4.06 | 0 | −50 | 26 | ||
| R Medial prefrontal cortex | 96 | 3.41 | 10 | 50 | 30 | ||
| pre > post | No regions survive the threshold | ||||||
| VGAIT vs. VGAIT control | post > pre | R Postcentral gyrus | 830 | 4.32 | 28 | −10 | 52 |
| R Superior temporal gyrus | 425 | 4.05 | 46 | −36 | −2 | ||
| L Superior temporal gyrus | 354 | 4.72 | −62 | −18 | 10 | ||
| R Superior temporal gyrus | 333 | 3.82 | 58 | −4 | 4 | ||
| R Superior temporal pole | 333 | 4.95 | 56 | 18 | −8 | ||
| R Middle temporal gyrus | 233 | 3.94 | 62 | −58 | 10 | ||
| L Postcentral gyrus | 146 | 3.68 | −20 | −26 | 72 | ||
| L Superior temporal gyrus | 145 | 3.55 | −62 | −50 | 14 | ||
| L Precentral gyrus | 140 | 4.27 | −38 | 0 | 42 | ||
| R Precentral gyrus | 132 | 4.01 | 58 | 8 | 38 | ||
| R Inferior temporal gyrus | 114 | 4.00 | 50 | −52 | −14 | ||
| L Middle temporal gyrus | 100 | 4.37 | −38 | −62 | 18 | ||
| R Inferior frontal gyrus | 94 | 3.73 | 44 | 26 | 4 | ||
| pre > post | L Inferior occipital gyrus | 140 | 4.01 | −24 | −84 | −6 | |
| VGAIT vs. Real-acu | post > pre | R Insula | 352 | 4.04 | 44 | 0 | 2 |
| pre > post | R Lingual gyrus | 209 | 3.76 | 22 | −76 | 2 | |
| L Middle occipital gyrus | 176 | 3.88 | −26 | −78 | −2 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, Q.; Sacca, V.; Walker, K.; Hodges, S.; Kong, J. Thalamocortical Mechanisms Underlying Real and Imagined Acupuncture. Biomedicines 2023, 11, 1830. https://doi.org/10.3390/biomedicines11071830
Kong Q, Sacca V, Walker K, Hodges S, Kong J. Thalamocortical Mechanisms Underlying Real and Imagined Acupuncture. Biomedicines. 2023; 11(7):1830. https://doi.org/10.3390/biomedicines11071830
Chicago/Turabian StyleKong, Qiao, Valeria Sacca, Kathryn Walker, Sierra Hodges, and Jian Kong. 2023. "Thalamocortical Mechanisms Underlying Real and Imagined Acupuncture" Biomedicines 11, no. 7: 1830. https://doi.org/10.3390/biomedicines11071830