
Association of the rs3039851 Insertion/Deletion in the Gene PPP3R1, Which Encodes the Regulatory Calcineurin Subunit B Type 1, with Left Ventricular Mass in Polish Full-Term Newborns
 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Newborns
2.2. Echocardiographic Measurements
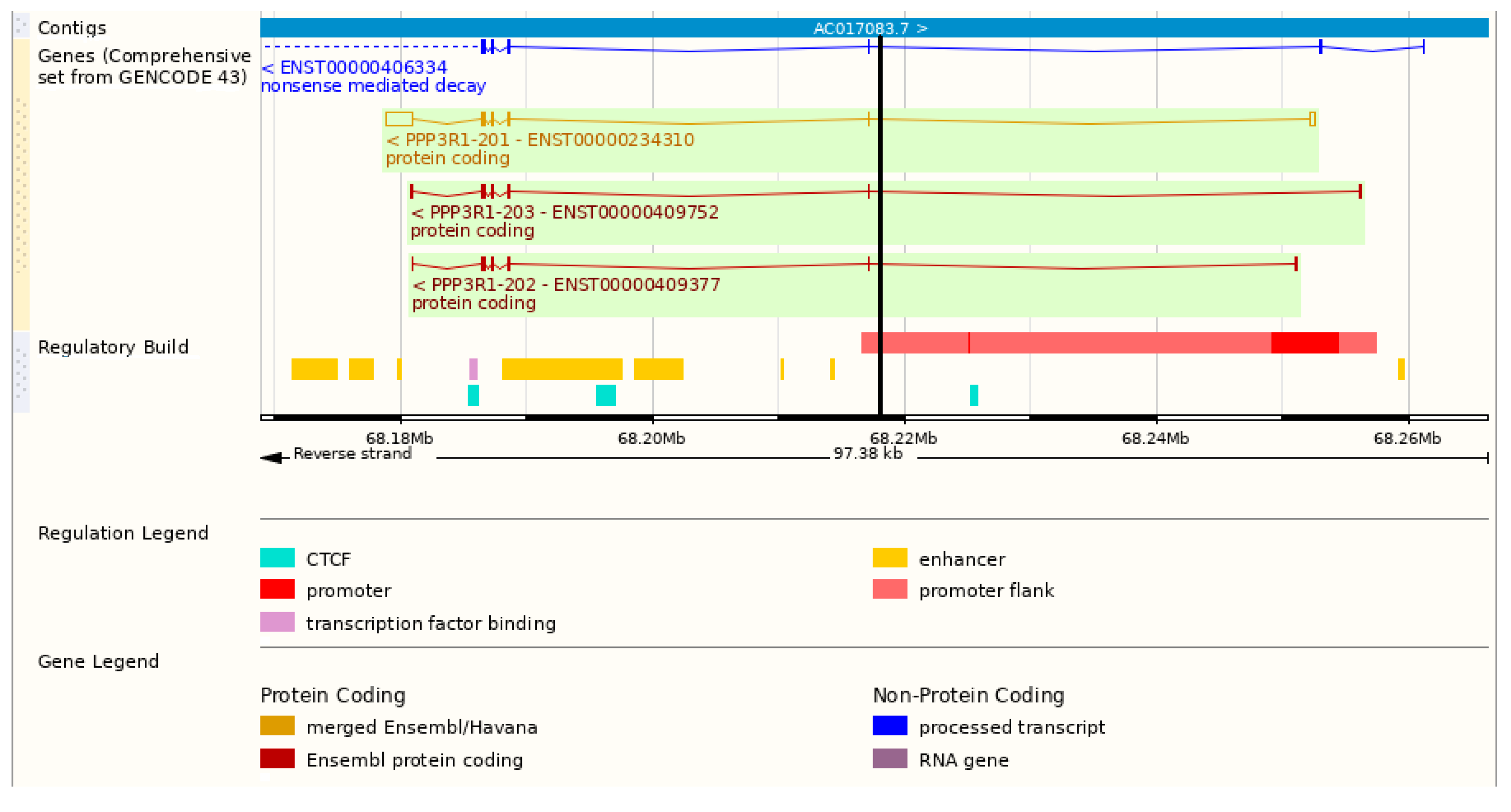
2.3. PPP3R1 Genotyping
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Levy, D.; Garrison, R.J.; Savage, D.D.; Kannel, W.B.; Castelli, W.P. Prognostic implications of echocardiographically determined left ventricular mass in the Framingham Heart Study. N. Engl. J. Med. 1990, 322, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Frey, N.; Olson, E.N. Cardiac hypertrophy: The good, the bad, and the ugly. Annu. Rev. Physiol. 2003, 65, 45–79. [Google Scholar] [CrossRef] [PubMed]
- Swan, L.; Birnie, D.H.; Padmanabhan, S.; Inglis, G.; Connell, J.M.; Hillis, W.S. The genetic determination of left ventricular mass in healthy adults. Eur. Heart J. 2003, 24, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Middelberg, R.P.; Andrew, T.; Johnson, M.R.; Christley, H.; Brown, M.J. Heritability of left ventricular mass in a large cohort of twins. J. Hypertens. 2006, 24, 321–324. [Google Scholar] [CrossRef]
- Bella, J.N.; MacCluer, J.W.; Roman, M.J.; Almasy, L.; North, K.E.; Best, L.G.; Lee, E.T.; Fabsitz, R.R.; Howard, B.V.; Devereux, R.B. Heritability of left ventricular dimensions and mass in American Indians: The Strong Heart Study. J. Hypertens. 2004, 22, 281–286. [Google Scholar] [CrossRef]
- Assimes, T.L.; Narasimhan, B.; Seto, T.B.; Yoon, S.; Curb, J.D.; Olshen, R.A.; Quertermous, T. Heritability of left ventricular mass in Japanese families living in Hawaii: The SAPPHIRe Study. J. Hypertens. 2007, 25, 985–992. [Google Scholar] [CrossRef]
- Verhaaren, H.A.; Schieken, R.M.; Mosteller, M.; Hewitt, J.K.; Eaves, L.J.; Nance, W.E. Bivariate genetic analysis of left ventricular mass and weight in pubertal twins (the Medical College of Virginia twin study). Am. J. Cardiol. 1991, 68, 661–668. [Google Scholar] [CrossRef]
- Hardingham, G.E.; Bading, H. Nuclear calcium: A key regulator of gene expression. Biometals 1998, 11, 345–358. [Google Scholar] [CrossRef]
- Chen, L.; Song, M.; Yao, C. Calcineurin in development and disease. Genes Dis. 2022, 9, 915–927. [Google Scholar] [CrossRef]
- Sakaue, S.; Kanai, M.; Tanigawa, Y.; Karjalainen, J.; Kurki, M.; Koshiba, S.; Narita, A.; Konuma, T.; Yamamoto, K.; Akiyama, M.; et al. A cross-population atlas of genetic associations for 220 human phenotypes. Nat. Genet. 2021, 53, 1415–1424. [Google Scholar] [CrossRef]
- Surendran, P.; Feofanova, E.V.; Lahrouchi, N.; Ntalla, I.; Karthikeyan, S.; Cook, J.; Chen, L.; Mifsud, B.; Yao, C.; Kraja, A.T.; et al. Discovery of rare variants associated with blood pressure regulation through meta-analysis of 1.3 million individuals. Nat. Genet. 2020, 52, 1314–1332. [Google Scholar] [CrossRef]
- Plotnikov, D.; Huang, Y.; Khawaja, A.P.; Foster, P.J.; Zhu, Z.; Guggenheim, J.A.; He, M. High Blood Pressure and Intraocular Pressure: A Mendelian Randomization Study. Investig. Ophthalmol. Vis. Sci. 2022, 63, 29. [Google Scholar] [CrossRef]
- Molkentin, J.D.; Lu, J.R.; Antos, C.L.; Markham, B.; Richardson, J.; Robbins, J.; Grant, S.R.; Olson, E.N. A calcineurin-dependent transcriptional pathway for cardiac hypertrophy. Cell 1998, 93, 215–228. [Google Scholar] [CrossRef]
- Bueno, O.F.; Wilkins, B.J.; Tymitz, K.M.; Glascock, B.J.; Kimball, T.F.; Lorenz, J.N.; Molkentin, J.D. Impaired cardiac hypertrophic response in Calcineurin Abeta -deficient mice. Proc. Natl. Acad. Sci. USA 2002, 99, 4586–4591. [Google Scholar] [CrossRef]
- Felkin, L.E.; Narita, T.; Germack, R.; Shintani, Y.; Takahashi, K.; Sarathchandra, P.; López-Olañeta, M.M.; Gómez-Salinero, J.M.; Suzuki, K.; Barton, P.J.; et al. Calcineurin splicing variant calcineurin Aβ1 improves cardiac function after myocardial infarction without inducing hypertrophy. Circulation 2011, 123, 2838–2847. [Google Scholar] [CrossRef]
- Kichaev, G.; Bhatia, G.; Loh, P.R.; Gazal, S.; Burch, K.; Freund, M.K.; Schoech, A.; Pasaniuc, B.; Price, A.L. Leveraging Polygenic Functional Enrichment to Improve GWAS Power. Am. J. Human Genet. 2019, 104, 65–75. [Google Scholar] [CrossRef]
- Giri, A.; Hellwege, J.N.; Keaton, J.M.; Park, J.; Qiu, C.; Warren, H.R.; Torstenson, E.S.; Kovesdy, C.P.; Sun, Y.V.; Wilson, O.D.; et al. Trans-ethnic association study of blood pressure determinants in over 750,000 individuals. Nat. Genet. 2019, 51, 51–62. [Google Scholar] [CrossRef]
- Maillet, M.; Davis, J.; Auger-Messier, M.; York, A.; Osinska, H.; Piquereau, J.; Lorenz, J.N.; Robbins, J.; Ventura-Clapier, R.; Molkentin, J.D. Heart-specific deletion of CnB1 reveals multiple mechanisms whereby calcineurin regulates cardiac growth and function. J. Biol. Chem. 2010, 285, 6716–6724. [Google Scholar] [CrossRef]
- Schaeffer, P.J.; Desantiago, J.; Yang, J.; Flagg, T.P.; Kovacs, A.; Weinheimer, C.J.; Courtois, M.; Leone, T.C.; Nichols, C.G.; Bers, D.M.; et al. Impaired contractile function and calcium handling in hearts of cardiac-specific calcineurin b1-deficient mice. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1263–H1273. [Google Scholar] [CrossRef]
- Parra, V.; Rothermel, B.A. Calcineurin signaling in the heart: The importance of time and place. J. Mol. Cell Cardiol. 2017, 103, 121–136. [Google Scholar] [CrossRef]
- Tang, W.; Arnett, D.K.; Devereux, R.B.; Panagiotou, D.; Province, M.A.; Miller, M.B.; de Simone, G.; Gu, C.; Ferrell, R.E. Identification of a novel 5-base pair deletion in calcineurin B (PPP3R1) promoter region and its association with left ventricular hypertrophy. Am. Heart J. 2005, 150, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Ahmetov, I.I.; Linde, E.V.; Popov, D.V.; Shihova, J.V.; Missina, S.S.; Vinogradova, O.L.; Rogozkin, V.A. The influence of calcineurin gene polymorphism on morphofunctional characteristics of cardiovascular system of athletes. Rus. J. Physiol. 2008, 94, 915–922. [Google Scholar]
- Mosteller, R.D. Simplified calculation of body-surface area. N. Engl. J. Med. 1987, 317, 1098. [Google Scholar] [PubMed]
- Huwez, F.U.; Houston, A.B.; Watson, J.; McLaughlin, S.; Macfarlane, P.W. Age and body surface area related normal upper and lower limits of M mode echocardiographic measurements and left ventricular volume and mass from infancy to early adulthood. Br. Heart J. 1994, 72, 276–280. [Google Scholar] [CrossRef]
- Gorący, I.; Gorący, J.; Skonieczna-Żydecka, K.; Kaczmarczyk, M.; Dawid, G.; Ciechanowicz, A. Association of genetic variation in calmodulin and left ventricular mass in full-term newborns. Int. J. Genom. 2013, 2013, 410407. [Google Scholar] [CrossRef]
- Han, T.; Wang, X.; Cui, Y.; Ye, H.; Tong, X.; Piao, M. Relationship between angiotensin-converting enzyme gene insertion or deletion polymorphism and insulin sensitivity in healthy newborns. Pediatrics 2007, 19, 1089–1094. [Google Scholar] [CrossRef]
- Gorący, I.; Safranow, K.; Dawid, G.; Skonieczna-Żydecka, K.; Kaczmarczyk, M.; Gorący, J.; Loniewska, B.; Ciechanowicz, A. Common genetic variants of the BMP4, BMPR1A, BMPR1B, and ACVR1 genes, left ventricular mass, and other parameters of the heart in newborns. Genet. Test. Mol. Biomark. 2012, 16, 1309–1316. [Google Scholar] [CrossRef]
- Gorący, I.; Dawid, G.; Łoniewska, B.; Gorący, J.; Ciechanowicz, A. Genetics of the renin-angiotensin system with respect to cardiac and blood pressure phenotypes in healthy newborn infants. J. Renin Angiotensin Aldosterone Syst. 2013, 14, 337–347. [Google Scholar] [CrossRef]
- Gorący, I.; Dawid, G.; Skonieczna-Żydecka, K.; Kaczmarczyk, M.; Łoniewska, B.; Gorący, J. Association of genetic variation in the natriuretic peptide system and left ventricular mass and blood pressure in newborns. Kardiol. Pol. 2015, 73, 366–372. [Google Scholar] [CrossRef]
- Ploski, R.; Wozniak, M.; Pawlowski, R.; Monies, D.M.; Branicki, W.; Kupiec, T.; Kloosterman, A.; Dobosz, T.; Bosch, E.; Nowak, M.; et al. Homogeneity and distinctiveness of Polish paternal lineages revealed by Y chromosome microsatellite haplotype analysis. Human Genet. 2002, 110, 592–600. [Google Scholar] [CrossRef]
- Kayser, M.; Lao, O.; Anslinger, K.; Augustin, C.; Bargel, G.; Edelmann, J.; Elias, S.; Heinrich, M.; Henke, J.; Henke, L.; et al. Significant genetic differentiation between Poland and Germany follows present-day political borders, as revealed by Y-chromosome analysis. Human Genet. 2005, 117, 428–443. [Google Scholar] [CrossRef]
- Debniak, T.; Scott, R.J.; Huzarski, T.; Byrski, T.; Rozmiarek, A.; Debniak, B.; Załuga, E.; Maleszka, R.; Kładny, J.; Górski, B.; et al. CDKN2A common variants and their association with melanoma risk: A population-based study. Cancer Res. 2005, 65, 835–839. [Google Scholar] [CrossRef]
- Adler, G.; Łoniewska, B.; Parczewski, M.; Kordek, A.; Ciechanowicz, A. Frequency of common CYP3A5 gene variants in healthy Polish newborn infants. Pharmacol. Rep. 2009, 61, 947–951. [Google Scholar] [CrossRef]
- Hand, B.D.; Kostek, M.C.; Ferrell, R.E.; Delmonico, M.J.; Douglass, L.W.; Roth, S.M.; Hagberg, J.M.; Hurley, B.F. Influence of promoter region variants of insulin-like growth factor pathway genes on the strength-training response of muscle phenotypes in older adults. J. Appl. Physiol. 2007, 103, 1678–1687. [Google Scholar] [CrossRef]
- Pitoulis, F.G.; Terracciano, C.M. Heart Plasticity in Response to Pressure- and Volume-Overload: A Review of Findings in Compensated and Decompensated Phenotypes. Front. Physiol. 2020, 11, 92. [Google Scholar] [CrossRef]
- Ritchie, J.A.; Ng, J.Q.; Kemi, O.J. When one says yes and the other says no; does calcineurin participate in physiologic cardiac hypertrophy? Adv. Physiol. Educ. 2022, 46, 84–95. [Google Scholar] [CrossRef]
- Bär, H.; Kreuzer, J.; Cojoc, A.; Jahn, L. Upregulation of embryonic transcription factors in right ventricular hypertrophy. Basic Res. Cardiol. 2003, 98, 285–294. [Google Scholar] [CrossRef]
- Nishida, W.; Nakamura, M.; Mori, S.; Takahashi, M.; Ohkawa, Y.; Tadokoro, S.; Yoshida, K.; Hiwada, K.; Hayashi, K.; Sobue, K. A triad of serum response factor and the GATA and NK families governs the transcription of smooth and cardiac muscle genes. J. Biol. Chem. 2002, 277, 7308–7317. [Google Scholar] [CrossRef]
- Schott, J.J.; Benson, D.W.; Basson, C.T.; Pease, W.; Silberbach, G.M.; Moak, J.P.; Maron, B.J.; Seidman, C.E.; Seidman, J.G. Congenital heart disease caused by mutations in the transcription factor NKX2-5. Science 1998, 281, 108–111. [Google Scholar] [CrossRef]
- Benson, D.W.; Silberbach, G.M.; Kavanaugh-McHugh, A.; Cottrill, C.; Zhang, Y.; Riggs, S.; Smalls, O.; Johnson, M.C.; Watson, M.S.; Seidman, J.G.; et al. Mutations in the cardiac transcription factor NKX2.5 affect diverse cardiac developmental pathways. J. Clin. Investig. 1999, 104, 1567–1573. [Google Scholar] [CrossRef]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef] [PubMed]
- Alsheikh, A.J.; Wollenhaupt, S.; King, E.A.; Reeb, J.; Ghosh, S.; Stolzenburg, L.R.; Tamim, S.; Lazar, J.; Davis, J.W.; Jacob, H.J. The landscape of GWAS validation; systematic review identifying 309 validated non-coding variants across 130 human diseases. BMC Med. Genom. 2022, 15, 74. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Calcineurin Subunit Abbreviation (Recommended Name: Human) UNIPROT References: Human/Mouse | Human Gene (locus) Mouse Homolog | Cardiovascular Phenotypes [Reference] |
|---|---|---|
| CnAα (Protein phosphatase 3 catalytic subunit alpha) Q08209/P63328 | PPP3CA (4: 101,023,409–101,348,278) Ppp3ca | Association of rs3017892, rs1441433, rs12116679 and rs1527351 with blood pressure [10,11,12]. Cardiac hypertrophy and development of heart failure within the first few weeks of age in transgenic mice overexpressing a constitutively active form of CnAα [13]. |
| CnAβ (Serine/threonine-protein phosphatase 2B catalytic subunit beta isoform) P16298/P48453 | PPP3CB (10: 73,436,433–73,496,024) Ppp3cb | Association of 12,644 with systolic blood pressure [11]. A 80% decrease in calcineurin enzymatic activity associated with a 12% reduction in basal heart size in CnAβ-deficient mice [14]. No induction of cardiac hypertrophy but improvement of cardiac function and reduction of scar formation after myocardial infarction in transgenic mice with overexpression of CnAβ1 isoform (splicing variant of CnAβ gene with unique C-terminal region) in postnatal cardiomyocytes [15]. |
| CnAγ (Serine/threonine-protein phosphatase 2B catalytic subunit gamma isoform) P48454/P48455 | PPP3CC (8: 22,440,819–22,541,142) Ppp3cc | Association of 73,227,871 with SBP [16]. |
| CnB1 (Calcineurin subunit B type 1) P63098/Q63810 | PPP3R1 (2: 68,178,857–68,256,237) Ppp3r1 | Association of rs6731373 with SBP and with pulse pressure [17]. Lethality at 1 day after birth due to altered right ventricular morphogenesis, reduced ventricular trabeculation, septal defects and valvular overgrowth in mice with the cardiomyocyte-specific deletion of CnB1 in embryonic development. Lethality in early mid adulthood with reduction in cardiac contractility, severe arrhythmia and reduced cardiomyocyte content in mice with postnatal-deletion of CnB1 [18]. Lethal cardiomyopathy, partially characterized by impairment of postnatal growth of cardiac-specific CnB1-deficient mice [19]. |
| CnB2 (Protein phosphatase 3 regulatory subunit B, beta) | PPP3R2 (9: 101,591,604–101,595,021) Ppp3r2 | No data. |
| Variable | Female Newborns (n = 71) | Male Newborns (n = 91) | p |
|---|---|---|---|
| Median (Minimum–Maximum) | |||
| BM [kg] | 3.32 (2.64–4.30) | 3.56 (2.65–5.09) | 0.014 |
| BL [m] | 0.55 (0.51–0.63) | 0.56 (0.47–0.63) | 0.311 |
| BSA [m2] | 0.230 (0.195–0.270) | 0.234 (0.199–0.289) | 0.037 |
| IVST [mm] | 3.9 (2.4–6.3) | 3.7 (2.4–5.1) | 0.139 |
| LVPWT [mm] | 2.7 (1.1–5.8) | 2.8 (2.0–4.0) | 0.307 |
| LVID [mm] | 18.3 (16.0–25.0) | 18.1 (15.0–23.2) | 0.402 |
| LVM [g] | 9.4 (4.9–21.9) | 9.7 (5.4–17.6) | 0.552 |
| LVM/BM [g/kg] | 2.8 (1.5–6.4) | 2.9 (1.8–5.6) | 0.962 |
| LVM/BL [g/m] | 16.8 (9.0–35.9) | 17.8 (10.0–32.1) | 0.790 |
| LVM/BSA [g/m2] | 53.3 (29.0–116.0) | 55.9 (34.5–106.3) | 0.869 |
| Variable | 5I/5I (n = 135) | 5I/5D + 5D/5D (n = 26 + 1) | p |
|---|---|---|---|
| Median, Minimum–Maximum | |||
| BM [kg] | 3.37, 2.64–4.48 | 3.47, 2.84–5.09 | 0.432 |
| BL [m] | 0.56, 0.47–0.63 | 0.56, 0.50–0.60 | 0.572 |
| BSA [m2] | 0.230, 0.195–0.276 | 0.232, 0.203–0.289 | 0.643 |
| IVST [mm] | 3.8, 2.4–6.1 | 3.8, 2.8–6.3 | 0.955 |
| LVPWT [mm] | 2.8, 1.1–5.8 | 3.0, 1.6–5.0 | 0.317 |
| LVID [mm] | 18.4, 15.0–25.0 | 18.1, 15.0–22.0 | 0.579 |
| LVM [g] | 9.7, 4.9–21.9 | 9.4, 6.0–19.5 | 0.946 |
| LVM/BM [g/kg] | 2.8, 1.5–6.4 | 3.0, 1.9–5.2 | 0.590 |
| LVM/BL [g/m] | 17.4, 9.0–35.9 | 18.3, 11.0–33.3 | 0.794 |
| LVM/BSA [g/m2] | 42.0, 22.3–90.6 | 44.7, 27.9–78.8 | 0.650 |
| Variable | Median | Group | 5I/5I n (%) | 5I/5D + 5D/5D n (%) | p |
|---|---|---|---|---|---|
| <2.8 | 68 (83.9) | 13 (16.1) | |||
| LVM/BM [g/kg] | 2.8 | 0.833 | |||
| >2.8 | 67 (82.7) | 14 (17.3) | |||
| <17.5 | 69 (85.2) | 12 (14.8) | |||
| LVM/BL [g/m] | 17.5 | 0.527 | |||
| >17.5 | 66 (81.5) | 15 (18.5) | |||
| <42.2 | 68 (83.9) | 13 (16.1) | |||
| LVM/BSA [g/m2] | 42.2 | 0.833 | |||
| >42.2 | 67 (82.7) | 14 (17.3) |
| Variable | Tertile | 5I/5I n (%) | 5I/5D + 5D/5D n (%) | p |
|---|---|---|---|---|
| Lower: ≤2.6 | 49 (90.7) | 5 (9.3) | ||
| LVM/BM [g/kg] | 0.007 | |||
| Upper: ≥3.1 | 38 (70.4) | 16 (29.6) | ||
| Lower: ≤15.5 | 47 (87.0) | 7 (13.0) | ||
| LVM/BL [g/m] | 0.056 | |||
| Upper: ≥19.4 | 39 (72.2) | 15 (27.8) | ||
| Lower: ≤38.4 | 48 (88.9) | 6 (11.1) | ||
| LVM/BSA [g/m2] | 0.029 | |||
| Upper: ≥45.8 | 39 (72.2) | 15 (27.8) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorący, I.; Łoniewska, B.; Lewandowska, K.; Boroń, A.; Grzegorczyk, M.; Nowak, R.; Clark, J.S.C.; Ciechanowicz, A. Association of the rs3039851 Insertion/Deletion in the Gene PPP3R1, Which Encodes the Regulatory Calcineurin Subunit B Type 1, with Left Ventricular Mass in Polish Full-Term Newborns. Biomedicines 2023, 11, 1415. https://doi.org/10.3390/biomedicines11051415
Gorący I, Łoniewska B, Lewandowska K, Boroń A, Grzegorczyk M, Nowak R, Clark JSC, Ciechanowicz A. Association of the rs3039851 Insertion/Deletion in the Gene PPP3R1, Which Encodes the Regulatory Calcineurin Subunit B Type 1, with Left Ventricular Mass in Polish Full-Term Newborns. Biomedicines. 2023; 11(5):1415. https://doi.org/10.3390/biomedicines11051415
Chicago/Turabian StyleGorący, Iwona, Beata Łoniewska, Klaudyna Lewandowska, Agnieszka Boroń, Małgorzata Grzegorczyk, Robert Nowak, Jeremy Simon C. Clark, and Andrzej Ciechanowicz. 2023. "Association of the rs3039851 Insertion/Deletion in the Gene PPP3R1, Which Encodes the Regulatory Calcineurin Subunit B Type 1, with Left Ventricular Mass in Polish Full-Term Newborns" Biomedicines 11, no. 5: 1415. https://doi.org/10.3390/biomedicines11051415