
The Role of Thrombomodulin in Estrogen-Receptor-Positive Breast Cancer Progression, Metastasis, and Curcumin Sensitivity
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Materials
2.1. Cell Culture and Chemicals
2.2. Public Breast Cancer Datasets Analysis
2.3. Electroporation-Based Transfer of shRNA
2.4. Transfection by Lipofectamine
2.5. Cell Cycle Determination
2.6. TUNEL Assay
2.7. Western Blot Analysis
2.8. Transwell Assay
2.9. xCELLigence Real-Time Cellular Analysis (RTCA) and SRB Assay
2.10. DAPI Staining
2.11. Reverse Transcription-Quantitative (RT-qPCR)
2.12. Statistical Analysis
3. Results
3.1. Higher TM Reflects Better Outcome in Breast Cancer Patients
3.2. Silencing TM Promote Proliferation, Migration, and Invasion in MCF7 Cells
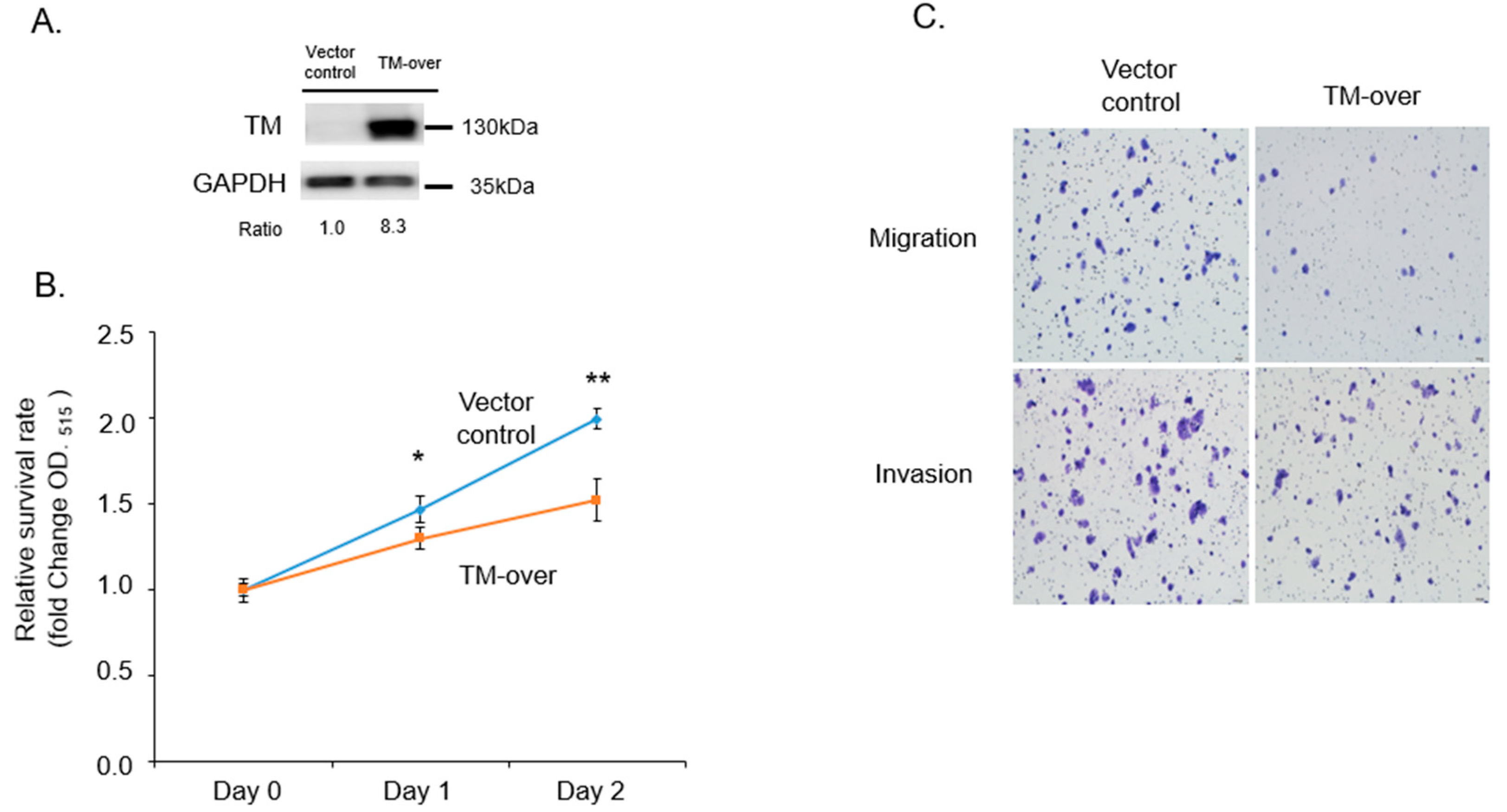
3.3. Overexpression of TM Reduce Proliferation, Migration, and Invasion in T47D Cells
3.4. TM Mediates the Sensitivity of MCF7 and T47D Cells to Curcumin Treatment
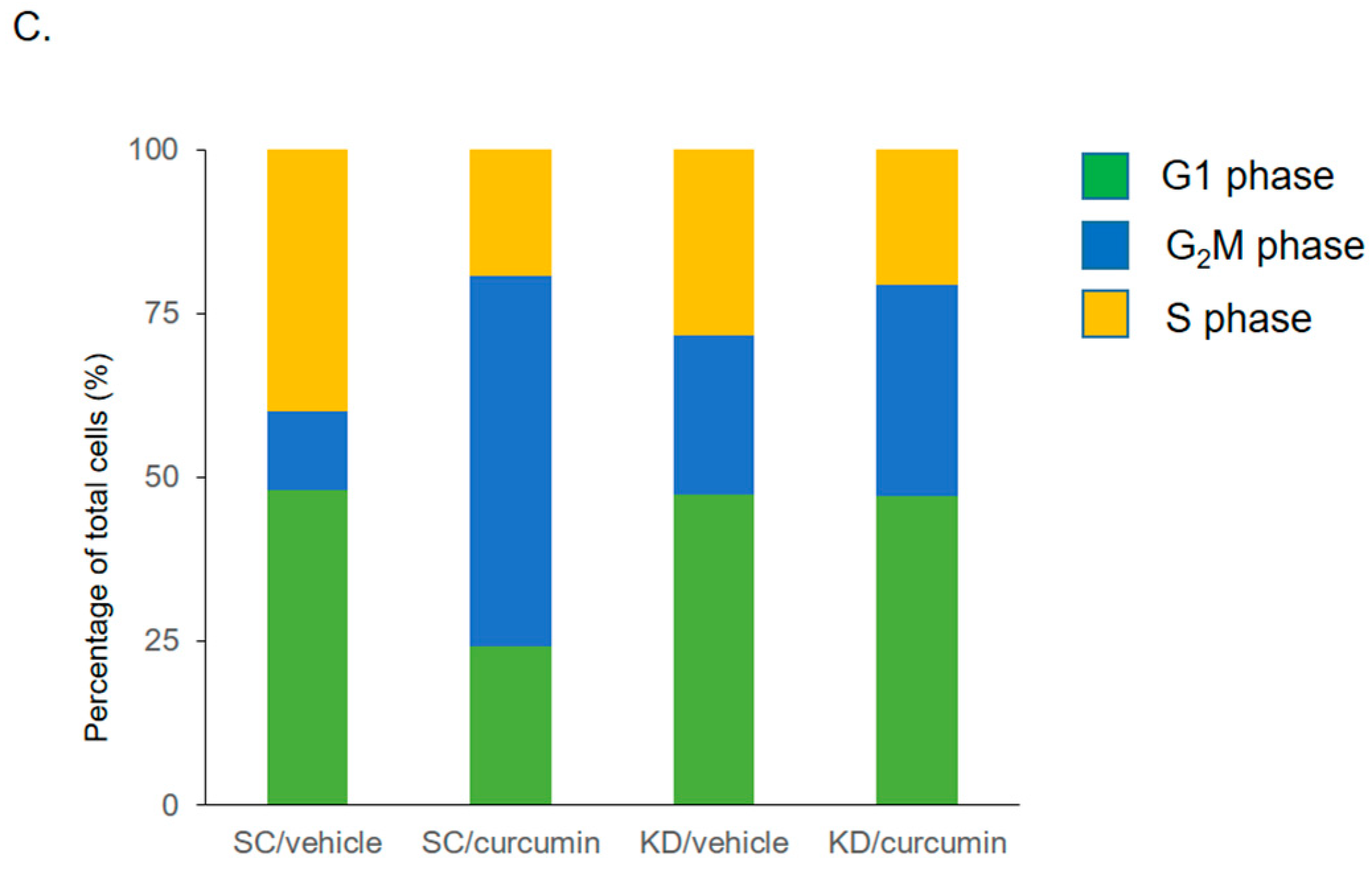
3.5. Knockdown of TM Alters Cell Cycle Distribution after Curcumin Treatment
3.6. TM-KD Cells Are More Susceptible to the Curcumin-Induced Apoptosis
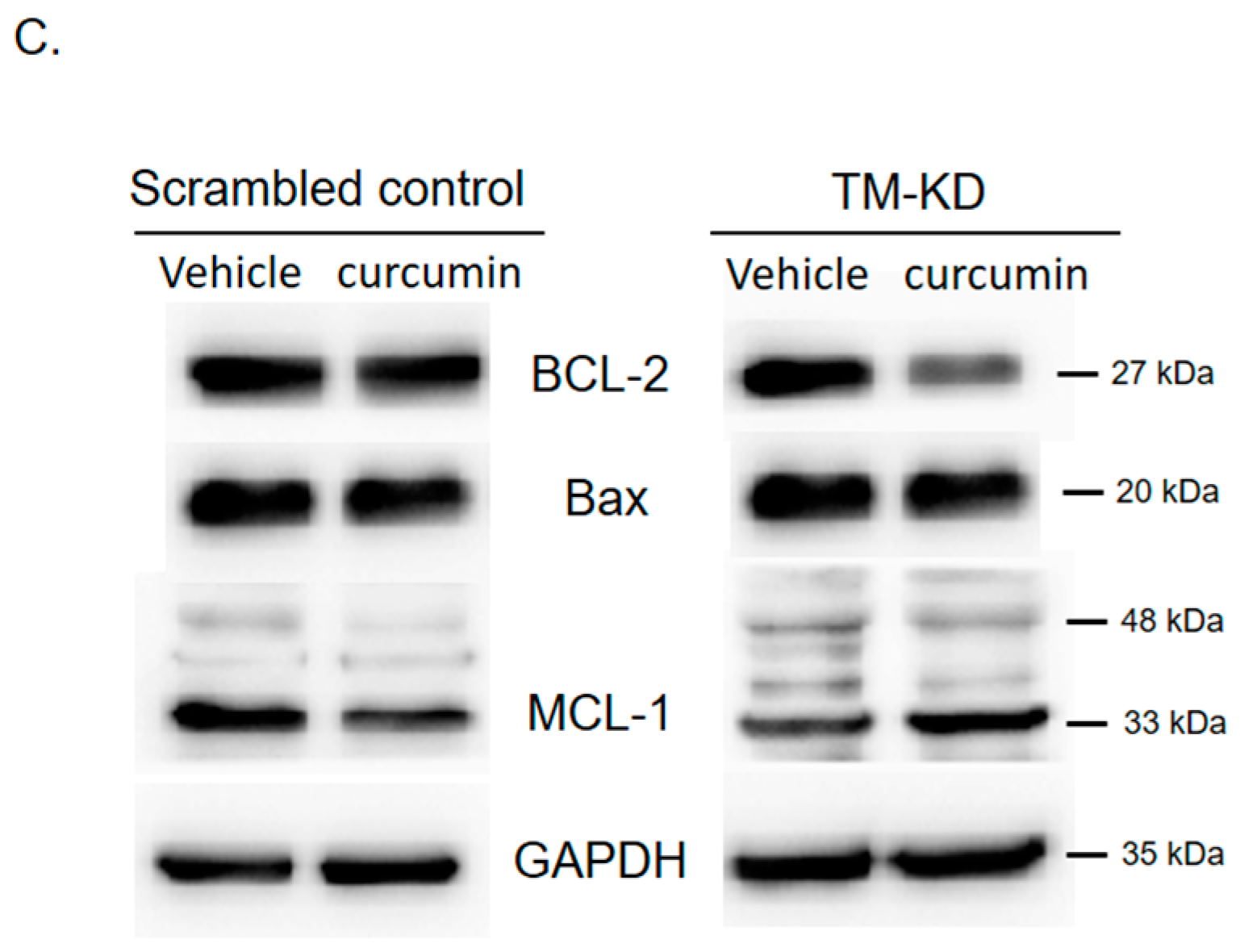
3.7. Silencing TM Sensitizes MCF7 Cells to the Curcumin Treatment through Regulating Drug-Resistant Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, C.T.; Chen, F.M.; Hsieh, H.M. Effect of a national population-based breast cancer screening policy on participation in mammography and stage at breast cancer diagnosis in Taiwan. Health Policy 2020, 124, 478–485. [Google Scholar] [CrossRef]
- Youn, H.J.; Han, W. A Review of the Epidemiology of Breast Cancer in Asia: Focus on Risk Factors. Asian Pac. J. Cancer Prev. 2020, 21, 867–880. [Google Scholar] [CrossRef]
- Godone, R.L.N.; Leitao, G.M.; Araujo, N.B.; Castelletti, C.H.M.; Lima-Filho, J.L.; Martins, D.B.G. Clinical and molecular aspects of breast cancer: Targets and therapies. Biomed. Pharmacother. 2018, 106, 14–34. [Google Scholar] [CrossRef]
- Barzaman, K.; Karami, J.; Zarei, Z.; Hosseinzadeh, A.; Kazemi, M.H.; Moradi-Kalbolandi, S.; Safari, E.; Farahmand, L. Breast cancer: Biology, biomarkers, and treatments. Int. Immunopharmacol. 2020, 84, 106535. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.C.; Hammond, M.E.; Hicks, D.G.; Dowsett, M.; McShane, L.M.; Allison, K.H.; Allred, D.C.; Bartlett, J.M.S.; Bilous, M.; Fritzgibbons, P.; et al. Recommendations for human epidermal growth factor receptor 2 testing in breast cancer: American Society of Clinical Oncology/College of American Pathologists clinical practice guideline update. Arch. Pathol. Lab. Med. 2014, 138, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Mangini, N.S.; Wesolowski, R.; Ramaswamy, B.; Lustberg, M.B.; Berger, M.J. Palbociclib: A Novel Cyclin-Dependent Kinase Inhibitor for Hormone Receptor-Positive Advanced Breast Cancer. Ann. Pharmacother. 2015, 49, 1252–1260. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xu, D.; Li, X.; Zhang, J.; Xu, W.; Hou, J.; Zhang, W.; Tang, J. Latest Overview of the Cyclin-Dependent Kinases 4/6 Inhibitors in Breast Cancer: The Past, the Present and the Future. J. Cancer. 2019, 10, 6608–6617. [Google Scholar] [CrossRef]
- Reinert, T.; Barrios, C.H. Optimal management of hormone receptor positive metastatic breast cancer in 2016. Ther. Adv. Med. Oncol. 2015, 7, 304–320. [Google Scholar] [CrossRef]
- Cardoso, F.; Bischoff, J.; Brain, E.; Zotano, A.G.; Luck, H.J.; Tjan-Heijnen, V.C.; Tanner, M.; Aapro, M. A review of the treatment of endocrine responsive metastatic breast cancer in postmenopausal women. Cancer Treat. Rev. 2013, 39, 457–465. [Google Scholar] [CrossRef]
- Esmon, C.T. The regulation of natural anticoagulant pathways. Science 1987, 235, 1348–1352. [Google Scholar] [CrossRef]
- Pindon, A.; Hantai, D.; Jandrot-Perrus, M.; Festoff, B.W. Novel expression and localization of active thrombomodulin on the surface of mouse brain astrocytes. Glia 1997, 19, 259–268. [Google Scholar] [CrossRef]
- McCachren, S.S.; Diggs, J.; Weinberg, J.B.; Dittman, W.A. Thrombomodulin expression by human blood monocytes and by human synovial tissue lining macrophages. Blood 1991, 78, 3128–3132. [Google Scholar] [CrossRef]
- Polster, B.J.; Westaway, S.K.; Nguyen, T.M.; Yoon, M.Y.; Hayflick, S.J. Discordant expression of miR-103/7 and pantothenate kinase host genes in mouse. Mol. Genet. Metab. 2010, 101, 292–295. [Google Scholar] [CrossRef]
- Kim, S.J.; Shiba, E.; Ishii, H.; Inoue, T.; Taguchi, T.; Tanji, Y.; Kimoto, Y.; Izukura, M.; Takai, S. Thrombomodulin is a new biological and prognostic marker for breast cancer: An immunohistochemical study. Anticancer. Res. 1997, 17, 2319–2323. [Google Scholar] [PubMed]
- Lindahl, A.K.; Boffa, M.C.; Abildgaard, U. Increased plasma thrombomodulin in cancer patients. Thromb. Haemost. 1993, 69, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.; Zhao, F.; Yu, P.; Zhang, X.; Xiao, H.; Qiang, W.; Zhu, H.; Zhao, L. The significance of coagulation and fibrinolysis-related parameters in predicting postoperative venous thrombosis in patients with breast cancer. Gland. Surg. 2021, 10, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Weiler-Guettler, H.; Chen, J.; Wilhelm, O.; Deng, Y.; Qiu, F.; Nakagawa, K.; Klevesath, M.; Wilhelm, S.; Böhrer, H.; et al. Thrombomodulin modulates growth of tumor cells independent of its anticoagulant activity. J. Clin. Invest. 1998, 101, 1301–1309. [Google Scholar] [CrossRef]
- Horowitz, N.A.; Blevins, E.A.; Miller, W.M.; Perry, A.R.; Talmage, K.E.; Mullins, E.S.; Flick, M.J.; Queiroz, K.C.S.; Spek, A.; Conway, E.M.; et al. Thrombomodulin is a determinant of metastasis through a mechanism linked to the thrombin binding domain but not the lectin-like domain. Blood 2011, 118, 2889–2895. [Google Scholar] [CrossRef]
- Kawamoto, E.; Nago, N.; Okamoto, T.; Gaowa, A.; Masui-Ito, A.; Akama, Y.; Darkwah, S.; Appiah, M.G.; Myint, P.K.; Obeng, G.; et al. The Lectin-Like Domain of Thrombomodulin Inhibits beta1 Integrin-Dependent Binding of Human Breast Cancer-Derived Cell Lines to Fibronectin. Biomedicines 2021, 9, 162. [Google Scholar] [CrossRef]
- Ella-Tongwiis, P.; Lamb, R.M.; Makanga, A.; Shergill, I.; Hughes, S.F. The role of antibody expression and their association with bladder cancer recurrence: A single-centre prospective clinical-pilot study in 35 patients. BMC Urol. 2020, 20, 187. [Google Scholar] [CrossRef]
- Suehiro, T.; Shimada, M.; Matsumata, T.; Taketomi, A.; Yamamoto, K.; Sugimachi, K. Thrombomodulin inhibits intrahepatic spread in human hepatocellular carcinoma. Hepatology 1995, 21, 1285–1290. [Google Scholar] [CrossRef]
- Ishii, H.; Majerus, P.W. Thrombomodulin is present in human plasma and urine. J. Clin. Invest. 1985, 76, 2178–2181. [Google Scholar] [CrossRef]
- Kajioka, H.; Kagawa, S.; Ito, A.; Yoshimoto, M.; Sakamoto, S.; Kikuchi, S.; Kuroda, S.; Yoshida, R.; Umeda, Y.; Noma, K.; et al. Targeting neutrophil extracellular traps with thrombomodulin prevents pancreatic cancer metastasis. Cancer Lett. 2021, 497, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A. Dual effect of curcumin in preventing atherosclerosis: The potential role of pro-oxidant-antioxidant mechanisms. Nat. Prod. Res. 2015, 29, 491–492. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Sundaram, C.; Malani, N.; Ichikawa, H. Curcumin: The Indian solid gold. Adv. Exp. Med. Biol. 2007, 595, 1–75. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Murad, W.; Mubin, S.; Ullah, O.; Rehman, N.U.; Rahman, M.H. Multiple health benefits of curcumin and its therapeutic potential. Environ. Sci. Pollut. Res. Int. 2022, 29, 43732–43744. [Google Scholar] [CrossRef]
- Sultana, S.; Munir, N.; Mahmood, Z.; Riaz, M.; Akram, M.; Rebezov, M.; Kuderinova, N.; Moldabayeva, Z.; Shariati, M.A.; Rauf, A.; et al. Molecular targets for the management of cancer using Curcuma longa Linn. phytoconstituents: A Review. Biomed. Pharmacother. 2021, 135, 111078. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Qu, K.; Liu, J.; Wen, Y.; Duan, B. Metabolomics study on liver of db/db mice treated with curcumin using UPLC-Q-TOF-MS. J. Pharm. Biomed. Anal. 2022, 215, 114771. [Google Scholar] [CrossRef]
- Tian, M.; Zhou, F.; Teng, Z.; Wang, C.; Zhang, X.; Wang, Y.; Li, Y. Curcumin ameliorates lipid metabolic disorder and cognitive dysfunction via the ABCA1 transmembrane transport system in APP/PS1 double transgenic mice. J. Integr. Neurosci. 2021, 20, 895–903. [Google Scholar] [CrossRef]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic roles of curcumin: Lessons learned from clinical trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef]
- Bashang, H.; Tamma, S. The use of curcumin as an effective adjuvant to cancer therapy: A short review. Biotechnol. Appl. Biochem. 2020, 67, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.H.; Huang, C.Y.; Chou, C.W.; Makondi, P.T.; Huang, M.T.; Wei, P.L.; Chang, Y.J. Heat shock protein 27 influences the anti-cancer effect of curcumin in colon cancer cells through ROS production and autophagy activation. Life Sci. 2018, 209, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A.; Mohammadi, A.; Atabati, A.; Rahiman, S.; Tavallaie, S.; Iranshahi, M.; Akhlaghi, S.; Ferns, G.A.A.; Ghayour-Mobarhan, M. Curcuminoids modulate pro-oxidant-antioxidant balance but not the immune response to heat shock protein 27 and oxidized LDL in obese individuals. Phytother. Res. 2013, 27, 1883–1888. [Google Scholar] [CrossRef]
- Panahi, Y.; Alishiri, G.H.; Parvin, S.; Sahebkar, A. Mitigation of Systemic Oxidative Stress by Curcuminoids in Osteoarthritis: Results of a Randomized Controlled Trial. J. Diet. Suppl. 2016, 13, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.H.; Wei, P.L.; Hung, C.S.; Wu, C.T.; Wang, W.; Huang, M.T.; Chang, Y.J. MicroRNA-200a/b influenced the therapeutic effects of curcumin in hepatocellular carcinoma (HCC) cells. Tumour Biol. 2013, 34, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhang, M.; Dai, E.; Luo, Y. Molecular targets of curcumin in breast cancer (Review). Mol. Med. Rep. 2019, 19, 23–29. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, J.; Cui, R.; Lin, J.; Ding, X. Curcumin in Treating Breast Cancer: A Review. J. Lab. Autom. 2016, 21, 723–731. [Google Scholar] [CrossRef]
- Cheng, W.L.; Huang, C.Y.; Tai, C.J.; Chang, Y.J.; Hung, C.S. Maspin Enhances the Anticancer Activity of Curcumin in Hormone-refractory Prostate Cancer Cells. Anticancer. Res. 2018, 38, 863–870. [Google Scholar] [CrossRef]
- Pereira, I.C.; Mascarenhas, I.F.; Capetini, V.C.; Ferreira, P.M.P.; Rogero, M.M.; Torres-Leal, F.L. Cellular reprogramming, chemoresistance, and dietary interventions in breast cancer. Crit. Rev. Oncol. Hematol. 2022, 179, 103796. [Google Scholar] [CrossRef]
- Emran, T.B.; Shahriar, A.; Mahmud, A.R.; Rahman, T.; Abir, M.H.; Siddiquee, M.F.; Ahmed, H.; Rahman, N.; Nainu, F.; Wahyudin, E.; et al. Multidrug Resistance in Cancer: Understanding Molecular Mechanisms, Immunoprevention and Therapeutic Approaches. Front. Oncol. 2022, 12, 891652. [Google Scholar] [CrossRef]
- Gyorffy, B. Survival analysis across the entire transcriptome identifies biomarkers with the highest prognostic power in breast cancer. Comput. Struct. Biotechnol. J. 2021, 19, 4101–4109. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cheng, B.J.; Lu, S. Thrombomodulin regulates doxorubicin sensitivity through epithelial-mesenchymal transition in non-small cell lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 95–101. [Google Scholar]
- Chang, Y.J.; Cheng, Y.W.; Lin, R.K.; Huang, C.C.; Chen, W.T.; Ke, T.W.; Wei, P.L. Thrombomodulin Influences the Survival of Patients with Non-Metastatic Colorectal Cancer through Epithelial-To-Mesenchymal Transition (EMT). PLoS ONE 2016, 11, e0160550. [Google Scholar] [CrossRef]
- Li, G.; Wang, Z.; Chong, T.; Yang, J.; Li, H.; Chen, H. Curcumin enhances the radiosensitivity of renal cancer cells by suppressing NF-kappaB signaling pathway. Biomed. Pharmacother. 2017, 94, 974–981. [Google Scholar] [CrossRef]
- Wang, Q.; Fan, H.; Liu, Y.; Yin, Z.; Cai, H.; Liu, J.; Wang, Z.; Shao, M.; Sun, X.; Diao, J.; et al. Curcumin enhances the radiosensitivity in nasopharyngeal carcinoma cells involving the reversal of differentially expressed long non-coding RNAs. Int. J. Oncol. 2014, 44, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.H.; el-Naggar, A.K.; Priebe, W.; Perez-Soler, R. Cell cycle-dependent cytotoxicity, G2/M phase arrest, and disruption of p34cdc2/cyclin B1 activity induced by doxorubicin in synchronized P388 cells. Mol. Pharmacol. 1996, 49, 832–841. [Google Scholar]
- Flint, A.L.; Hansen, D.W.; Brown, L.D.; Stewart, L.E.; Ortiz, E.; Panda, S.S. Modified Curcumins as Potential Drug Candidates for Breast Cancer: An Overview. Molecules 2022, 27, 8891. [Google Scholar] [CrossRef]
- Tabanelli, R.; Brogi, S.; Calderone, V. Improving Curcumin Bioavailability: Current Strategies and Future Perspectives. Pharmaceutics. 2021, 13, 1715. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhou, X.; He, X.; Dai, M.; Zhang, Q. Curcumin induces apoptosis involving bax/bcl-2 in human hepatoma SMMC-7721 cells. Asian Pac. J. Cancer Prev. 2011, 12, 1925–1929. [Google Scholar]
- Elyaman, W.; Terro, F.; Suen, K.C.; Yardin, C.; Chang, R.C.; Hugon, J. BAD and Bcl-2 regulation are early events linking neuronal endoplasmic reticulum stress to mitochondria-mediated apoptosis. Brain Res. Mol. Brain Res. 2002, 109, 233–238. [Google Scholar] [CrossRef]
- Bae, J.; Leo, C.P.; Hsu, S.Y.; Hsueh, A.J. MCL-1S, a splicing variant of the antiapoptotic BCL-2 family member MCL-1, encodes a proapoptotic protein possessing only the BH3 domain. J. Biol. Chem. 2000, 275, 25255–25261. [Google Scholar] [CrossRef]
- Minafra, L.; Porcino, N.; Bravata, V.; Gaglio, D.; Bonanomi, M.; Amore, E.; Cammarata, F.P.; Russo, G.; Militello, C.; Savoca, G.; et al. Radiosensitizing effect of curcumin-loaded lipid nanoparticles in breast cancer cells. Sci. Rep. 2019, 9, 11134. [Google Scholar] [CrossRef] [PubMed]
- Patel, L.R.; Camacho, D.F.; Shiozawa, Y.; Pienta, K.J.; Taichman, R.S. Mechanisms of cancer cell metastasis to the bone: A multistep process. Future Oncol. 2011, 7, 1285–1297. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.H.; Yu, A.M. ABC transporters in multidrug resistance and pharmacokinetics, and strategies for drug development. Curr. Pharm. Des. 2014, 20, 793–807. [Google Scholar] [CrossRef]
- Higgins, C.F. Multiple molecular mechanisms for multidrug resistance transporters. Nature 2007, 446, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, A.; Ierano, C.; Szakacs, G.; Robey, R.W.; Bates, S.E. The controversial role of ABC transporters in clinical oncology. Essays Biochem. 2011, 50, 209–232. [Google Scholar] [CrossRef]
- Kang, H.S.; Kim, J.; Lee, H.J.; Kwon, B.M.; Lee, D.K.; Hong, S.H. LRP1-dependent pepsin clearance induced by 2’-hydroxycinnamaldehyde attenuates breast cancer cell invasion. Int. J. Biochem. Cell. Biol. 2014, 53, 15–23. [Google Scholar] [CrossRef]
- Mantuano, E.; Lam, M.S.; Gonias, S.L. LRP1 assembles unique co-receptor systems to initiate cell signaling in response to tissue-type plasminogen activator and myelin-associated glycoprotein. J. Biol. Chem. 2013, 288, 34009–34018. [Google Scholar] [CrossRef]
- Kasza, A.; Petersen, H.H.; Heegaard, C.W.; Oka, K.; Christensen, A.; Dubin, A.; Chan, L.; Andreasen, P.A. Specificity of serine proteinase/serpin complex binding to very-low-density lipoprotein receptor and alpha2-macroglobulin receptor/low-density-lipoprotein-receptor-related protein. Eur. J. Biochem. 1997, 248, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Xing, P.; Liao, Z.; Ren, Z.; Zhao, J.; Song, F.; Wang, G.; Chen, K.; Yang, J. Roles of low-density lipoprotein receptor-related protein 1 in tumors. Chin. J. Cancer. 2016, 35, 6. [Google Scholar] [CrossRef] [PubMed]
- Roura, S.; Cal, R.; Galvez-Monton, C.; Revuelta-Lopez, E.; Nasarre, L.; Badimon, L.; Bayes-Genis, A.; Llorente-Cortés, V. Inverse relationship between raft LRP1 localization and non-raft ERK1,2/MMP9 activation in idiopathic dilated cardiomyopathy: Potential impact in ventricular remodeling. Int. J. Cardiol. 2014, 176, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Fuentealba, R.A.; Liu, Q.; Kanekiyo, T.; Zhang, J.; Bu, G. Low density lipoprotein receptor-related protein 1 promotes anti-apoptotic signaling in neurons by activating Akt survival pathway. J. Biol. Chem. 2009, 284, 34045–34053. [Google Scholar] [CrossRef]
- Wolking, S.; Schaeffeler, E.; Lerche, H.; Schwab, M.; Nies, A.T. Impact of Genetic Polymorphisms of ABCB1 (MDR1, P-Glycoprotein) on Drug Disposition and Potential Clinical Implications: Update of the Literature. Clin. Pharmacokinet. 2015, 54, 709–735. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.; Fan, Z.Z.; Li, Q. Signal transduction pathways and transcriptional mechanisms of ABCB1/Pgp-mediated multiple drug resistance in human cancer cells. J. Int. Med. Res. 2012, 40, 426–435. [Google Scholar] [CrossRef]
- Breier, A.; Gibalova, L.; Seres, M.; Barancik, M.; Sulova, Z. New insight into p-glycoprotein as a drug target. Anticancer. Agents Med. Chem. 2013, 13, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.P. Multidrug resistance protein 1 (MRP1, ABCC1), a "multitasking" ATP-binding cassette (ABC) transporter. J. Biol. Chem. 2014, 289, 30880–30888. [Google Scholar] [CrossRef]
- Yakusheva, E.N.; Titov, D.S. Structure and Function of Multidrug Resistance Protein 1. Biochemistry 2018, 83, 907–929. [Google Scholar] [CrossRef]
- Rosenberg, M.F.; Mao, Q.; Holzenburg, A.; Ford, R.C.; Deeley, R.G.; Cole, S.P. The structure of the multidrug resistance protein 1 (MRP1/ABCC1). crystallization and single-particle analysis. J. Biol. Chem. 2001, 276, 16076–16082. [Google Scholar] [CrossRef]
- Wang, H.; Jin, G.; Wang, H.; Liu, G.; Qian, J.; Jin, L.; Wei, Q.; Shen, H.; Huang, W.; Lu, D. Genetic susceptibility of lung cancer associated with common variants in the 3’ untranslated regions of the adenosine triphosphate-binding cassette B1 (ABCB1) and ABCC1 candidate transporter genes for carcinogen export. Cancer 2009, 115, 595–607. [Google Scholar] [CrossRef]
- Low, F.G.; Shabir, K.; Brown, J.E.; Bill, R.M.; Rothnie, A.J. Roles of ABCC1 and ABCC4 in Proliferation and Migration of Breast Cancer Cell Lines. Int. J. Mol. Sci. 2020, 21, 7664. [Google Scholar] [CrossRef] [PubMed]
- Behfarjam, F.; Rostamzadeh, J.; Zarei, M.A.; Nikkhoo, B. Association of Two Polymorphic Codons in P53 and ABCC1 Promoter with Prostate Cancer. Iran. J. Biotechnol. 2015, 13, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Emmanouilidi, A.; Casari, I.; Gokcen Akkaya, B.; Maffucci, T.; Furic, L.; Guffanti, F.; Broggini, M.; Chen, X.; Maxuitenko, Y.Y.; Keeton, A.B.; et al. Inhibition of the Lysophosphatidylinositol Transporter ABCC1 Reduces Prostate Cancer Cell Growth and Sensitizes to Chemotherapy. Cancers 2020, 12, 2022. [Google Scholar] [CrossRef] [PubMed]
- Allikmets, R.; Gerrard, B.; Hutchinson, A.; Dean, M. Characterization of the human ABC superfamily: Isolation and mapping of 21 new genes using the expressed sequence tags database. Hum. Mol. Genet. 1996, 5, 1649–1655. [Google Scholar] [CrossRef] [PubMed]
- Oguri, T.; Achiwa, H.; Sato, S.; Bessho, Y.; Takano, Y.; Miyazaki, M.; Muramatsu, H.; Maeda, H.; Niimi, T.; Ueda, R. The determinants of sensitivity and acquired resistance to gemcitabine differ in non-small cell lung cancer: A role of ABCC5 in gemcitabine sensitivity. Mol. Cancer Ther. 2006, 5, 1800–1806. [Google Scholar] [CrossRef]
- Nedeljkovic, M.; Tanic, N.; Prvanovic, M.; Milovanovic, Z.; Tanic, N. Friend or foe: ABCG2, ABCC1 and ABCB1 expression in triple-negative breast cancer. Breast Cancer. 2021, 28, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, V.A.; Da Silva, K.A.E.P.; Delou, J.M.A.; Da Fonseca, L.M.; Lopes, A.G.; Capella, M.A.M. Modulation of ABCC1 and ABCG2 proteins by ouabain in human breast cancer cells. Anticancer Res. 2014, 34, 1441–1448. [Google Scholar] [PubMed]
- Issa, M.E.; Hall, S.R.; Dupuis, S.N.; Graham, C.L.; Jakeman, D.L.; Goralski, K.B. Jadomycins are cytotoxic to ABCB1-, ABCC1-, and ABCG2-overexpressing MCF7 breast cancer cells. Anticancer Drugs 2014, 25, 255–269. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Z.; Gao, S.; Wu, K.; Bai, F.; Zhang, Q.; Wang, H.; Ye, Q.; Xu, F.; Sun, H.; et al. Human drug efflux transporter ABCC5 confers acquired resistance to pemetrexed in breast cancer. Cancer Cell. Int. 2021, 21, 136. [Google Scholar] [CrossRef]
- Mourskaia, A.A.; Amir, E.; Dong, Z.; Tiedemann, K.; Cory, S.; Omeroglu, A.; Bertos, N.; Ouellet, V.; Clemons, M.; Scheffer, G.L.; et al. ABCC5 supports osteoclast formation and promotes breast cancer metastasis to bone. Breast Cancer Res. 2012, 14, R149. [Google Scholar] [CrossRef]
- D’Andrea, F.P. Intrinsic radiation resistance of mesenchymal cancer stem cells and implications for treatment response in a murine sarcoma model. Dan. Med. J. 2012, 59, B4388. [Google Scholar]
- Choi, H.J.; Jhe, Y.L.; Kim, J.; Lim, J.Y.; Lee, J.E.; Shin, M.K.; Cheong, J.H. FoxM1-dependent and fatty acid oxidation-mediated ROS modulation is a cell-intrinsic drug resistance mechanism in cancer stem-like cells. Redox Biol. 2020, 36, 101589. [Google Scholar] [CrossRef] [PubMed]
- Rochat, B. Importance of influx and efflux systems and xenobiotic metabolizing enzymes in intratumoral disposition of anticancer agents. Curr. Cancer Drug. Targets. 2009, 9, 652–674. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.; Parida, S.; Lingipilli, B.T.; Krishnan, R.; Podipireddy, D.R.; Muniraj, N. Role of Gut Microbiota in Breast Cancer and Drug Resistance. Pathogens 2023, 12, 468. [Google Scholar] [CrossRef]
- Terrisse, S.; Derosa, L.; Iebba, V.; Ghiringhelli, F.; Vaz-Luis, I.; Kroemer, G.; Fidelle, M.; Christodoulidis, S.; Segata, N.; Thomas, A.M.; et al. Intestinal microbiota influences clinical outcome and side effects of early breast cancer treatment. Cell. Death Differ. 2021, 28, 2778–2796. [Google Scholar] [CrossRef]
- Hou, M.F.; Ou-Yang, F.; Li, C.L.; Chen, F.M.; Chuang, C.H.; Kan, J.Y.; Wu, C.C.; Shih, S.L.; Shiau, J.P.; Kao, L.C.; et al. Comprehensive profiles and diagnostic value of menopausal-specific gut microbiota in premenopausal breast cancer. Exp. Mol. Med. 2021, 53, 1636–1646. [Google Scholar] [CrossRef] [PubMed]
- Pricci, M.; Girardi, B.; Giorgio, F.; Losurdo, G.; Ierardi, E.; Di Leo, A. Curcumin and Colorectal Cancer: From Basic to Clinical Evidences. Int. J. Mol. Sci. 2020, 21, 2364. [Google Scholar] [CrossRef]
- Pluta, R.; Januszewski, S.; Ulamek-Koziol, M. Mutual Two-Way Interactions of Curcumin and Gut Microbiota. Int. J. Mol. Sci. 2020, 21, 1055. [Google Scholar] [CrossRef]
- McFadden, R.M.; Larmonier, C.B.; Shehab, K.W.; Midura-Kiela, M.; Ramalingam, R.; Harrison, C.A.; Besselsen, D.G.; Chase, J.H.; Caporaso, J.G.; Jobin, C.; et al. The Role of Curcumin in Modulating Colonic Microbiota During Colitis and Colon Cancer Prevention. Inflamm. Bowel Dis. 2015, 21, 2483–2494. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.-Y.; Wei, P.-L.; Prince, G.M.S.H.; Batzorig, U.; Lee, C.-C.; Chang, Y.-J.; Hung, C.-S. The Role of Thrombomodulin in Estrogen-Receptor-Positive Breast Cancer Progression, Metastasis, and Curcumin Sensitivity. Biomedicines 2023, 11, 1384. https://doi.org/10.3390/biomedicines11051384
Huang C-Y, Wei P-L, Prince GMSH, Batzorig U, Lee C-C, Chang Y-J, Hung C-S. The Role of Thrombomodulin in Estrogen-Receptor-Positive Breast Cancer Progression, Metastasis, and Curcumin Sensitivity. Biomedicines. 2023; 11(5):1384. https://doi.org/10.3390/biomedicines11051384
Chicago/Turabian StyleHuang, Chien-Yu, Po-Li Wei, G. M. Shazzad Hossain Prince, Uyanga Batzorig, Cheng-Chin Lee, Yu-Jia Chang, and Chin-Sheng Hung. 2023. "The Role of Thrombomodulin in Estrogen-Receptor-Positive Breast Cancer Progression, Metastasis, and Curcumin Sensitivity" Biomedicines 11, no. 5: 1384. https://doi.org/10.3390/biomedicines11051384