
Low Temperature Delays the Effects of Ischemia in Bergmann Glia and in Cerebellar Tissue Swelling
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cerebellar Slice Preparation
2.2. Tissue Swelling Assay
2.3. Confocal Microscopy and Image Acquisition
2.4. Image Processing
2.5. Electrophysiology
2.6. Ion-Sensitive Microelectrode Recordings
2.7. Data Analysis and Statistics
3. Results
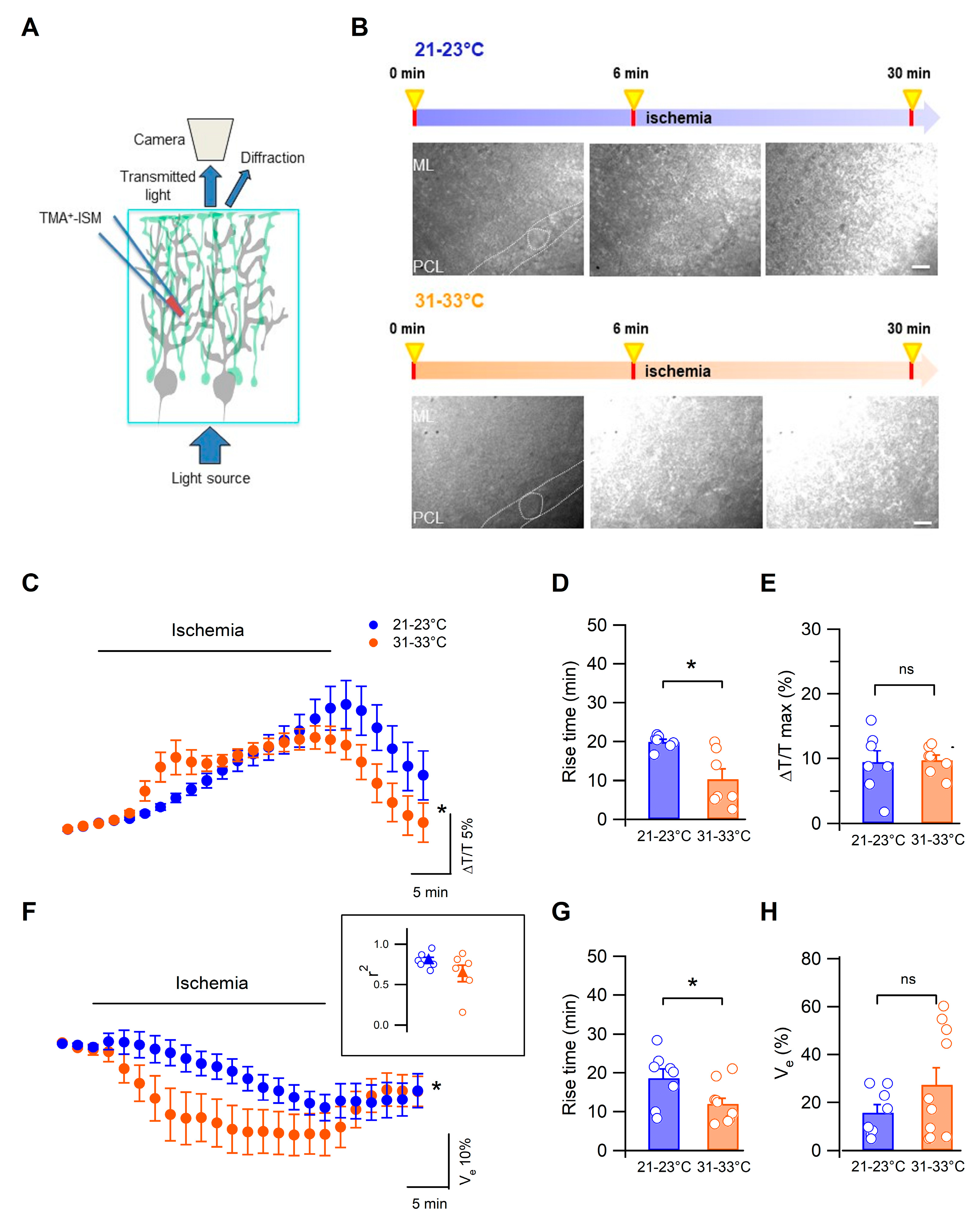
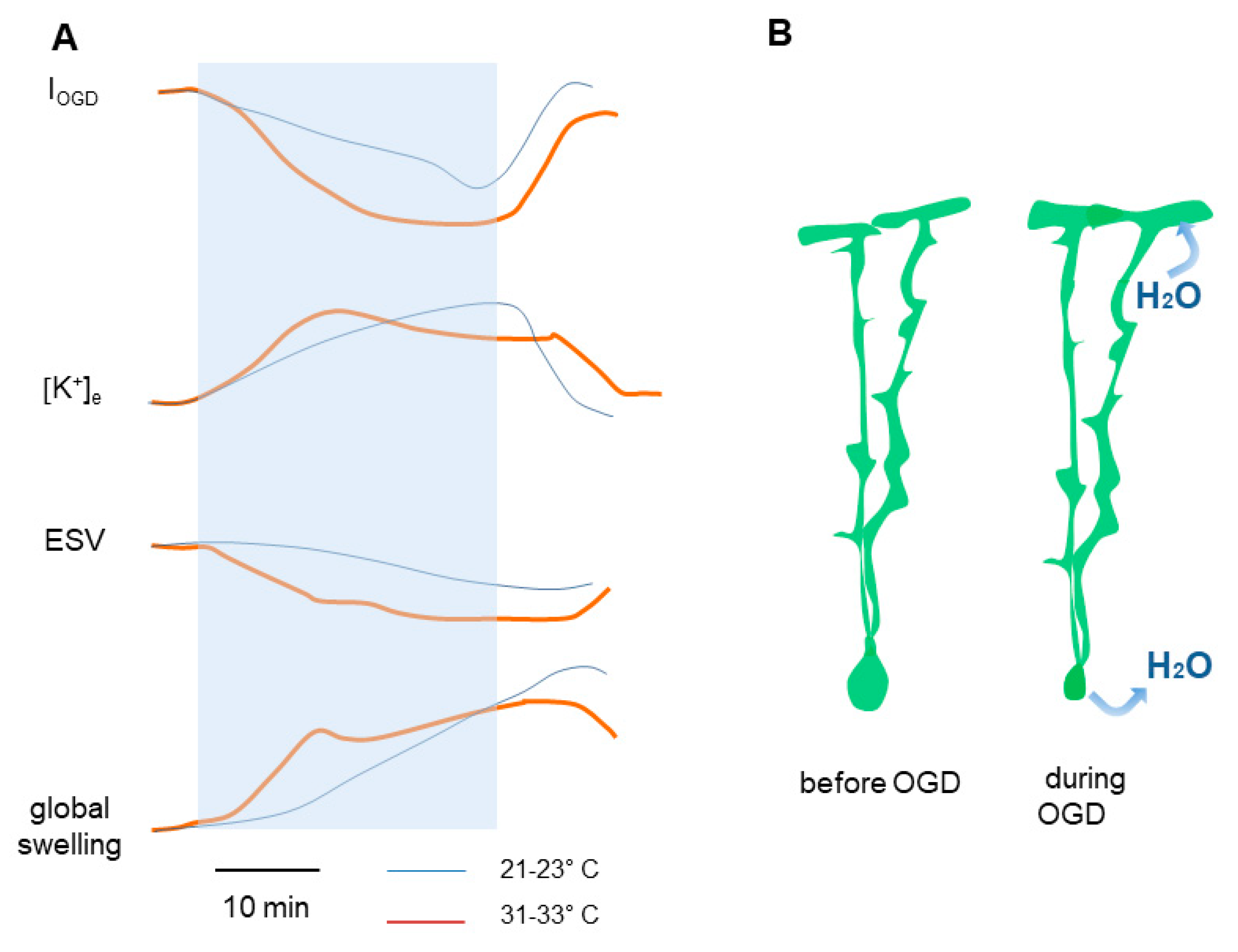
3.1. Low Temperature Delays Cerebellar Tissue Swelling during Ischemia
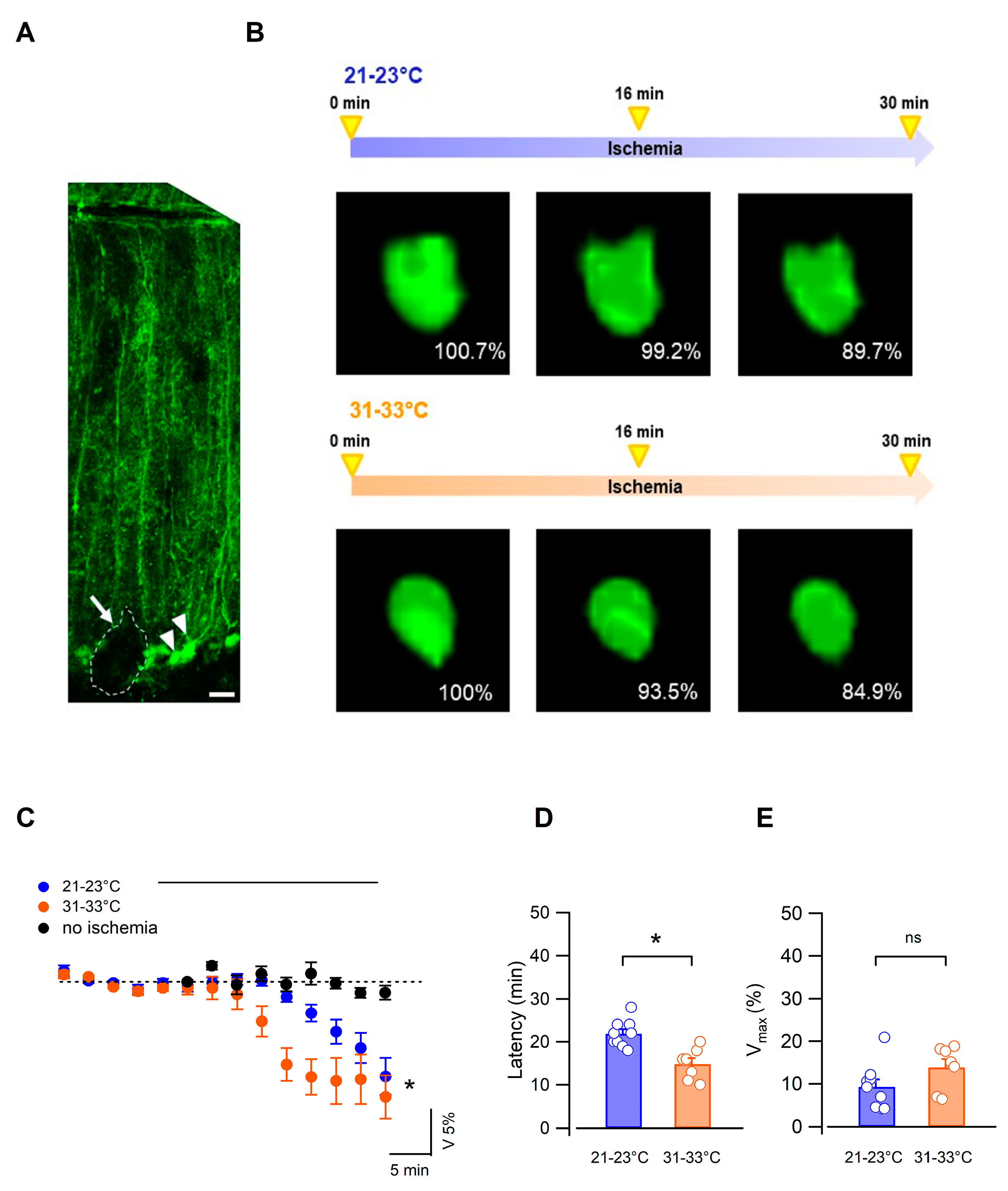
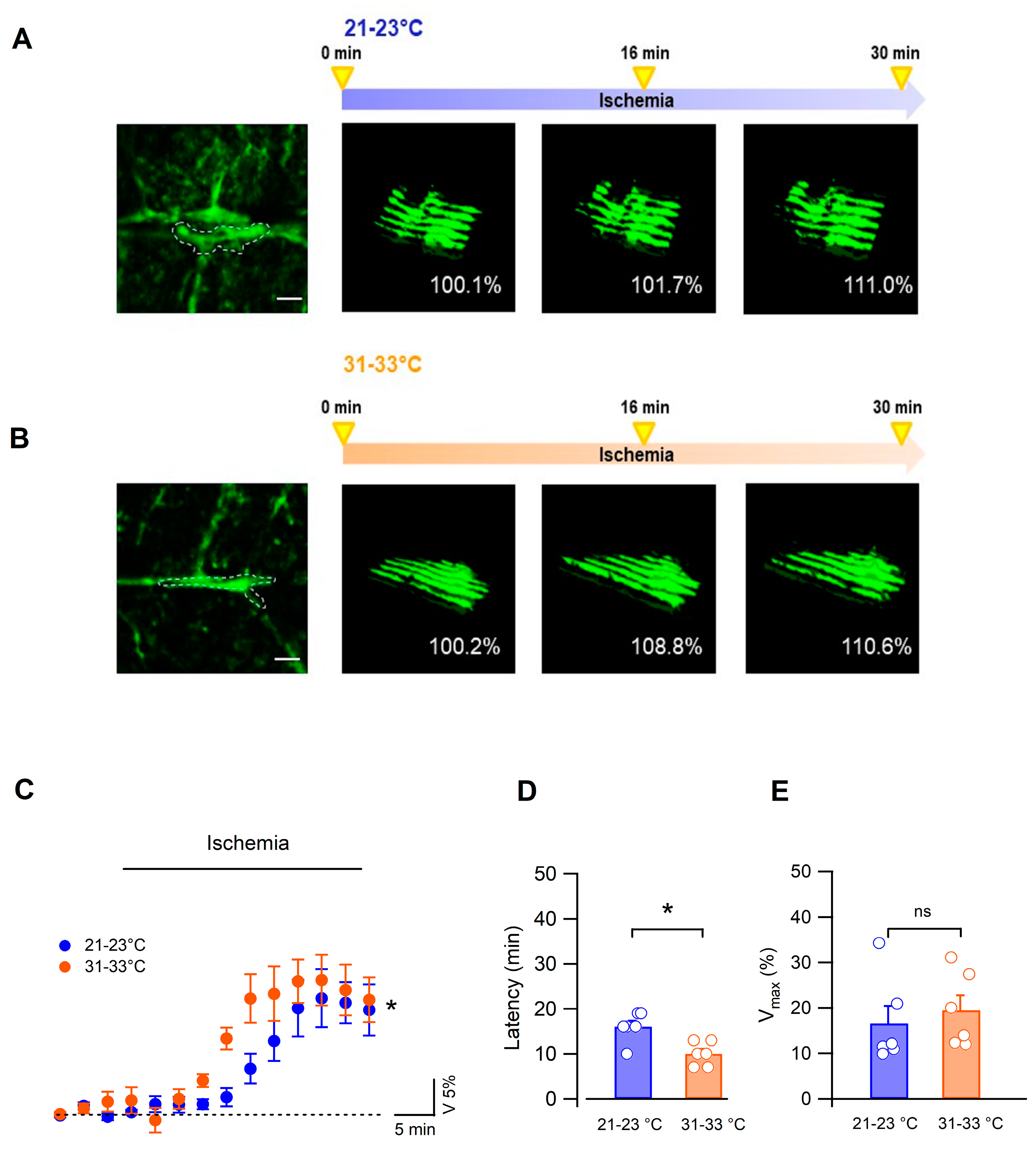
3.2. Bergmann Glial Cells Adapt Their Volume to the Extracellular Environment during OGD
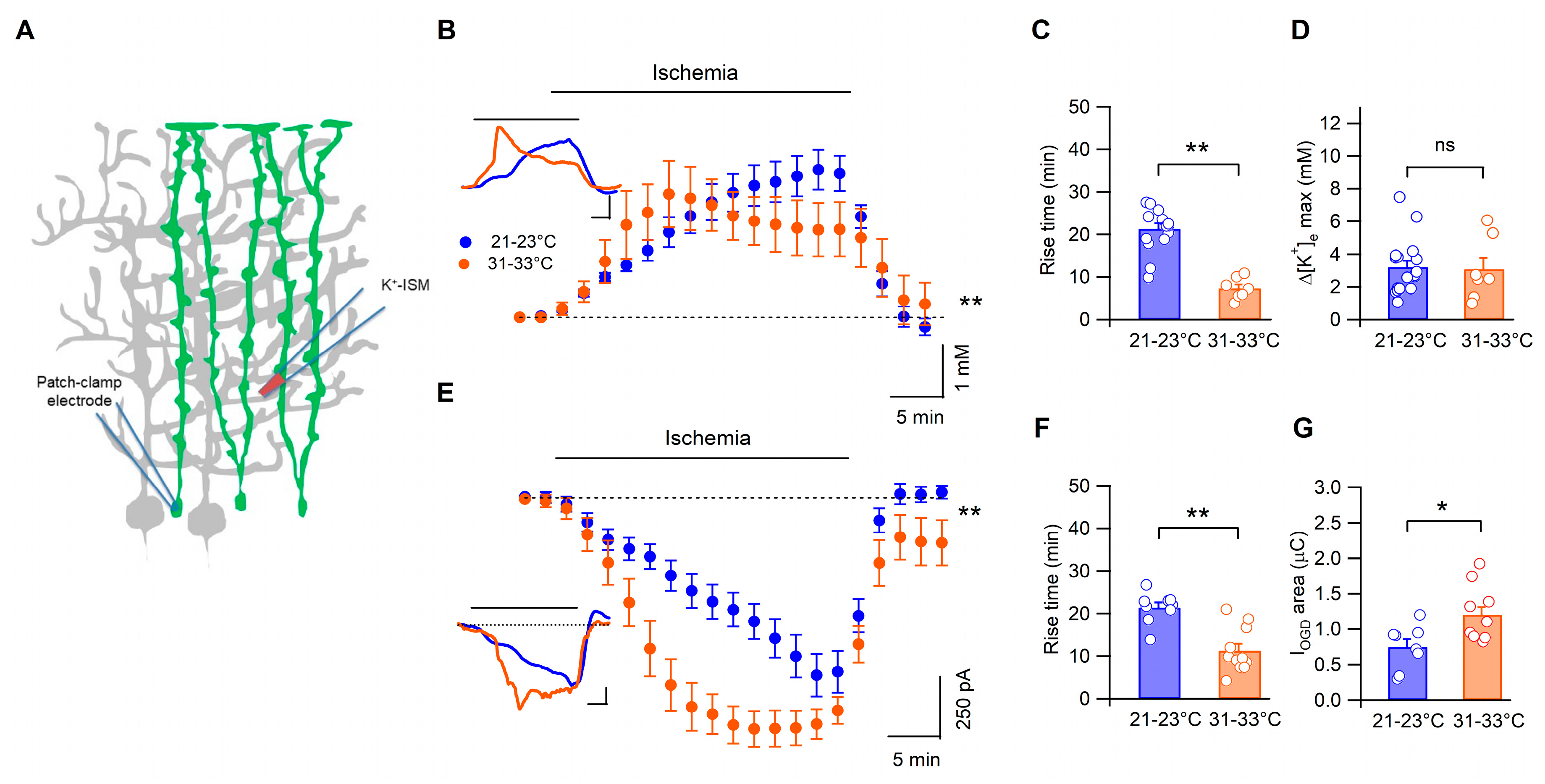
3.3. Temperature Modulates the Kinetics of Bergmann Glia Currents and K+ Accumulation during Ischemia
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stokum, J.A.; Kurland, D.B.; Gerzanich, V.; Simard, J.M. Mechanisms of astrocyte-mediated cerebral edema. Neurochem. Res. 2015, 40, 317–328. [Google Scholar] [CrossRef]
- Gupta, R.; Jovin, T.G.; Krieger, D.W. Therapeutic hypothermia for stroke: Do new outfits change an old friend? Expert Rev. Neurother. 2005, 5, 235–246. [Google Scholar] [CrossRef]
- Zhao, H.; Steinberg, G.K.; Sapolsky, R.M. General versus specific actions of mild-moderate hypothermia in attenuating cerebral ischemic damage. J. Cereb. Blood Flow. Metab. 2007, 27, 1879–1894. [Google Scholar] [CrossRef]
- Jacobs, S.E.; Morley, C.J.; Inder, T.E.; Stewart, M.J.; Smith, K.R.; McNamara, P.J.; Wright, I.M.; Kirpalani, H.M.; Darlow, B.A.; Doyle, L.W. Infant Cooling Evaluation Collaboration. Whole-body hypothermia for term and near-term newborns with hypoxic-ischemic encephalopathy: A randomized controlled trial. Arch. Pediatr. Adolesc. Med. 2011, 165, 692–700. [Google Scholar] [CrossRef]
- Lemyre, B.; Chau, V. Hypothermia for newborns with hypoxic-ischemic encephalopathy. Paediatr. Child Health 2018, 23, 285–291. [Google Scholar] [CrossRef]
- Sinclair, H.L.; Andrews, P.J. Bench-to-bedside review: Hypothermia in traumatic brain injury. Crit. Car 2010, 14, 204. [Google Scholar] [CrossRef]
- van der Worp, H.B.; Sena, E.S.; Donnan, G.A.; Howells, D.W.; Macleod, M.R. Hypothermia in animal models of acute ischaemic stroke: A systematic review and meta-analysis. Brain 2007, 130, 3063–3074. [Google Scholar] [CrossRef]
- Mohr, C.; Brady, J.D.; Rossi, D.J. Young age and low temperature, but not female gender delay ATP loss and glutamate release, and protect Purkinje cells during simulated ischemia in cerebellar slices. Neuropharmacology 2010, 58, 392–403. [Google Scholar] [CrossRef]
- Campos, F.; Pérez-Mato, M.; Agulla, J.; Blanco, M.; Barral, D.; Almeida, A.; Brea, D.; Waeber, C.; Castillo, J.; Ramos-Cabrer, P. Glutamate excitoxicity is the key molecular mechanism which is influenced by body temperature during the acute phase of brain stroke. PLoS ONE 2012, 7, e44191. [Google Scholar] [CrossRef]
- Yenari, M.A.; Han, H.S. Neuroprotective mechanisms of hypothermia in brain ischaemia. Nat. Rev. Neurosci. 2012, 13, 267–278. [Google Scholar] [CrossRef]
- Kimelberg, H.K. Astrocytic swelling in cerebral ischemia as a possible cause of injury and target for therapy. Glia 2005, 50, 389–397. [Google Scholar] [CrossRef]
- Belov Kirdajova, D.; Kriska, J.; Tureckova, J.; Anderova, M. Ischemia-Triggered Glutamate Excitotoxicity From the Perspective of Glial Cells. Front. Cell. Neurosci. 2020, 14, 51. [Google Scholar] [CrossRef]
- Gu, Y.; Zhou, C.; Piao, Z.; Yuan, H.; Jiang, H.; Wei, H.; Zhou, Y.; Nan, G.; Ji, X. Cerebral edema after ischemic stroke: Pathophysiology and underlying mechanisms. Front. Neurosci. 2022, 16, 988283. [Google Scholar] [CrossRef]
- MacAulay, N.; Zeuthen, T. Water transport between CNS compartments: Contributions of aquaporins and cotransporters. Neuroscience 2010, 168, 941–956. [Google Scholar] [CrossRef]
- Dallérac, G.; Chever, O.; Rouach, N. How do astrocytes shape synaptic transmission? Insights from electrophysiology. Front. Cell. Neurosci. 2013, 7, 159. [Google Scholar] [CrossRef]
- Kitaura, H.; Tsujita, M.; Huber, V.J.; Kakita, A.; Shibuki, K.; Sakimura, K.; Kwee, I.L.; Nakada, T. Activity-dependent glial swelling is impaired in aquaporin-4 knockout mice. Neurosci. Res. 2009, 64, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Haj-Yasein, N.N.; Bugge, C.E.; Jensen, V.; Østby, I.; Ottersen, O.P.; Hvalby, Ø.; Nagelhus, E.A. Deletion of aquaporin-4 increases extracellular K+ concentration during synaptic stimulation in mouse hippocampus. Brain Struct. Funct. 2015, 220, 2469–2474. [Google Scholar] [CrossRef] [PubMed]
- Poopalasundaram, S.; Knott, C.; Shamotienko, O.G.; Foran, P.G.; Dolly, J.O.; Ghiani, C.A.; Gallo, V.; Wilkin, G.P. Glial heterogeneity in expression of the inwardly rectifying K+ channel, Kir4.1, in adult rat CNS. Glia 2000, 30, 362–372. [Google Scholar] [CrossRef]
- Higashi, K.; Fujita, A.; Inanobe, A.; Tanemoto, M.; Doi, K.; Kubo, T.; Kurachi, Y. An inwardly rectifying K+ channel, Kir4.1, expressed in astrocytes surrounds synapses and blood vessels in brain. Am. J. Physiol. Cell Physiol. 2001, 281, C922–C931. [Google Scholar] [CrossRef]
- Hirrlinger, P.G.; Wurm, A.; Hirrlinger, J.; Bringmann, A.; Reichenbach, A. Osmotic swelling characteristics of glial cells in the murine hippocampus, cerebellum, and retina in situ. J. Neurochem. 2008, 105, 1405–1417. [Google Scholar] [CrossRef] [PubMed]
- Nicchia, G.P.; Rossi, A.; Nudel, U.; Svelto, M.; Frigeri, A. Dystrophin-dependent and -independent AQP4 pools are expressed in the mouse brain. Glia 2008, 56, 869–876. [Google Scholar] [CrossRef]
- Tanaka, M.; Yamaguchi, K.; Tatsukawa, T.; Theis, M.; Willecke, K.; Itohara, S. Connexin43 and Bergmann glial gap junctions in cerebellar function. Front. Neurosci. 2008, 2, 225–233. [Google Scholar] [CrossRef]
- Nagy, J.I.; Rash, J.E. Cx36, Cx43 and Cx45 in mouse and rat cerebellar cortex: Species-specific expression, compensation in Cx36 null mice and co-localization in neurons vs. glia. Eur. J. Neurosci. 2017, 46, 1790–1804. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, A.; Siegel, A.; Rickmann, M.; Wolff, J.R.; Noone, D.; Robinson, S.R. Distribution of Bergmann glial somata and processes: Implications for function. J. Hirnforsch. 1995, 36, 509–517. [Google Scholar]
- De Zeeuw, C.I.; Hoogland, T.M. Reappraisal of Bergmann glial cells as modulators of cerebellar circuit function. Front. Cell. Neurosci. 2015, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Helleringer, R.; Chever, O.; Daniel, H.; Galante, M. Oxygen and Glucose Deprivation Induces Bergmann Glia Membrane Depolarization and Ca2+ Rises Mainly Mediated by K+ and ATP Increases in the Extracellular Space. Front. Cell. Neurosci. 2017, 11, 349. [Google Scholar] [CrossRef]
- Allen, N.J.; Rossi, D.J.; Attwell, D. Sequential release of GABA by exocytosis and reversed uptake leads to neuronal swelling in simulated ischemia of hippocampal slices. J. Neurosci. 2004, 24, 3837–3849. [Google Scholar] [CrossRef]
- Prakash, Y.S.; Smithson, K.G.; Sieck, G.C. Application of the Cavalieri principle in volume estimation using laser confocal microscopy. Neuroimage 1994, 1, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Kubinová, L.; Janáček, J.; Guilak, F.; Opatrný, Z. Comparison of several digital and stereological methods for estimating surface area and volume of cells studied by confocal microscopy. Cytometry 1999, 36, 85–95. [Google Scholar] [CrossRef]
- Duerstock, B.S.; Bajaj, C.L.; Borgens, R.B. A comparative study of the quantitative accuracy of three-dimensional reconstructions of spinal cord from serial histological sections. J. Microsc. 2003, 210, 138–148. [Google Scholar] [CrossRef]
- Slivka, A.; Murphy, E.; Horrocks, L. Cerebral edema after temporary and permanent middle cerebral artery occlusion in the rat. Stroke 1995, 26, 1061–1065. [Google Scholar] [CrossRef] [PubMed]
- Hamann, M.; Rossi, D.J.; Mohr, C.; Andrade, A.L.; Attwell, D. The electrical response of cerebellar Purkinje neurons to simulated ischaemia. Brain 2005, 128, 2408–2420. [Google Scholar] [CrossRef]
- Brady, J.D.; Mohr, C.; Rossi, D.J. Vesicular GABA release delays the onset of the Purkinje cell terminal depolarization without affecting tissue swelling in cerebellar slices during simulated ischemia. Neuroscience 2010, 168, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Rosic, B.; Dukefoss, D.B.; Åbjørsbråten, K.S.; Tang, W.; Jensen, V.; Ottersen, O.P.; Enger, R.; Nagelhus, E.A. Aquaporin-4-independent volume dynamics of astroglial endfeet during cortical spreading depression. Glia 2019, 67, 1113–1121. [Google Scholar] [CrossRef]
- Nedergaard, M.; Dirnagl, U. Role of glial cells in cerebral ischemia. Glia 2005, 50, 281–286. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.; Guo, W.; Burnstock, G.; He, C.; Xiang, Z. The superficial glia limitans of mouse and monkey brain and spinal cord. Anat. Rec. 2013, 296, 995–1007. [Google Scholar] [CrossRef]
- Nagelhus, E.A.; Mathiisen, T.M.; Ottersen, O.P. Aquaporin-4 in the central nervous system: Cellular and subcellular distribution and coexpression with KIR4.1. Neuroscience 2004, 129, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Amiry-Moghaddam, M.; Frydenlund, D.S.; Ottersen, O.P. Anchoring of aquaporin-4 in brain: Molecular mechanisms and implications for the physiology and pathophysiology of water transport. Neuroscience 2004, 129, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Salmon, M.; Slaoui, L.; Mazaré, N.; Gilbert, A.; Oudart, M.; Alvear-Perez, R.; Elorza-Vidal, X.; Chever, O.; Boulay, A.C. Astrocytes in the regulation of cerebrovascular functions. Glia 2021, 69, 817–841. [Google Scholar] [CrossRef] [PubMed]
- Erecinska, M.; Thoresen, M.; Silver, I.A. Effects of hypothermia on energy metabolism in Mammalian central nervous system. J. Cereb. Blood Flow Metab. 2003, 23, 513–530. [Google Scholar] [CrossRef]
- Xiao, F.; Zhang, S.; Arnold, T.C.; Alexander, J.S.; Huang, J.; Carden, D.L.; Conrad, S.A. Mild hypothermia induced before cardiac arrest reduces brain edema formation in rats. Acad. Emerg. Med. 2002, 9, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Lyden, P.D.; Lamb, J.; Kothari, S.; Toossi, S.; Boitano, P.; Rajput, P.S. Differential effects of hypothermia on neurovascular unit determine protective or toxic results: Toward optimized therapeutic hypothermia. J. Cereb. Blood Flow Metab. 2019, 39, 1693–1709. [Google Scholar] [CrossRef]
- MacAulay, N. Molecular mechanisms of brain water transport. Nat. Rev. Neurosci. 2021, 22, 326–344. [Google Scholar] [CrossRef]
- Amiry-Moghaddam, M.; Xue, R.; Haug, F.M.; Neely, J.D.; Bhardwaj, A.; Agre, P.; Adams, M.E.; Froehner, S.C.; Mori, S.; Ottersen, O.P. Alpha-syntrophin deletion removes the perivascular but not endothelial pool of aquaporin-4 at the blood-brain barrier and delays the development of brain edema in an experimental model of acute hyponatremia. FASEB J. 2004, 18, 542–544. [Google Scholar] [CrossRef]
- Dibaj, P.; Kaiser, M.; Hirrlinger, J.; Kirchhoff, F.; Neusch, C. Kir4.1 channels regulate swelling of astroglial processes in experimental spinal cord edema. J. Neurochem. 2007, 103, 2620–2628. [Google Scholar] [CrossRef] [PubMed]
- Benesova, J.; Hock, M.; Butenko, O.; Prajerova, I.; Anderova, M.; Chvatal, A. Quantification of astrocyte volume changes during ischemia in situ reveals two populations of astrocytes in the cortex of GFAP/EGFP mice. J. Neurosci. Res. 2009, 87, 96–111. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Helleringer, R.; Martucci, L.L.; Dallérac, G.; Cancela, J.-M.; Galante, M. Low Temperature Delays the Effects of Ischemia in Bergmann Glia and in Cerebellar Tissue Swelling. Biomedicines 2023, 11, 1363. https://doi.org/10.3390/biomedicines11051363
Li X, Helleringer R, Martucci LL, Dallérac G, Cancela J-M, Galante M. Low Temperature Delays the Effects of Ischemia in Bergmann Glia and in Cerebellar Tissue Swelling. Biomedicines. 2023; 11(5):1363. https://doi.org/10.3390/biomedicines11051363
Chicago/Turabian StyleLi, Xia, Romain Helleringer, Lora L. Martucci, Glenn Dallérac, José-Manuel Cancela, and Micaela Galante. 2023. "Low Temperature Delays the Effects of Ischemia in Bergmann Glia and in Cerebellar Tissue Swelling" Biomedicines 11, no. 5: 1363. https://doi.org/10.3390/biomedicines11051363