
Hemoglobin β Expression Is Associated with Poor Prognosis in Clear Cell Renal Cell Carcinoma
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Cell Lines
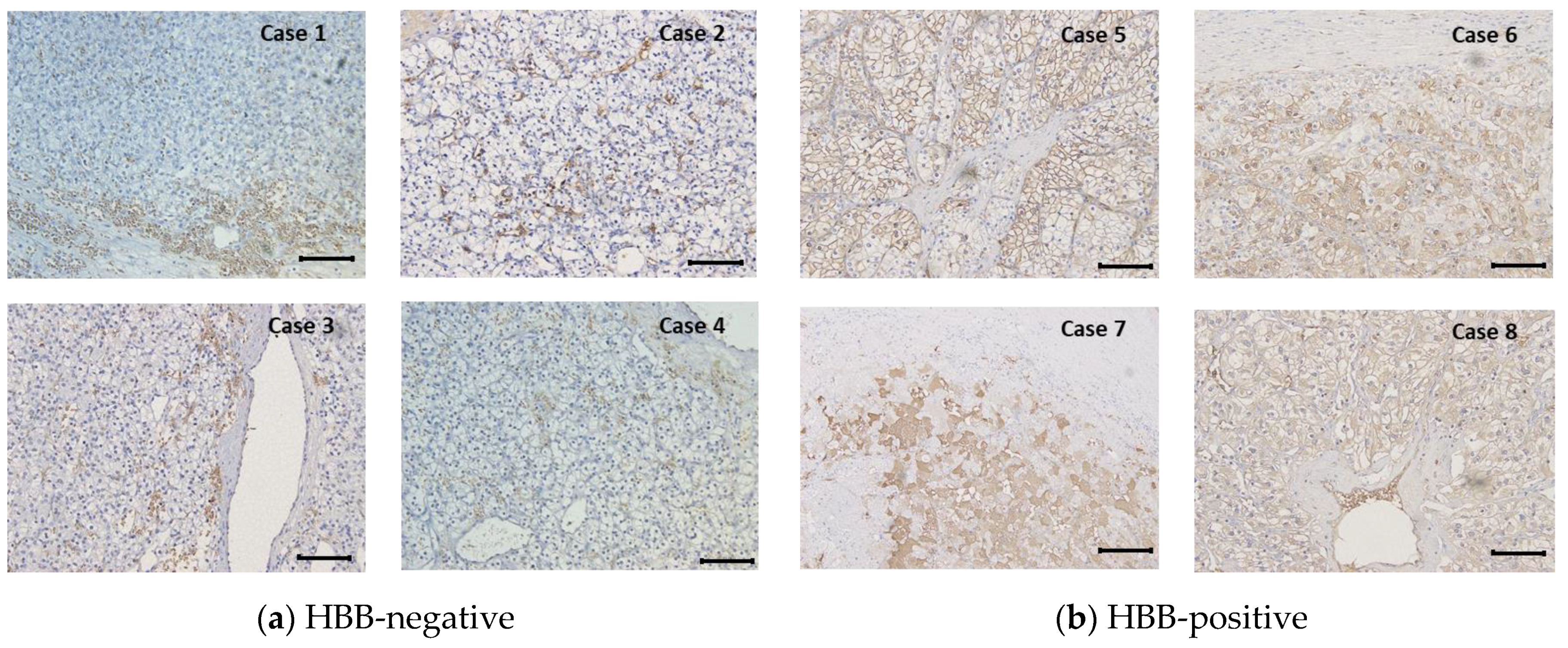
2.3. Immunohistochemical Observation
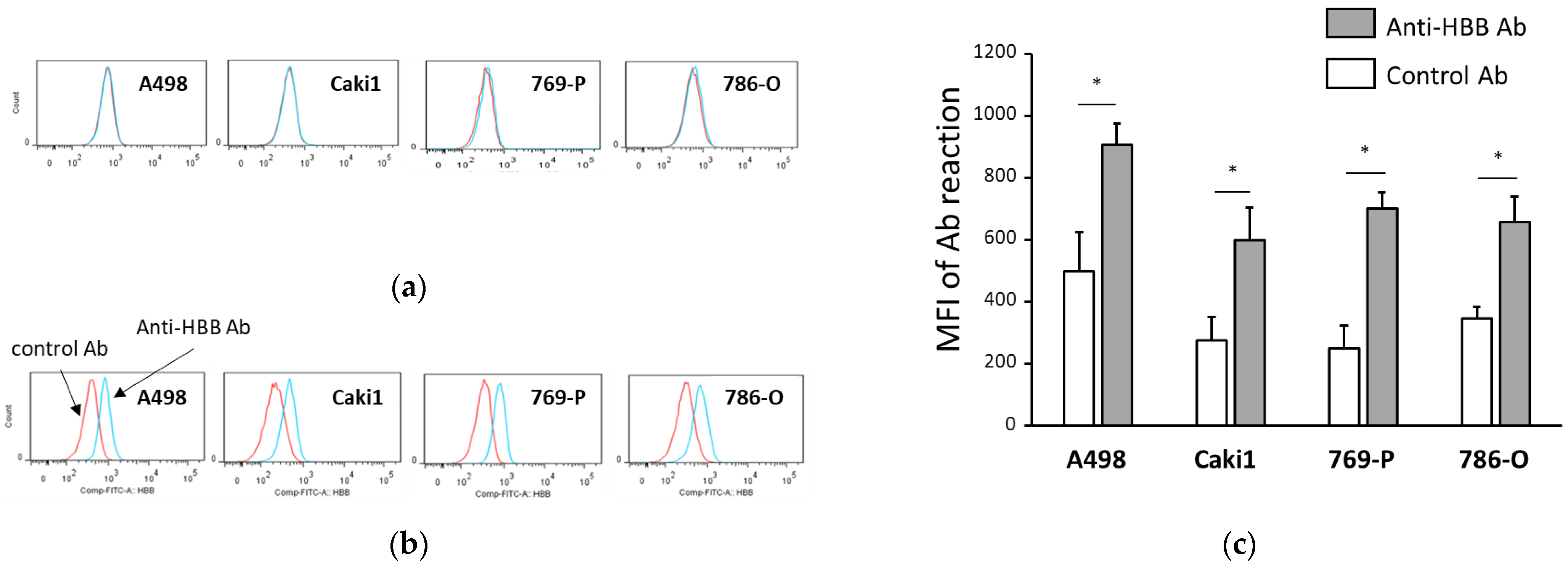
2.4. Measurement of HBB Expression Using Flow Cytometry
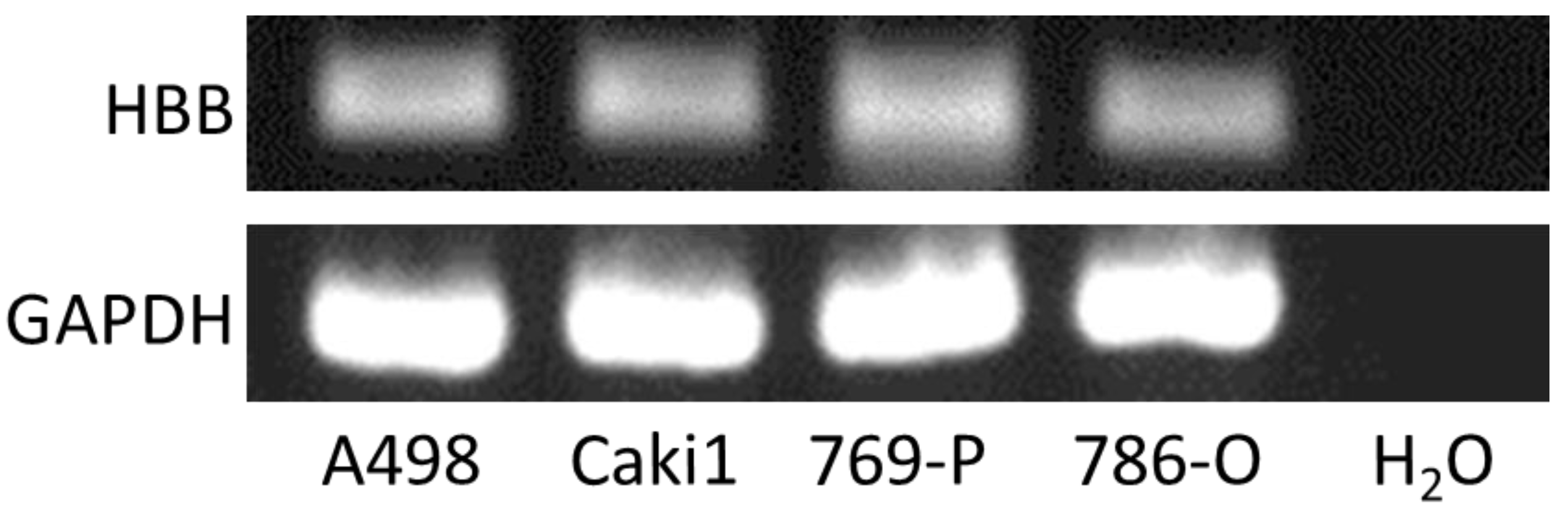
2.5. HBB mRNA Measurement by Quantitative Real-Time Reverse Transcript (RT) Polymerase Chain Reaction (PCR)
2.6. Suppression of HBB Expression by Short Interfering RNA (siRNA)
2.7. Cell Proliferation Assay
2.8. Cell Invasion Assay
2.9. ROS Measurement
2.10. Statistical Analysis
3. Results
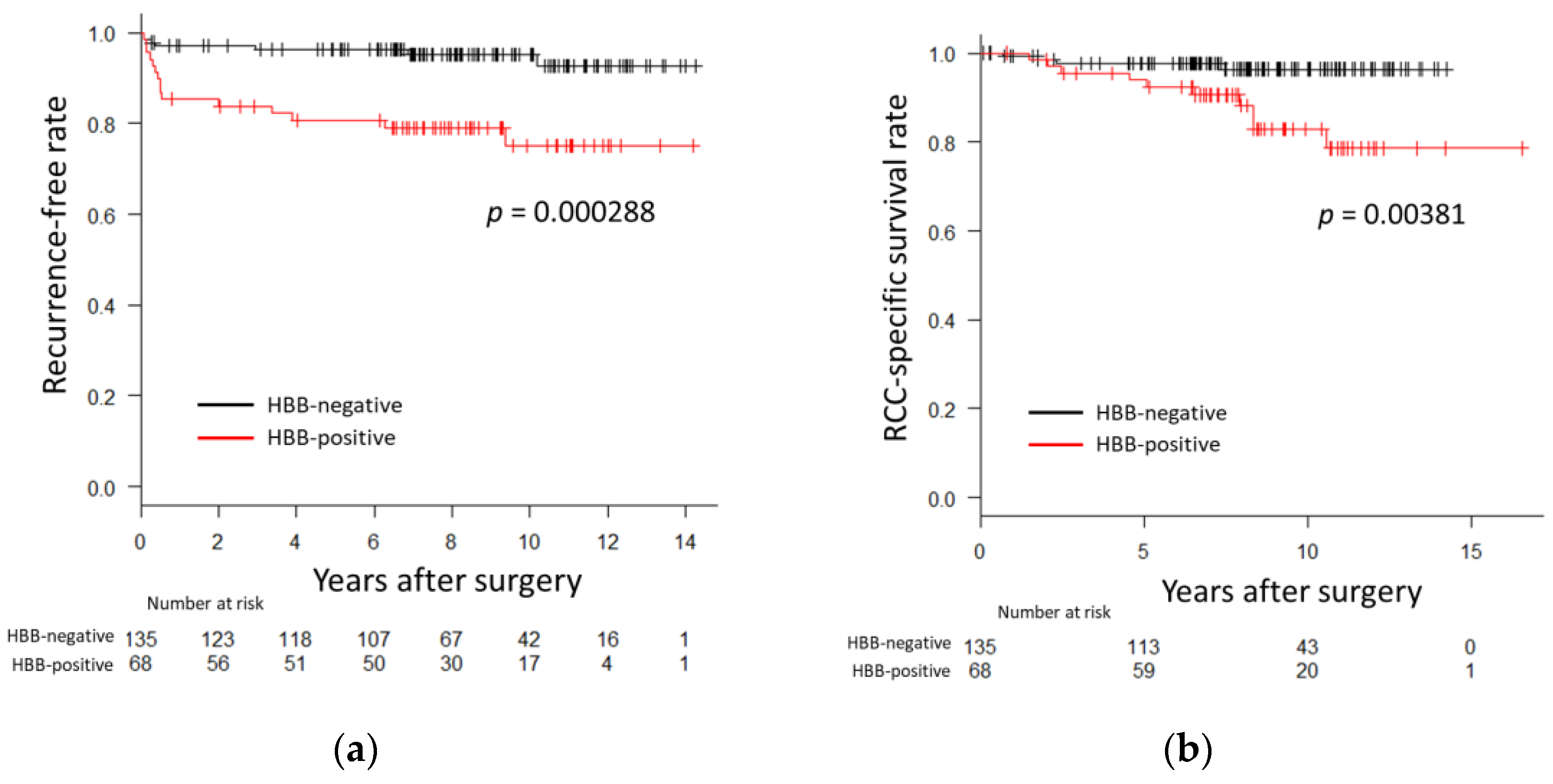
3.1. HBB Expression in ccRCC Is Associated with Prognosis
3.2. Several ccRCC Cell Lines Express HBB
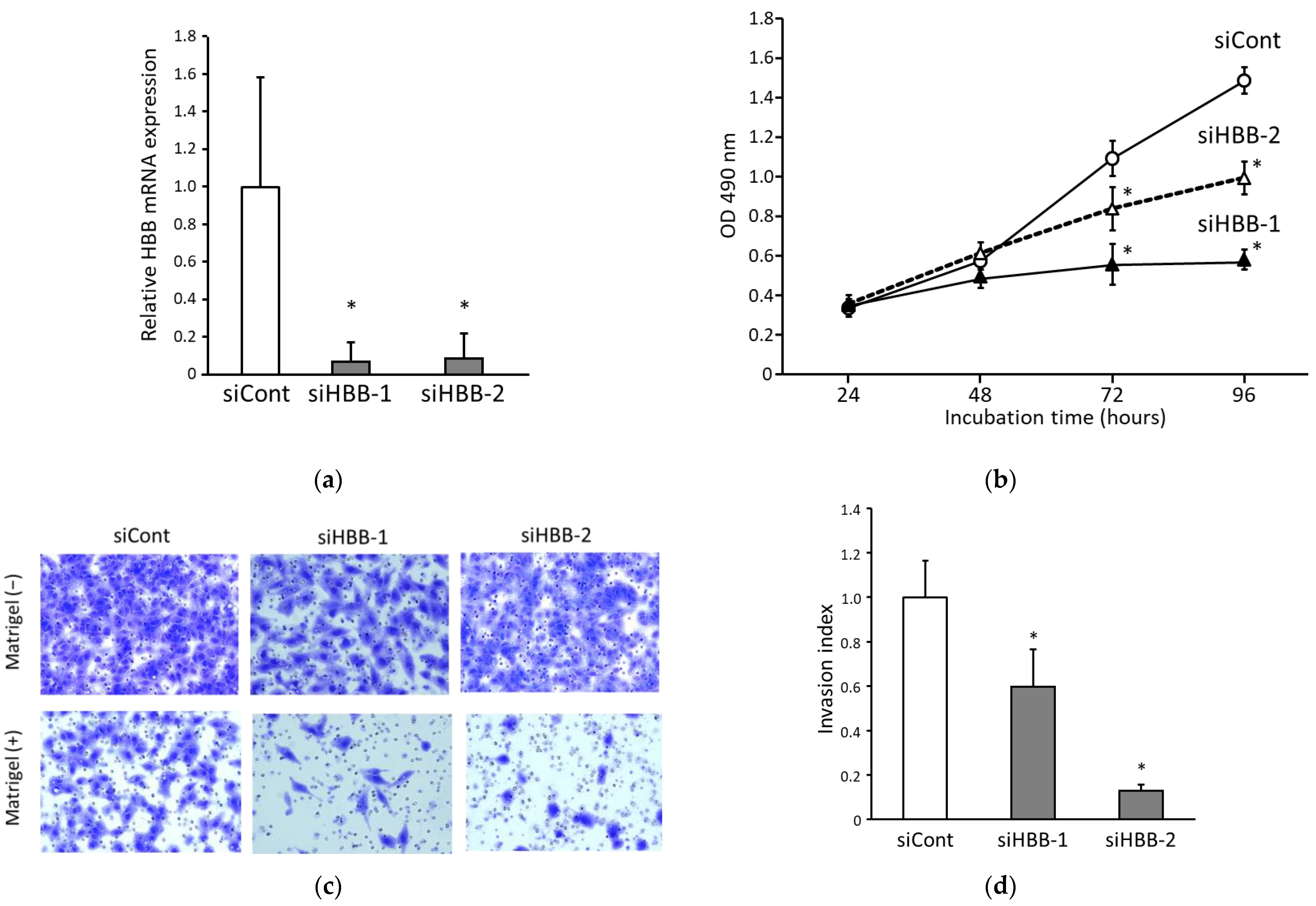
3.3. Suppression of HBB in ccRCC Cells Inhibits Both Cell Proliferation and Invasion
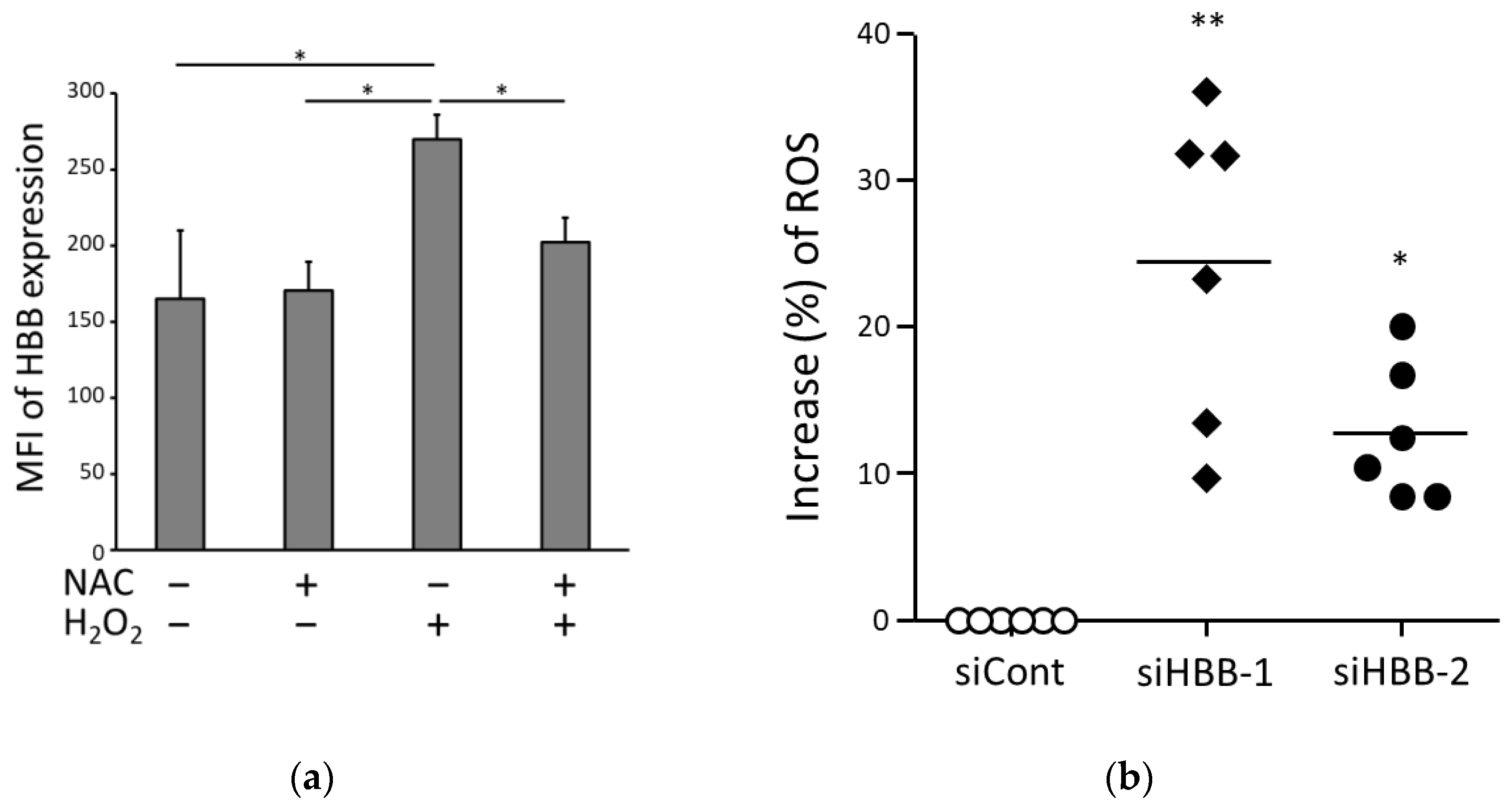
3.4. HBB Expression in ccRCC Cells Is Involved in Oxidative Adaptation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manuelli, V.; Pecorari, C.; Filomeni, G.; Zito, E. Regulation of Redox Signaling in HIF-1-Dependent Tumor Angiogenesis. FEBS J. 2022, 289, 5413–5425. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of Oxidative Stress as an Anticancer Strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Z.; Wang, Y.; Mei, Q.; Fu, X.; Han, W. Characterization of Adult α- and β-Globin Elevated by Hydrogen Peroxide in Cervical Cancer Cells that Play a Cytoprotective Role against Oxidative Insults. PLoS ONE 2013, 8, e54342. [Google Scholar] [CrossRef]
- Saha, D.; Patgaonkar, M.; Shroff, A.; Ayyar, K.; Bashir, T.; Reddy, K.V. Hemoglobin Expression in Nonerythroid Cells: Novel or Ubiquitous? Int. J. Inflam. 2014, 2014, 803237. [Google Scholar] [CrossRef]
- Bhaskaran, M.; Chen, H.; Chen, Z.; Liu, L. Hemoglobin is Expressed in Alveolar Epithelial Type II Cells. Biochem. Biophys. Res. Commun. 2005, 333, 1348–1352. [Google Scholar] [CrossRef]
- Grek, C.L.; Newton, D.A.; Spyropoulos, D.D.; Baatz, J.E. Hypoxia Up-Regulates Expression of Hemoglobin in Alveolar Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2011, 44, 439–447. [Google Scholar] [CrossRef]
- Newton, D.A.; Rao, K.M.; Dluhy, R.A.; Baatz, J.E. Hemoglobin is Expressed by Alveolar Epithelial Cells. J. Biol. Chem. 2006, 281, 5668–5676. [Google Scholar] [CrossRef]
- Tezel, T.H.; Geng, L.; Lato, E.B.; Schaal, S.; Liu, Y.; Dean, D.; Klein, J.B.; Kaplan, H.J. Synthesis and Secretion of Hemoglobin by Retinal Pigment Epithelium. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1911–1919. [Google Scholar] [CrossRef]
- Nishi, H.; Inagi, R.; Kato, H.; Tanemoto, M.; Kojima, I.; Son, D.; Fujita, T.; Nangaku, M. Hemoglobin is Expressed by Mesangial Cells and Reduces Oxidant Stress. J. Am. Soc. Nephrol. 2008, 19, 1500–1508. [Google Scholar] [CrossRef]
- Liu, W.; Baker, S.S.; Baker, R.D.; Nowak, N.J.; Zhu, L. Upregulation of Hemoglobin Expression by Oxidative Stress in Hepatocytes and Its Implication in Nonalcoholic Steatohepatitis. PLoS ONE 2011, 6, e24363. [Google Scholar] [CrossRef] [PubMed]
- Biagioli, M.; Pinto, M.; Cesselli, D.; Zaninello, M.; Lazarevic, D.; Roncaglia, P.; Simone, R.; Vlachouli, C.; Plessy, C.; Bertin, N.; et al. Unexpected Expression of Alpha- and Beta-Globin in Mesencephalic Dopaminergic Neurons and Glial Cells. Proc. Natl. Acad. Sci. USA 2009, 106, 15454–15459. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Zucchelli, S.; Codrich, M.; Marcuzzi, F.; Verde, C.; Gustincich, S. Hemoglobin is Present as a Canonical α2β2 Tetramer in Dopaminergic Neurons. Biochim. Biophys. Acta 2013, 1834, 1939–1943. [Google Scholar] [CrossRef] [PubMed]
- Schelshorn, D.W.; Schneider, A.; Kuschinsky, W.; Weber, D.; Kruger, C.; Dittgen, T.; Burgers, H.F.; Sabouri, F.; Gassler, N.; Bach, A.; et al. Expression of Hemoglobin in Rodent Neurons. J. Cereb. Blood Flow Metab. 2009, 29, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Ponzetti, M.; Capulli, M.; Angelucci, A.; Ventura, L.; Monache, S.D.; Mercurio, C.; Calgani, A.; Sanita, P.; Teti, A.; Rucci, N. Non-Conventional role of Haemoglobin Beta in Breast Malignancy. Br. J. Cancer 2017, 117, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Miyamoto, D.T.; Wittner, B.S.; Sullivan, J.P.; Aceto, N.; Jordan, N.V.; Yu, M.; Karabacak, N.M.; Comaills, V.; Morris, R.; et al. Expression of β-Globin by Cancer Cells Promotes Cell Survival During Blood-Borne Dissemination. Nat. Commun. 2017, 8, 14344. [Google Scholar] [CrossRef]
- Gupta, K.; Miller, J.D.; Li, J.Z.; Russell, M.W.; Charbonneau, C. Epidemiologic and Socioeconomic Burden of Metastatic Renal Cell Carcinoma (mRCC): A Literature Review. Cancer Treat. Rev. 2008, 34, 193–205. [Google Scholar] [CrossRef]
- Kanayama, H.O.; Fukumori, T.; Fujimoto, H.; Nakanishi, H.; Ohyama, C.; Suzuki, K.; Nishiyama, H.; Eto, M.; Miki, T.; Kamoi, K.; et al. Clinicopathological Characteristics and Oncological Outcomes in Patients with Renal Cell Carcinoma Registered in 2007: The First Large-Scale Multicenter Study from the Cancer Registration Committee of the Japanese Urological Association. Int. J. Urol. 2015, 22, S1–S7. [Google Scholar] [CrossRef]
- Stewart, S.B.; Thompson, R.H.; Psutka, S.P.; Cheville, J.C.; Lohse, C.M.; Boorjian, S.A.; Leibovich, B.C. Evaluation of the National Comprehensive Cancer Network and American Urological Association Renal Cell Carcinoma Surveillance Guidelines. J. Clin. Oncol. 2014, 32, 4059–4065. [Google Scholar] [CrossRef]
- Calvo, E.; Schmidinger, M.; Heng, D.Y.; Grunwald, V.; Escudier, B. Improvement in Survival End Points of Patients with Metastatic Renal Cell Carcinoma through Sequential Targeted Therapy. Cancer Treat. Rev. 2016, 50, 109–117. [Google Scholar] [CrossRef]
- Kanno, H.; Naito, S.; Obara, Y.; Ito, H.; Ichiyanagi, O.; Narisawa, T.; Kato, T.; Nagaoka, A.; Tsuchiya, N. Effect of Extracellular Signal-Regulated Protein Kinase 5 Inhibition in Clear Cell Renal Cell Carcinoma. Int. J. Mol. Sci. 2022, 23, 8448. [Google Scholar] [CrossRef] [PubMed]
- Moch, H.; Cubilla, A.L.; Humphrey, P.A.; Reuter, V.E.; Ulbright, T.M. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs-Part A: Renal, Penile, and Testicular Tumours. Eur. Urol. 2016, 70, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Ichiyanagi, O.; Ito, H.; Naito, S.; Kabasawa, T.; Kanno, H.; Narisawa, T.; Ushijima, M.; Kurota, Y.; Ozawa, M.; Sakurai, T.; et al. Impact of eIF4E Phosphorylation at Ser209 via MNK2a on Tumour Recurrence after Curative Surgery in Localized Clear Cell Renal Cell Carcinoma. Oncotarget 2019, 10, 4053–4068. [Google Scholar] [CrossRef] [PubMed]
- Kanda, Y. Investigation of the Freely Available Easy-To-Use Software ‘EZR’ for Medical Statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef]
- Dassen, H.; Kamps, R.; Punyadeera, C.; Dijcks, F.; de Goeij, A.; Ederveen, A.; Dunselman, G.; Groothuis, P. Haemoglobin Expression in Human Endometrium. Hum. Reprod. 2008, 23, 635–641. [Google Scholar] [CrossRef]
- Maman, S.; Sagi-Assif, O.; Yuan, W.; Ginat, R.; Meshel, T.; Zubrilov, I.; Keisari, Y.; Lu, W.; Witz, I.P. The Beta Subunit of Hemoglobin (HBB2/HBB) Suppresses Neuroblastoma Growth and Metastasis. Cancer Res. 2017, 77, 14–26. [Google Scholar] [CrossRef]
- Onda, M.; Akaishi, J.; Asaka, S.; Okamoto, J.; Miyamoto, S.; Mizutani, K.; Yoshida, A.; Ito, K.; Emi, M. Decreased Expression of Haemoglobin Beta (HBB) Gene in Anaplastic Thyroid Cancer and Recovery of Its Expression Inhibits Cell Growth. Br. J. Cancer 2005, 92, 2216–2224. [Google Scholar] [CrossRef]
- Masuoka, N.; Kodama, H.; Abe, T.; Wang, D.H.; Nakano, T. Characterization of Hydrogen Peroxide Removal Reaction by Hemoglobin in the Presence of Reduced Pyridine Nucleotides. Biochim. Biophys. Acta 2003, 1637, 46–54. [Google Scholar] [CrossRef]
- Finkel, T. Signal Transduction by Reactive Oxygen Species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting Cancer Cells by ROS-Mediated Mechanisms: A Radical Therapeutic Approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef]
- Schafer, Z.T.; Grassian, A.R.; Song, L.; Jiang, Z.; Gerhart-Hines, Z.; Irie, H.Y.; Gao, S.; Puigserver, P.; Brugge, J.S. Antioxidant and Oncogene Rescue of Metabolic Defects Caused by Loss of Matrix Attachment. Nature 2009, 461, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Harris, I.S.; Treloar, A.E.; Inoue, S.; Sasaki, M.; Gorrini, C.; Lee, K.C.; Yung, K.Y.; Brenner, D.; Knobbe-Thomsen, C.B.; Cox, M.A.; et al. Glutathione and Thioredoxin Antioxidant Pathways Synergize to Drive Cancer Initiation and Progression. Cancer Cell 2015, 27, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Piskounova, E.; Agathocleous, M.; Murphy, M.M.; Hu, Z.; Huddlestun, S.E.; Zhao, Z.; Leitch, A.M.; Johnson, T.M.; DeBerardinis, R.J.; Morrison, S.J. Oxidative Stress Inhibits Distant Metastasis by Human Melanoma Cells. Nature 2015, 527, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Chen, H.Y.; Harris, I.S.; Stover, D.G.; Selfors, L.M.; Bronson, R.T.; Deraedt, T.; Cichowski, K.; Welm, A.L.; Mori, Y.; et al. Cancer Cells Co-opt the Neuronal Redox-Sensing Channel TRPA1 to Promote Oxidative-Stress Tolerance. Cancer Cell 2018, 33, 985–1003.e7. [Google Scholar] [CrossRef]
- Lignitto, L.; LeBoeuf, S.E.; Homer, H.; Jiang, S.; Askenazi, M.; Karakousi, T.R.; Pass, H.I.; Bhutkar, A.J.; Tsirigos, A.; Ueberheide, B.; et al. Nrf2 Activation Promotes Lung Cancer Metastasis by Inhibiting the Degradation of Bach1. Cell 2019, 178, 316–329 e18. [Google Scholar] [CrossRef]
- Takemoto, K.; Kobatake, K.; Miura, K.; Fukushima, T.; Babasaki, T.; Miyamoto, S.; Sekino, Y.; Kitano, H.; Goto, K.; Ikeda, K.; et al. BACH1 promotes clear cell renal cell carcinoma progression by upregulating oxidative stress-related tumorigenicity. Cancer Sci. 2023, 114, 436–448. [Google Scholar] [CrossRef]
- Wiel, C.; Le Gal, K.; Ibrahim, M.X.; Jahangir, C.A.; Kashif, M.; Yao, H.; Ziegler, D.V.; Xu, X.; Ghosh, T.; Mondal, T.; et al. BACH1 Stabilization by Antioxidants Stimulates Lung Cancer Metastasis. Cell 2019, 178, 330–345.e22. [Google Scholar] [CrossRef]
- Gattoni, M.; Boffi, A.; Sarti, P.; Chiancone, E. Stability of the heme-globin linkage in alphabeta dimers and isolated chains of human hemoglobin. A study of the heme transfer reaction from the immobilized proteins to albumin. J. Biol. Chem. 1996, 271, 10130–10136. [Google Scholar] [CrossRef]
- Cheung, E.C.; Vousden, K.H. The role of ROS in tumour development and progression. Nat. Rev. Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef]
- Ding, Y.; Xiong, S.; Chen, X.; Pan, Q.; Fan, J.; Guo, J. HAPLN3 inhibits apoptosis and promotes EMT of clear cell renal cell carcinoma via ERK and Bcl-2 signal pathways. J. Cancer Res. Clin. Oncol. 2023, 149, 79–90. [Google Scholar] [CrossRef]
- Moi, P.; Chan, K.; Asunis, I.; Cao, A.; Kan, Y.W. Isolation of NF-E2-Related Factor 2 (Nrf2), a NF-E2-Like Basic Leucine Zipper Transcriptional Activator That Binds to the Tandem NF-E2/AP1 Repeat of the Beta-Globin Locus Control Region. Proc. Natl. Acad. Sci. USA 1994, 91, 9926–9930. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Yoshizato, T.; Shiraishi, Y.; Maekawa, S.; Okuno, Y.; Kamura, T.; Shimamura, T.; Sato-Otsubo, A.; Nagae, G.; Suzuki, H.; et al. Integrated Molecular Analysis of Clear-Cell Renal Cell Carcinoma. Nat. Genet. 2013, 45, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Kurota, Y.; Department of Urology, Yamagata University Faculty of Medicine, Yamagata, Japan; Takeda, Y.; Department of Immunology, Yamagata University Faculty of Medicine, Yamagata Japan. Personal communication, 2023.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | All Patients | HBB | |||
|---|---|---|---|---|---|
| Negative | Positive | p Value | |||
| Number of patients | 203 | 135 | 68 | ||
| Age at surgery | Mean ± SD | 63.51 ± 11.58 | 63.2 ± 11.7 | 64.1 ± 11.4 | 0.61 b |
| Range | 28–85 | 28–82 | 36–85 | ||
| Sex | Male | 125 | 86 (68.8) | 39 (31.2) | 0.468 a |
| Female | 78 | 49 (62.8) | 29 (37.2) | ||
| Tumor laterality | Left | 95 | 64 (67.4) | 31 (32.6) | 0.923 a |
| Right | 108 | 71 (65.7) | 37 (34.3) | ||
| Pathological T stage | pT1a | 124 | 100 (74.1) | 24 (35.3) | <0.001 a |
| pT1b | 40 | 19 (14.1) | 21 (30.9) | ||
| pT2a | 8 | 2 (1.5) | 6 (8.8) | ||
| pT2b | 6 | 4 (3.0) | 2 (2.9) | ||
| pT3a | 15 | 7 (5.2) | 8 (11.8) | ||
| pT3b | 7 | 2 (1.5) | 5 (7.4) | ||
| pT3c | 1 | 0 (0.0) | 1 (1.5) | ||
| pT4 | 2 | 1 (0.7) | 1 (1.5) | ||
| Pathological N stage | pN0/pNX | 199 (98.0) | 134 (99.3) | 65 (86.8) | 0.0021 a |
| pN1 | 4 (2.0) | 1 (0.7) | 3 (4.4) | ||
| Grade | G1 | 96 | 80 (59.3) | 16 (23.5) | <0.001 a |
| G2 | 82 | 48 (35.6) | 34 (50.0) | ||
| G3 | 20 | 4 (3.0) | 16 (23.5) | ||
| G4 | 5 | 3 (2.2) | 2 (2.9) | ||
| Microvascular invasion | Negative | 180 (88.6) | 127 (70.6) | 53 (29.4) | 0.00143 a |
| Positive | 23 (11.4) | 8 (34.8) | 15 (65.2) | ||
| INF | INFa | 171 (84.2) | 121 (89.6) | 50 (73.5) | 0.00566 a |
| INFb | 32 (15.8) | 14 (10.4) | 18 (26.5) | ||
| Event | |||||
| Recurrence | No | 181 | 128 (70.7) | 53 (29.3) | <0.001 a |
| Yes | 22 | 7 (31.8) | 15 (68.2) | ||
| Cancer specific survival | Alive | 189 | 131 (69.3) | 58 (30.7) | 0.00476 a |
| Deceased | 14 | 4 (28.6) | 10 (71.4) | ||
| Overall survival | Alive | 169 | 119 (70.4) | 50 (29.6) | 0.015 a |
| Deceased | 34 | 16 (47.1) | 18 (52.9) | ||
| Follow-up Years | Median | 8.12 | 8.06 | 8.24 | 0.797 c |
| (IQR) | (6.56–10.72) | (6.52–10.79) | (6.69–10.69) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurota, Y.; Takeda, Y.; Ichiyanagi, O.; Saitoh, S.; Ito, H.; Naito, S.; Asao, H.; Tsuchiya, N. Hemoglobin β Expression Is Associated with Poor Prognosis in Clear Cell Renal Cell Carcinoma. Biomedicines 2023, 11, 1330. https://doi.org/10.3390/biomedicines11051330
Kurota Y, Takeda Y, Ichiyanagi O, Saitoh S, Ito H, Naito S, Asao H, Tsuchiya N. Hemoglobin β Expression Is Associated with Poor Prognosis in Clear Cell Renal Cell Carcinoma. Biomedicines. 2023; 11(5):1330. https://doi.org/10.3390/biomedicines11051330
Chicago/Turabian StyleKurota, Yuta, Yuji Takeda, Osamu Ichiyanagi, Shinichi Saitoh, Hiromi Ito, Sei Naito, Hironobu Asao, and Norihiko Tsuchiya. 2023. "Hemoglobin β Expression Is Associated with Poor Prognosis in Clear Cell Renal Cell Carcinoma" Biomedicines 11, no. 5: 1330. https://doi.org/10.3390/biomedicines11051330