
Age-Dependent Changes in Calcium Regulation after Myocardial Ischemia–Reperfusion Injury
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Langendorff Model of Ischemia–Reperfusion
2.3. Preparation of Tissue Homogenates and SR Vesicles
2.4. Assay of Ca2+-ATPase Activity
2.5. Dot Blot
2.6. Western Blot and Immunodetection
2.7. Data Analysis
3. Results
3.1. Effect of IR on Contractile Function Parameters during Aging
3.2. Ca2+-ATPase Activity in Aging Hearts after IR
3.3. Impact of Myocardial IR on SERCA2a and PLN Contents during Aging
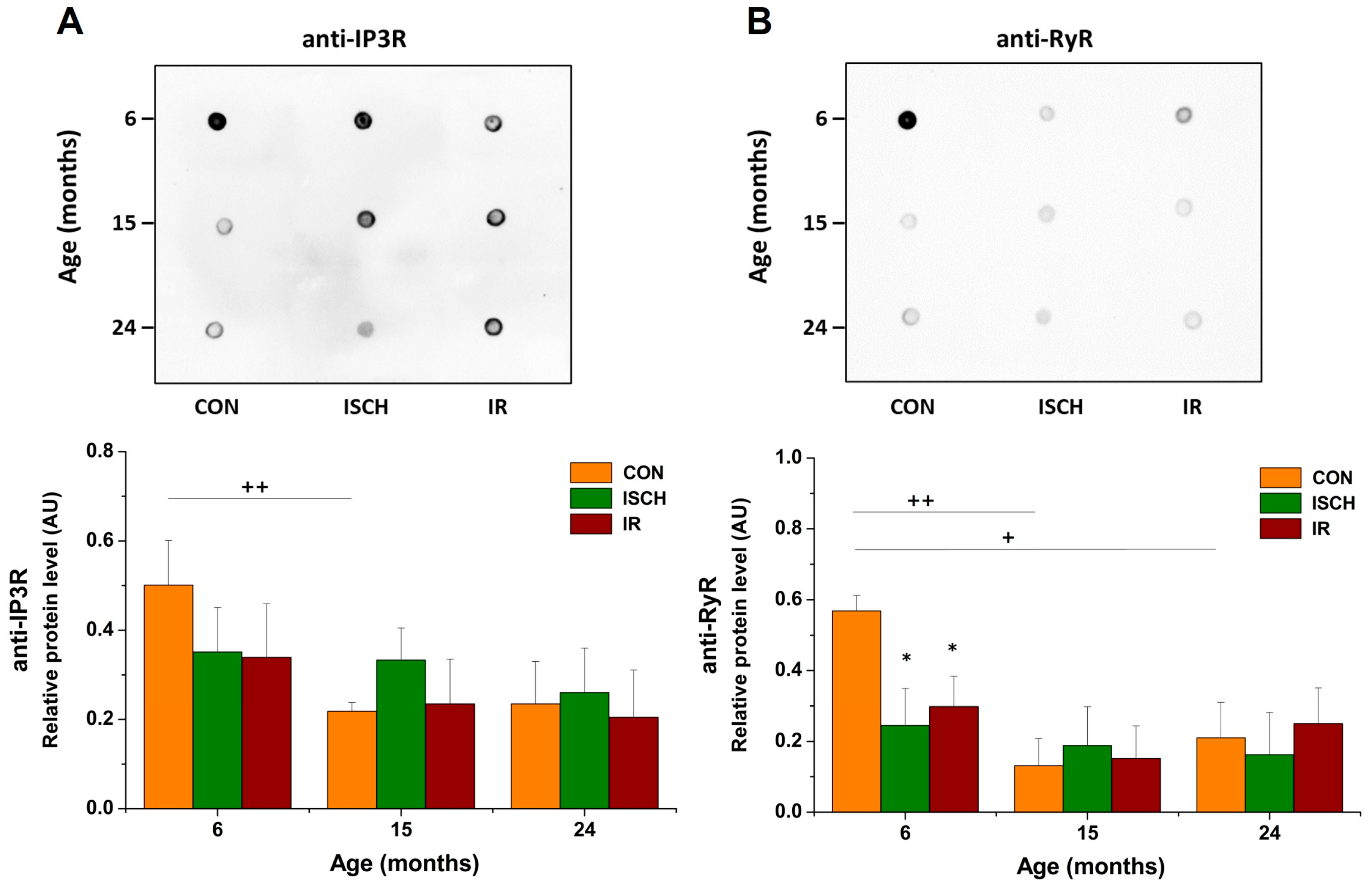
3.4. Impact of Myocardial IR on the Level of Other Ca2+-Regulating Proteins during Aging
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cardiovascular Diseases (CVDs). 2021. Available online: www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 11 June 2021).
- Díaz-Vesga, M.C.; Zúñiga-Cuevas, Ú.; Ramírez-Reyes, A.; Herrera-Zelada, N.; Palomo, I.; Bravo-Sagua, R.; Riquelme, J.A. Potential therapies to protect the aging heart against ischemia/reperfusion injury. Front. Cardiovasc. Med. 2021, 8, 770421. [Google Scholar] [CrossRef] [PubMed]
- Loubani, M.; Ghosh, S.; Galiñanes, M. The aging human myocardium: Tolerance to ischemia and responsiveness to ischemic preconditioning. J. Thorac. Cardiovasc. Surg. 2003, 126, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Boucher, F.; Tanguy, S.; Besse, S.; Tresallet, N.; Favier, A.; de Leiris, J. Age-dependent changes in myocardial susceptibility to zero flow ischemia and reperfusion in isolated perfused rat hearts: Relation to antioxidant status. Mech. Ageing Dev. 1998, 103, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Sniecinski, R.; Liu, H. Reduced efficacy of volatile anesthetic preconditioning with advanced age in isolated rat myocardium. Anesthesiology 2004, 100, 589–597. [Google Scholar] [CrossRef]
- Riess, M.L.; Camara, A.K.S.; Rhodes, S.S.; McCormick, J.; Jiang, M.T.; Stowe, D.F. Increasing heart size and age attenuate anesthetic preconditioning in guinea pig isolated hearts. Anesth. Analg. 2005, 101, 1572–1576. [Google Scholar] [CrossRef]
- Headrick, J.P. Aging impairs functional, metabolic and ionic recovery from ischemia-reperfusion and hypoxia-reoxygenation. J. Mol. Cell. Cardiol. 1998, 30, 1415–1430. [Google Scholar] [CrossRef]
- Tani, M.; Suganuma, Y.; Hasegawa, H.; Shinmura, K.; Ebihara, Y.; Hayashi, Y.; Guo, X.; Takayama, M. Decrease in ischemic tolerance with aging in isolated perfused Fischer 344 rat hearts: Relation to increases in intracellular Na+ after ischemia. J. Mol. Cell. Cardiol. 1997, 29, 3081–3089. [Google Scholar] [CrossRef]
- Quan, N.; Sun, W.; Wang, L.; Chen, X.; Bogan, J.S.; Zhou, X.; Li, J. Sestrin2 prevents age-related intolerance to ischemia and reperfusion injury by modulating substrate metabolism. FASEB J. 2017, 31, 4153–4167. [Google Scholar] [CrossRef]
- Noble, M.I. Whatever happened to measuring ventricular contractility in heart failure? Card. Fail. Rev. 2017, 3, 79–82. [Google Scholar] [CrossRef]
- Valera-Alberni, M.; Canto, C. Mitochondrial stress management: A dynamic journey. Cell Stress 2018, 2, 253–274. [Google Scholar] [CrossRef]
- De Lucia, C.; Eguchi, A.; Koch, W.J. New insights in cardiac β-adrenergic signaling during heart failure and aging. Front. Pharmacol. 2018, 9, 904. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.B.; Kalkhoran, S.B.; Hernández-Reséndiz, S.; Samangouei, P.; Ong, S.G.; Hausenloy, D.J. Mitochondrial-shaping proteins in cardiac health and disease–the long and the short of it! Cardiovasc. Drugs Ther. 2017, 31, 87–107. [Google Scholar] [CrossRef]
- Hoppel, C.L.; Tandler, B.; Fujioka, H.; Riva, A. Dynamic organization of mitochondria in human heart and in myocardial disease. Int. J. Biochem. Cell Biol. 2009, 41, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Nunnari, J.; Suomalainen, A. Mitochondria: In sickness and in health. Cell 2012, 148, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Meana, M.; Bou-Teen, D.; Ferdinandy, P.; Gyongyosi, M.; Pesce, M.; Perrino, C.; Schulz, R.; Sluijter, J.P.G.; Tocchetti, C.G.; Thum, T.; et al. Cardiomyocyte ageing and cardioprotection: Consensus document from the ESC working groups cell biology of the heart and myocardial function. Cardiovasc. Res. 2020, 116, 1835–1849. [Google Scholar] [CrossRef] [PubMed]
- Boengler, K.; Kosiol, M.; Mayr, M.; Schulz, R.; Rohrbach, S. Mitochondria and ageing: Role in heart, skeletal muscle and adipose tissue. J. Cachexia Sarcopenia Muscle 2017, 8, 349–369. [Google Scholar] [CrossRef]
- Cooper, L.L.; Li, W.; Lu, Y.; Centracchio, J.; Terentyeva, R.; Koren, G.; Terentyev, D. Redox modification of ryanodine receptors by mitochondria-derived reactive oxygen species contributes to aberrant Ca2+ handling in ageing rabbit hearts. J. Physiol. 2013, 591, 5895–5911. [Google Scholar] [CrossRef]
- Janczewski, A.M.; Lakatta, E.G. Modulation of sarcoplasmic reticulum Ca2+ cycling in systolic and diastolic heart failure associated with aging. Heart Fail. Rev. 2010, 15, 431–445. [Google Scholar] [CrossRef]
- Feridooni, H.A.; Dibb, K.M.; Howlett, S.E. How cardiomyocyte excitation, calcium release and contraction become altered with age. J. Mol. Cell. Cardiol. 2015, 83, 62–72. [Google Scholar] [CrossRef]
- Fernandez-Sanz, C.; Ruiz-Meana, M.; Castellano, J.; Miro-Casas, E.; Nuñez, E.; Inserte, J.; Garcia-Dorado, D. Altered FoF1 ATP synthase and susceptibility to mitochondrial permeability transition pore during ischaemia and reperfusion in aging cardiomyocytes. Thromb. Haemost. 2015, 113, 441–451. [Google Scholar] [CrossRef]
- Piquereau, J.; Caffin, F.; Novotova, M.; Lemaire, C.; Veksler, V.; Garnier, A.; Ventura-Clapier, R.; Joubert, F. Mitochondrial dynamics in the adult cardiomyocytes: Which roles for a highly specialized cell? Front. Physiol. 2013, 4, 102. [Google Scholar] [CrossRef]
- Verdejo, H.E.; Del Campo, A.; Troncoso, R.; Gutierrez, T.; Toro, B.; Quiroga, C.; Pedrozo, Z.; Munoz, J.P.; Garcia, L.; Castro, P.F.; et al. Mitochondria, myocardial remodeling, and cardiovascular disease. Curr. Hypertens. Rep. 2012, 14, 532–539. [Google Scholar] [CrossRef]
- Henning, R.H.; Brundel, B.J. Proteostasis in cardiac health and disease. Nat. Rev. Cardiol. 2017, 14, 637–653. [Google Scholar] [CrossRef]
- Chung, J.; Tilūnaitė, A.; Ladd, D.; Hunt, H.; Soeller, C.; Crampin, E.J.; Johnston, S.T.; Roderick, H.L.; Rajagopal, V. IP3R activity increases propensity of RyR-mediated sparks by elevating dyadic [Ca2+]. Math. Biosci. 2023, 355, 108923. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Kim, K.; Kim, J.H.; Park, Y. The role of endoplasmic reticulum stress in cardiovascular disease and exercise. Int. J. Vasc. Med. 2017, 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gao, H.; Zhou, H.; Liu, Q.; Qi, Z.; Zhang, Y.; Zhang, J. The role of mitochondria-derived peptides in cardiovascular disease: Recent updates. Biomed. Pharmacother. 2019, 117, 109075. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.D.; Enomoto, M.; Ishiyama, N.; Stathopulos, P.B.; Ikura, M. Structural insights into endoplasmic reticulum stored calcium regulation by inositol 1,4,5-trisphosphate and ryanodine receptors. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 1980–1991. [Google Scholar] [CrossRef] [PubMed]
- Fares, E.; Howlett, S.E. Effect of age on cardiac excitation–contraction coupling. Clin. Exp. Pharmacol. Physiol. 2010, 37, 1–7. [Google Scholar] [CrossRef]
- Roderick, H.L.; Bootman, M.D. Calcium influx: Is Homer the missing link? Curr. Biol. 2003, 13, R976–R978. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.; Rhee, J.; Chaudhari, V.; Rosenzweig, A. The role of exercise in cardiac aging: From physiology to molecular mechanisms. Circ. Res. 2016, 118, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Prather, E.R.; Garrison, D.E.; Zuo, L. Interplay between ROS and antioxidants during ischemia-reperfusion injuries in cardiac and skeletal muscle. Int. J. Mol. Sci. 2018, 19, 417. [Google Scholar] [CrossRef]
- Taffet, G.E.; Tate, C.A. CaATPase content is lower in cardiac sarcoplasmic reticulum isolated from old rats. Am. J. Physiol. Heart Circ. 1993, 264, H1609–H1614. [Google Scholar] [CrossRef]
- Schmidt, U.; Del Monte, F.; Miyamoto, M.I.; Matsui, T.; Gwathmey, J.K.; Rosenzweig, A.; Hajjar, R.J. Restoration of diastolic function in senescent rat hearts through adenoviral gene transfer of sarcoplasmic reticulum Ca2+-ATPase. Circulation 2000, 101, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Saeed, Y.; Temple, I.P.; Borbas, Z.; Atkinson, A.; Yanni, J.; Maczewski, M.; Mackiewicz, U.; Aly, M.; Logantha, S.J.R.J.; Garratt, C.J.; et al. Structural and functional remodeling of the atrioventricular node with aging in rats: The role of hyperpolarization-activated cyclic nucleotide–gated and ryanodine 2 channels. Heart Rhythm 2018, 15, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yan, F.; Li, L.; Fu, H.; Song, D.; Wu, D.; Wang, X. New focuses on roles of communications between endoplasmic reticulum and mitochondria in identification of biomarkers and targets. Clin. Transl. Med. 2021, 11, e626. [Google Scholar] [CrossRef] [PubMed]
- Giamogante, F.; Barazzuol, L.; Brini, M.; Calì, T. Er–mitochondria contact sites reporters: Strengths and weaknesses of the available approaches. Int. J. Mol. Sci. 2020, 21, 8157. [Google Scholar] [CrossRef] [PubMed]
- Mughal, W.; Martens, M.; Field, J.; Chapman, D.; Huang, J.; Rattan, S.; Hai, Y.; Cheung, K.G.; Kereliuk, S.; West, A.R.; et al. Myocardin regulates mitochondrial calcium homeostasis and prevents permeability transition. Cell Death Differ. 2018, 25, 1732–1748. [Google Scholar] [CrossRef]
- Hamilton, S.; Terentyev, D. Altered intracellular calcium homeostasis and arrhythmogenesis in the aged heart. Int. J. Mol. Sci. 2019, 20, 2386. [Google Scholar] [CrossRef]
- Bolli, R.; Marbán, E. Molecular and cellular mechanisms of myocardial stunning. Physiol. Rev. 1999, 79, 609–634. [Google Scholar] [CrossRef] [PubMed]
- Kloner, R.A. Stunned and Hibernating Myocardium: Where Are We Nearly 4 Decades Later? J. Am. Heart Assoc. 2020, 9, e015502. [Google Scholar] [CrossRef]
- Tatarkova, Z.; Kovalska, M.; Sivonova, M.K.; Racay, P.; Lehotsky, J.; Kaplan, P. Tyrosine nitration of mitochondrial proteins during myocardial ischemia and reperfusion. J. Physiol. Biochem. 2019, 75, 217–227. [Google Scholar] [CrossRef]
- Palmer, B.S.; Hadziahmetovic, M.; Veci, T.; Angelos, M.G. Global ischemic duration and reperfusion function in the isolated perfused rat heart. Resuscitation 2004, 62, 97–106. [Google Scholar] [CrossRef]
- Kaplan, P.; Babusikova, E.; Lehotsky, J.; Dobrota, D. Free radical-induced protein modification and inhibition of Ca2+-ATPase of cardiac sarcoplasmic reticulum. Mol. Cell. Biochem. 2003, 248, 41–47. [Google Scholar] [CrossRef]
- Partridge, L.; Deelen, J.; Slagboom, P.E. Facing up to the global challenges of ageing. Nature 2018, 561, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hastings, M.H.; Rhee, J.; Trager, L.E.; Roh, J.D.; Rosenzweig, A. Targeting age-related pathways in heart failure. Circ. Res. 2020, 126, 533–551. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lee, C.K.; Kang, S.; Park, I.; Kim, Y.H.; Kim, S.K.; Hong, S.P.; Bae, H.; He, Y.; Kubota, Y.; et al. Angiopoietin-2 exacerbates cardiac hypoxia and inflammation after myocardial infarction. J. Clin. Investig. 2018, 128, 5018–5033. [Google Scholar] [CrossRef]
- Mullany, L.K.; Rohira, A.D.; Leach, J.P.; Kim, J.H.; Monroe, T.O.; Ortiz, A.R.; Stork, B.; Gaber, M.W.; Sarkar, P.; Sikora, A.G.; et al. A steroid receptor coactivator stimulator (MCB-613) attenuates adverse remodeling after myocardial infarction. Proc. Natl. Acad. Sci. USA 2020, 117, 31353–31364. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Sreejayan, N.; Ren, J. Views from within and beyond: Narratives of cardiac contractile dysfunction under senescence. Endocrine 2005, 26, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Willems, L.; Zatta, A.; Holmgren, K.; Ashton, K.J.; Headrick, J.P. Age-related changes in ischemic tolerance in male and female mouse hearts. J. Mol. Cell. Cardiol. 2005, 38, 245–256. [Google Scholar] [CrossRef]
- Headrick, J.P.; Willems, L.; Ashton, K.J.; Holmgren, K.; Peart, J.; Matherne, G.P. Ischaemic tolerance in aged mouse myocardium: The role of adenosine and effects of A1 adenosine receptor overexpression. J. Physiol. 2003, 549, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Fert-Bober, J.; Basran, R.S.; Sawicka, J.; Sawicki, G. Effect of duration of ischemia on myocardial proteome in ischemia/reperfusion injury. Proteomics 2008, 8, 2543–2555. [Google Scholar] [CrossRef] [PubMed]
- Tourki, B.; Matéo, P.; Morand, J.; Elayeb, M.; Godin-Ribuot, D.; Marrakchi, N.; Belaidi, E.; Messadi, E. Lebetin 2, a snake venom-derived natriuretic peptide, attenuates acute myocardial ischemic injury through the modulation of mitochondrial permeability transition pore at the time of reperfusion. PLoS ONE 2016, 11, e0162632. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, S.S.; Camara, A.K.; Heisner, J.S.; Riess, M.L.; Aldakkak, M.; Stowe, D.F. Reduced mitochondrial Ca2+ loading and improved functional recovery after ischemia-reperfusion injury in old vs. young guinea pig hearts. Am. J. Physiol. Heart Circ. 2012, 302, H855–H863. [Google Scholar] [CrossRef] [PubMed]
- Hacker, T.A.; McKiernan, S.H.; Douglas, P.S.; Wanagat, J.; Aiken, J.M. Age-related changes in cardiac structure and function in Fischer 344× Brown Norway hybrid rats. Am. J. Physiol. Heart Circ. 2006, 290, H304–H311. [Google Scholar] [CrossRef] [PubMed]
- Preston, C.C.; Oberlin, A.S.; Holmuhamedov, E.L.; Gupta, A.; Sagar, S.; Syed, R.H.K.; Siddiqui, S.A.; Raghavakaimal, S.; Terzic, A.; Jahangir, A. Aging-induced alterations in gene transcripts and functional activity of mitochondrial oxidative phosphorylation complexes in the heart. Mech. Ageing Dev. 2008, 129, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Xu, T.; Luo, Y.; Zhang, Y.; Xuan, H.; Ma, Y.; Pan, D.; Zhu, H.; Li, D. Luteolin enhances sarcoplasmic reticulum Ca2+-ATPase activity through p38 MAPK signaling thus improving rat cardiac function after ischemia/reperfusion. Cell. Physiol. Biochem. 2017, 41, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Giladi, M.; Tal, I.; Khananshvili, D. Structural features of ion transport and allosteric regulation in sodium-calcium exchanger (NCX) proteins. Front. Physiol. 2016, 7, 30. [Google Scholar] [CrossRef]
- Chu, L.; Greenstein, J.L.; Winslow, R.L. Modeling Na+-Ca2+ exchange in the heart: Allosteric activation, spatial localization, sparks and excitation-contraction coupling. J. Mol. Cell. Cardiol. 2016, 99, 174–187. [Google Scholar] [CrossRef]
- Khananshvili, D. The SLC8 gene family of sodium–calcium exchangers (NCX)–Structure, function, and regulation in health and disease. Mol. Asp. Med. 2013, 34, 220–235. [Google Scholar] [CrossRef]
- Periasamy, M.; Bhupathy, P.; Babu, G.J. Regulation of sarcoplasmic reticulum Ca2+ ATPase pump expression and its relevance to cardiac muscle physiology and pathology. Cardiovasc. Res. 2008, 77, 265–273. [Google Scholar] [CrossRef]
- Liu, A.Y.; Aguayo-Ortiz, R.; Guerrero-Serna, G.; Wang, N.; Blin, M.G.; Goldstein, D.R.; Espinoza-Fonseca, L.M. Homologous cardiac calcium pump regulators phospholamban and sarcolipin adopt distinct oligomeric states in the membrane. Comput. Struct. Biotechnol. J. 2022, 20, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.M.; Hou, Z.; Bossuyt, J.; Bers, D.M.; Robia, S.L. Phospholamban oligomerization, quaternary structure, and sarco (endo) plasmic reticulum calcium ATPase binding measured by fluorescence resonance energy transfer in living cells. J. Biol. Chem. 2008, 283, 12202–12211. [Google Scholar] [CrossRef]
- Reddy, L.G.; Jones, L.R.; Thomas, D.D. Depolymerization of phospholamban in the presence of calcium pump: A fluorescence energy transfer study. Biochemistry 1999, 38, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, D.H.; Kranias, E.G. Phospholamban: A crucial regulator of cardiac contractility. Nat. Rev. Mol. Cell Biol. 2003, 4, 566–577. [Google Scholar] [CrossRef]
- Hamstra, S.I.; Whitley, K.C.; Baranowski, R.W.; Kurgan, N.; Braun, J.L.; Messner, H.N.; Fajardo, V.A. The role of phospholamban and GSK3 in regulating rodent cardiac SERCA function. Am. J. Physiol. Cell Physiol. 2020, 319, C694–C699. [Google Scholar] [CrossRef]
- Tian, R.; Halow, J.M.; Meyer, M.; Dillmann, W.H.; Figueredo, V.M.; Ingwall, J.S.; Camacho, S.A. Thermodynamic limitation for Ca2+ handling contributes to decreased contractile reserve in rat hearts. Am. J. Physiol. 1998, 275, H2064–H2071. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lu, X.; Rong, X.; Jiang, W.; Lai, D.; Ma, Y.; Zhou, K.; Fe, G.; Xu, S. Inhibition of the mevalonate pathway ameliorates anoxia-induced down-regulation of FKBP12. 6 and intracellular calcium handling dysfunction in H9c2 cells. J. Mol. Cell. Cardiol. 2015, 80, 166–174. [Google Scholar] [CrossRef]
- Zucchi, R.; Ronca-Testoni, S.; Yu, G.; Galbani, P.; Ronca, G.; Mariani, M. Effect of ischemia and reperfusion on cardiac ryanodine receptors--sarcoplasmic reticulum Ca2+ channels. Circ. Res. 1994, 74, 271–280. [Google Scholar] [CrossRef]
- Li, W.; Liu, B.; Wang, L.; Liu, J.; Yang, X.; Zheng, J. Melatonin attenuates cardiac ischemia-reperfusion injury through modulation of IP3R-mediated mitochondria-ER contact. Oxid. Med. Cell. Longev. 2021, 2021, 12. [Google Scholar] [CrossRef]
- Patergnani, S.; Suski, J.M.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Giorgi, C.; Marchi, S.; Missiroli, S.; Poletti, F.; et al. Calcium signaling around mitochondria associated membranes (MAMs). Cell Commun. Signal. 2011, 9, 19. [Google Scholar] [CrossRef]
- Hodge, T.; Colombini, M. Regulation of metabolite flux through voltage-gating of VDAC channels. J. Membr. Biol. 1997, 157, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Klapper-Goldstein, H.; Verma, A.; Elyagon, S.; Gillis, R.; Murninkas, M.; Pittala, S.; Paul, A.; Shoshan-Barmatz, V.; Etzion, Y. VDAC1 in the diseased myocardium and the effect of VDAC1-interacting compound on atrial fibrosis induced by hyperaldosteronism. Sci. Rep. 2020, 10, 22101. [Google Scholar] [CrossRef] [PubMed]
- Branco, A.F.; Pereira, S.L.; Moreira, A.C.; Holy, J.; Sardão, V.A.; Oliveira, P.J. Isoproterenol cytotoxicity is dependent on the differentiation state of the cardiomyoblast H9c2 cell line. Cardiovasc. Toxicol. 2011, 11, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Liu, D.; Tang, L.; Yin, D.; Yin, S.; Lai, S.; Yao, J.; He, M. Long-term oral resveratrol intake provides nutritional preconditioning against myocardial ischemia/reperfusion injury: Involvement of VDAC1 downregulation. Mol. Nutr. Food Res. 2015, 59, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, X.; Fujioka, H.; Liu, J.; Chen, S.; Zhu, X. DJ-1 regulates the integrity and function of ER-mitochondria association through interaction with IP3R3-Grp75-VDAC1. Proc. Natl. Acad. Sci. USA 2019, 116, 25322–25328. [Google Scholar] [CrossRef] [PubMed]
- Honrath, B.; Culmsee, C.; Dolga, A.M. One protein, different cell fate: The differential outcome of depleting GRP75 during oxidative stress in neurons. Cell Death Dis. 2018, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Altamimi, T.R.; Karwi, Q.G.; Uddin, G.M.; Fukushima, A.; Kwong, J.Q.; Molkentin, J.D.; Lopaschuk, G.D. Cardiac-specific deficiency of the mitochondrial calcium uniporter augments fatty acid oxidation and functional reserve. J. Mol. Cell. Cardiol. 2019, 127, 223–231. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bencurova, M.; Lysikova, T.; Leskova Majdova, K.; Kaplan, P.; Racay, P.; Lehotsky, J.; Tatarkova, Z. Age-Dependent Changes in Calcium Regulation after Myocardial Ischemia–Reperfusion Injury. Biomedicines 2023, 11, 1193. https://doi.org/10.3390/biomedicines11041193
Bencurova M, Lysikova T, Leskova Majdova K, Kaplan P, Racay P, Lehotsky J, Tatarkova Z. Age-Dependent Changes in Calcium Regulation after Myocardial Ischemia–Reperfusion Injury. Biomedicines. 2023; 11(4):1193. https://doi.org/10.3390/biomedicines11041193
Chicago/Turabian StyleBencurova, Maria, Terezia Lysikova, Katarina Leskova Majdova, Peter Kaplan, Peter Racay, Jan Lehotsky, and Zuzana Tatarkova. 2023. "Age-Dependent Changes in Calcium Regulation after Myocardial Ischemia–Reperfusion Injury" Biomedicines 11, no. 4: 1193. https://doi.org/10.3390/biomedicines11041193