
Transcriptomic Approaches in Studies on and Applications of Chimeric Antigen Receptor T Cells



Abstract
:1. Introduction
2. Methods
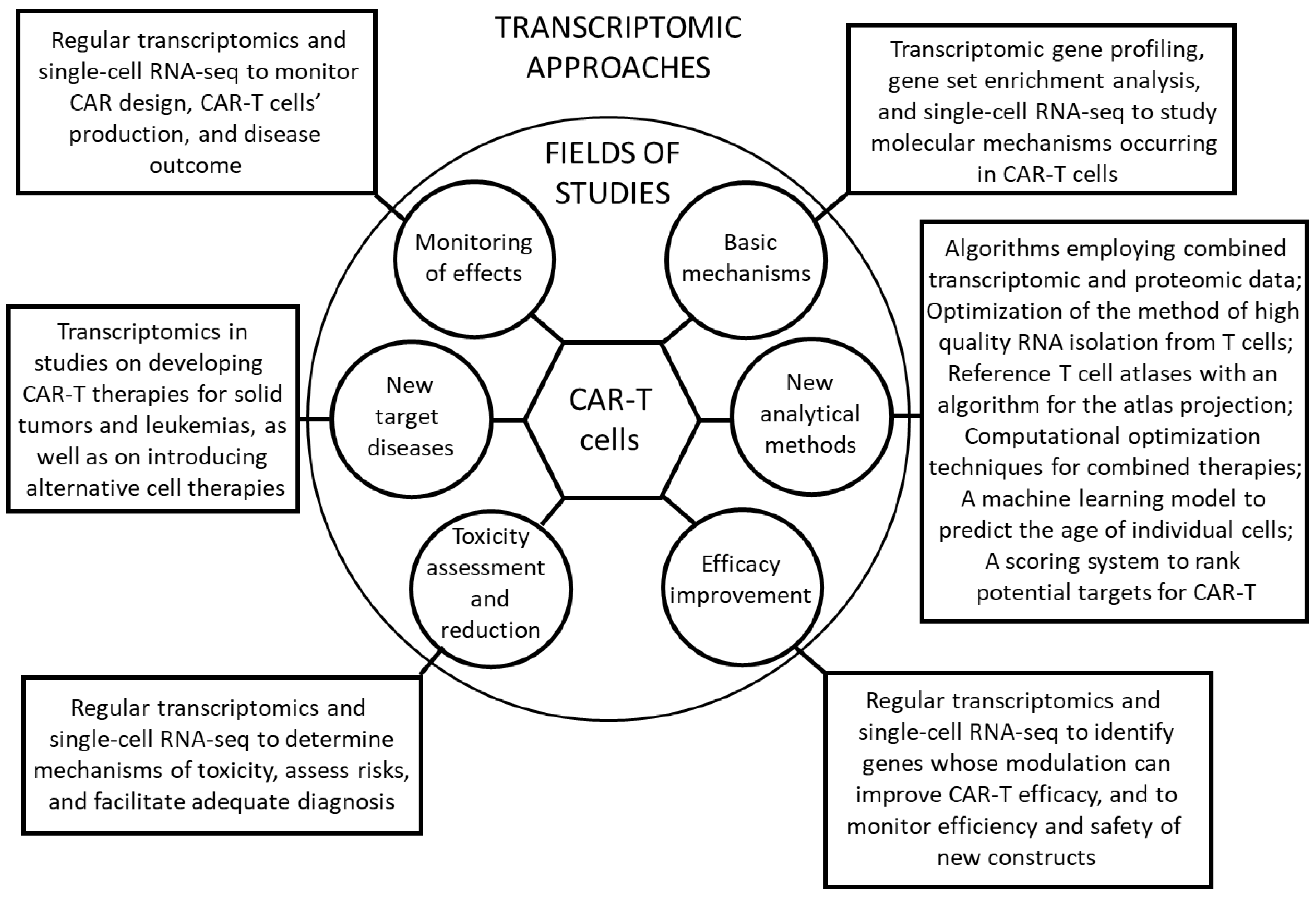
3. Transcriptomics in Elucidating Basic Mechanisms of CAR-T Functions
4. Transcriptomics in Developing New Methods of CAR-T Analyses
5. Transcriptomics in Improving the Efficacy of CAR-T Therapies
6. Transcriptomics in Assessing and Reducing the Toxicity of CAR-T Cells
7. Transcriptomics in Identifying New Target Diseases for CAR-T Therapies
8. Transcriptomics in Monitoring the Efficacy of CAR-T Therapies
9. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ślebioda, T.J.; Stanisławowski, M.; Kaszubowska, L.; Zaucha, J.M.; Żmijewski, M.A. Current and Future Perspectives for Chimeric Antigen Receptor T Cells Development in Poland. Biomedicines 2022, 10, 2912. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Abila, B.; Mostafa Kamel, Y. CAR-T: What Is Next? Cancers 2023, 15, 663. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, Ł.; Pierzynowska, K.; Gaffke, L.; Cyske, Z.; Mincewicz, G.; Węgrzyn, G. The use of phage display systems to combat infectious diseases in poultry: Diagnostic, vaccine, and therapeutic approaches. J. Appl. Microbiol. 2023, 134, lxac012. [Google Scholar] [CrossRef]
- Yang, J.; Chen, Y.; Jing, Y.; Green, M.R.; Han, L. Advancing CAR T cell therapy through the use of multidimensional omics data. Nat. Rev. Clin. Oncol. 2023; in press. [Google Scholar] [CrossRef]
- Vandereyken, K.; Sifrim, A.; Thienpont, B.; Voet, T. Methods and applications for single-cell and spatial multi-omics. Nat. Rev. Genet. 2023; in press. [Google Scholar] [CrossRef]
- Gabig, M.; Wegrzyn, G. An introduction to DNA chips: Principles, technology, applications and analysis. Acta Biochim. Pol. 2001, 48, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Moskot, M.; Montefusco, S.; Jakóbkiewicz-Banecka, J.; Mozolewski, P.; Węgrzyn, A.; Di Bernardo, D.; Węgrzyn, G.; Medina, D.L.; Ballabio, A.; Gabig-Cimińska, M. The phytoestrogen genistein modulates lysosomal metabolism and transcription factor EB (TFEB) activation. J. Biol. Chem. 2014, 289, 17054–17069. [Google Scholar] [CrossRef] [Green Version]
- Gaffke, L.; Pierzynowska, K.; Podlacha, M.; Hoinkis, D.; Rintz, E.; Brokowska, J.; Cyske, Z.; Wegrzyn, G. Underestimated Aspect of Mucopolysaccharidosis Pathogenesis: Global Changes in Cellular Processes Revealed by Transcriptomic Studies. Int. J. Mol. Sci. 2020, 21, 1204. [Google Scholar] [CrossRef] [Green Version]
- Wiśniewska, K.; Gaffke, L.; Krzelowska, K.; Węgrzyn, G.; Pierzynowska, K. Differences in gene expression patterns, revealed by RNA-seq analysis, between various Sanfilippo and Morquio disease subtypes. Gene 2022, 812, 146090. [Google Scholar] [CrossRef] [PubMed]
- Brokowska, J.; Gaffke, L.; Pierzynowska, K.; Cyske, Z.; Węgrzyn, G. Cell cycle disturbances in mucopolysaccharidoses: Transcriptomic and experimental studies on cellular models. Exp. Biol. Med. 2022, 247, 1639–1649. [Google Scholar] [CrossRef]
- Ryu, Y.; Han, G.H.; Jung, E.; Hwang, D. Integration of Single-Cell RNA-Seq Datasets: A Review of Computational Methods. Mol. Cells 2023, 46, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Fraietta, J.A.; Lacey, S.F.; Orlando, E.J.; Pruteanu-Malinici, I.; Gohil, M.; Lundh, S.; Boesteanu, A.C.; Wang, Y.; O’Connor, R.S.; Hwang, W.-T.; et al. Determinants of response and resistance to CD19 chimeric antigen receptor (CAR) T cell therapy of chronic lymphocytic leukemia. Nat. Med. 2018, 24, 563–571. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, H.; Chen, S.Y.; Liu, C.J.; Hu, F.F.; Yu, J.; Wu, Y.; Guo, A.Y. Transcriptome and Regulatory Network Analyses of CD19-CAR-T Immunotherapy for B-ALL. Genom. Proteom. Bioinform. 2019, 17, 190–200. [Google Scholar] [CrossRef]
- Sheih, A.; Voillet, V.; Hanafi, L.A.; DeBerg, H.A.; Yajima, M.; Hawkins, R.; Gersuk, V.; Riddell, S.R.; Maloney, D.G.; Wohlfahrt, M.E.; et al. Clonal kinetics and single-cell transcriptional profiling of CAR-T cells in patients undergoing CD19 CAR-T immunotherapy. Nat. Commun. 2020, 11, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boroughs, A.C.; Larson, R.C.; Marjanovic, N.D.; Gosik, K.; Castano, A.P.; Porter, C.B.M.; Lorrey, S.J.; Ashenberg, O.; Jerby, L.; Hofree, M.; et al. A Distinct Transcriptional Program in Human CAR T Cells Bearing the 4-1BB Signaling Domain Revealed by scRNA-Seq. Mol. Ther. 2020, 28, 2577–2592. [Google Scholar] [CrossRef]
- Adnan Awad, S.; Dufva, O.; Ianevski, A.; Ghimire, B.; Koski, J.; Maliniemi, P.; Thomson, D.; Schreiber, A.; Heckman, C.A.; Koskenvesa, P.; et al. RUNX1 mutations in blast-phase chronic myeloid leukemia associate with distinct phenotypes, transcriptional profiles, and drug responses. Leukemia 2021, 35, 1087–1099. [Google Scholar] [CrossRef]
- Li, X.; Guo, X.; Zhu, Y.; Wei, G.; Zhang, Y.; Li, X.; Xu, H.; Cui, J.; Wu, W.; He, J.; et al. Single-Cell Transcriptomic Analysis Reveals BCMA CAR-T Cell Dynamics in a Patient with Refractory Primary Plasma Cell Leukemia. Mol. Ther. 2021, 29, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Peticone, C.; Kotsopoulou, E.; Göttgens, B.; Calero-Nieto, F.J. Single-cell transcriptome analysis of CAR T-cell products reveals subpopulations, stimulation, and exhaustion signatures. Oncoimmunology 2021, 10, 1866287. [Google Scholar] [CrossRef] [PubMed]
- Gennert, D.G.; Lynn, R.C.; Granja, J.M.; Weber, E.W.; Mumbach, M.R.; Zhao, Y.; Duren, Z.; Sotillo, E.; Greenleaf, W.J.; Wong, W.H.; et al. Dynamic chromatin regulatory landscape of human CAR T cell exhaustion. Proc. Natl. Acad. Sci. USA 2021, 118, e2104758118. [Google Scholar] [CrossRef] [PubMed]
- Ung, S.; Choochuen, P.; Khopanlert, W.; Maneechai, K.; Sangkhathat, S.; Terakura, S.; Julamanee, J. Enrichment of T-cell proliferation and memory gene signatures of CD79A/CD40 costimulatory domain potentiates CD19CAR-T cell functions. Front. Immunol. 2022, 13, 1064339. [Google Scholar] [CrossRef]
- Panetti, S.; McJannett, N.J.; Fultang, L.; Booth, S.; Gneo, L.; Scarpa, U.; Smith, C.; Vardon, A.; Vettore, L.; Whalley, C.; et al. Engineering amino acid uptake or catabolism promotes CAR-T cell adaption to the tumour environment. Blood Adv. 2023; in press. [Google Scholar] [CrossRef]
- Huang, B.; Lang, X.; Li, X. The role of IL-6/JAK2/STAT3 signaling pathway in cancers. Front. Oncol. 2023, 12, 1023177. [Google Scholar] [CrossRef]
- Kerr, J.S.; Wilson, C.H. Nuclear receptor-binding protein 1: A novel tumour suppressor and pseudokinase. Biochem. Soc. Transact. 2013, 41, 1055–1060. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Q.; Zhang, Y.; Han, J. The Role of Histone Modification in DNA Replication-Coupled Nucleosome Assembly and Cancer. Int. J. Mol. Sci. 2023, 24, 4939. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shahir, M.; Taheri, M.; Salimi, A. A review on the role of chemokines in the pathogenesis of systemic lupus erythematosus. Cytokine 2021, 146, 155640. [Google Scholar] [CrossRef] [PubMed]
- Gicobi, J.K.; Dellacecca, E.R.; Dong, H. Resilient T-cell responses in patients with advanced cancers. Int. J. Hematol. 2022; in press. [Google Scholar] [CrossRef]
- Borza, R.; Salgado-Polo, F.; Moolenaar, W.H.; Perrakis, A. Structure and function of the ecto-nucleotide pyrophosphatase/phosphodiesterase (ENPP) family: Tidying up diversity. J. Biol. Chem. 2022, 298, 101526. [Google Scholar] [CrossRef] [PubMed]
- Mesas-Fernández, A.; Bodner, E.; Hilke, F.J.; Meier, K.; Ghoreschi, K.; Solimani, F. Interleukin-21 in autoimmune and inflammatory skin diseases. Eur. J. Immunol. 2023; e2250075, in press. [Google Scholar] [CrossRef]
- Han, J.; Correa da Rosa, J.; Agarwal, A.; Owji, S.; Yassky, D.; Luu, Y.; Shah, A.; Estrada, Y.; Ungar, J.; Sarin, K.Y.; et al. Modulation of Inflammatory Proteins in Serum May Reflect Cutaneous Immune Responses in Cancer Immunotherapy. JID Innov. 2023, 3, 100179. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhu, H.; Chen, B.; He, X.; Shen, Y.; Zhang, X.; Chen, W.; Liu, X.; Xu, Y.; Xu, X. Tubulin Alpha 1b Is Associated with the Immune Cell Infiltration and the Response of HCC Patients to Immunotherapy. Diagnostics 2022, 12, 858. [Google Scholar] [CrossRef]
- Chen, D.P.; Wen, Y.H.; Wang, W.T.; Lin, W.T. Exploring the Bio-Functional Effect of Single Nucleotide Polymorphisms in the Promoter Region of the TNFSF4, CD28, and PDCD1 Genes. J. Clin. Med. 2023, 12, 2157. [Google Scholar] [CrossRef]
- Schütz, C.; Baraliakos, X. What do we know about co-stimulatory and co-inhibitory immune checkpoint signals in ankylosing spondylitis? Clin. Exp. Immunol. 2023; uxad032, in press. [Google Scholar] [CrossRef]
- Koh, J.; Jang, I.; Mun, S.; Lee, C.; Cha, H.J.; Oh, Y.H.; Kim, J.M.; Han, J.H.; Paik, J.H.; Cho, J.; et al. Genetic profiles of subcutaneous panniculitis-like T-cell lymphoma and clinicopathological impact of HAVCR2 mutations. Blood Adv. 2021, 5, 3919–3930. [Google Scholar] [CrossRef]
- Fukasawa, M.; Nishio, K.; Oikawa, D.; Itou, T.; Iinuma, T.; Asano, M. Allograft inflammatory factor-1 released from the cerebral microglia affect several organs in the body. J. Mol. Histol. 2023; in press. [Google Scholar] [CrossRef]
- Shapiro, I.E.; Bassani-Sternberg, M. The impact of immunopeptidomics: From basic research to clinical implementation. Semin. Immunol. 2023, 66, 101727. [Google Scholar] [CrossRef]
- Côrte-Real, J.V.; Baldauf, H.M.; Melo-Ferreira, J.; Abrantes, J.; Esteves, P.J. Evolution of Guanylate Binding Protein (GBP) Genes in Muroid Rodents (Muridae and Cricetidae) Reveals an Outstanding Pattern of Gain and Loss. Front. Immunol. 2022, 13, 752186. [Google Scholar] [CrossRef]
- Wu, H.; Wang, Y.; Chen, X.; Yao, Y.; Zhao, W.; Fang, L.; Sun, X.; Wang, N.; Jiang, J.; Gao, L.; et al. Cell-Dependent Pathogenic Roles of Filamin B in Different Skeletal Malformations. Oxid. Med. Cell. Longev. 2022, 2022, 8956636. [Google Scholar] [CrossRef]
- Perna, F.; Berman, S.H.; Soni, R.K.; Mansilla-Soto, J.; Eyquem, J.; Hamieh, M.; Hendrickson, R.C.; Brennan, C.W.; Sadelain, M. Integrating Proteomics and Transcriptomics for Systematic Combinatorial Chimeric Antigen Receptor Therapy of AML. Cancer Cell. 2017, 32, 506–519.e5. [Google Scholar] [CrossRef] [Green Version]
- DeLucia, D.C.; Lee, J.K. Identification of Cell Surface Targets for CAR T Cell Immunotherapy. Methods Mol. Biol. 2020, 2097, 45–54. [Google Scholar]
- Healy, Z.R.; Weinhold, K.J.; Murdoch, D.M. Transcriptional Profiling of CD8+ CMV-Specific T Cell Functional Subsets Obtained Using a Modified Method for Isolating High-Quality RNA From Fixed and Permeabilized Cells. Front. Immunol. 2020, 11, 1859. [Google Scholar] [CrossRef] [PubMed]
- Andreatta, M.; Corria-Osorio, J.; Müller, S.; Cubas, R.; Coukos, G.; Carmona, S.J. Interpretation of T cell states from single-cell transcriptomics data using reference atlases. Nat. Commun. 2021, 12, 2965. [Google Scholar] [CrossRef]
- Ahmadi, S.; Sukprasert, P.; Vegesna, R.; Sinha, S.; Schischlik, F.; Artzi, N.; Khuller, S.; Schäffer, A.A.; Ruppin, E. The landscape of receptor-mediated precision cancer combination therapy via a single-cell perspective. Nat. Commun. 2022, 13, 1613. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Ahmad, R.; Nguyen, T.; Cifello, J.; Hemani, H.; Li, J.; Chen, J.; Li, S.; Wang, J.; Achour, A.; et al. Heterogeneity and transcriptome changes of human CD8+ T cells across nine decades of life. Nat. Commun. 2022, 13, 5128. [Google Scholar] [CrossRef]
- Patiño-Escobar, B.; Ferguson, I.D.; Wiita, A.P. Unraveling the surface proteomic profile of multiple myeloma to reveal new immunotherapeutic targets and markers of drug resistance. Cell. Stress 2022, 6, 89–92. [Google Scholar] [CrossRef]
- Jiang, Z.; Liao, R.; Lv, J.; Li, S.; Zheng, D.; Qin, L.; Wu, D.; Chen, S.; Long, Y.; Wu, Q.; et al. IL-6 trans-signaling promotes the expansion and anti-tumor activity of CAR T cells. Leukemia 2021, 35, 1380–1391. [Google Scholar] [CrossRef]
- Bandey, I.N.; Adolacion, J.R.T.; Romain, G.; Paniagua, M.M.; An, X.; Saeedi, A.; Liadi, I.; You, Z.; Rajanayake, R.B.; Hwu, P.; et al. Designed improvement to T-cell immunotherapy by multidimensional single cell profiling. J. Immunother. Cancer 2021, 9, e001877. [Google Scholar] [CrossRef]
- Xia, L.; Zheng, Z.; Liu, J.Y.; Chen, Y.J.; Ding, J.; Hu, G.S.; Hu, Y.H.; Liu, S.; Luo, W.X.; Xia, N.S.; et al. Targeting Triple-Negative Breast Cancer with Combination Therapy of EGFR CAR T Cells and CDK7 Inhibition. Cancer Immunol. Res. 2021, 9, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Sutra Del Galy, A.; Menegatti, S.; Fuentealba, J.; Lucibello, F.; Perrin, L.; Helft, J.; Darbois, A.; Saitakis, M.; Tosello, J.; Rookhuizen, D.; et al. In vivo genome-wide CRISPR screens identify SOCS1 as intrinsic checkpoint of CD4+ TH1 cell response. Sci. Immunol. 2021, 6, eabe8219. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, H.; Dunne, S.; Martin, D.S.; Kavanagh, H.; Dunne, S.; Martin, D.S.; McFadden, E.; Gallagher, L.; Schwaber, J.; Leonard, S.; et al. A novel non-viral delivery method that enables efficient engineering of primary human T cells for ex vivo cell therapy applications. Cytotherapy 2021, 23, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Park, J.J.; Peng, L.; Yang, Q.; Chow, R.D.; Dong, M.B.; Lam, S.Z.; Guo, J.; Tang, E.; Zhang, Y.; et al. A genome-scale gain-of-function CRISPR screen in CD8 T cells identifies proline metabolism as a means to enhance CAR-T therapy. Cell. Metab. 2022, 34, 595–614.e14. [Google Scholar] [CrossRef] [PubMed]
- Wirges, A.; Bunse, M.; Joedicke, J.J.; Blanc, E.; Gudipati, V.; Moles, M.W.; Shiku, H.; Beule, D.; Huppa, J.B.; Höpken, U.E.; et al. EBAG9 silencing exerts an immune checkpoint function without aggravating adverse effects. Mol. Ther. 2022, 30, 3358–3378. [Google Scholar] [CrossRef]
- Michelozzi, I.M.; Gomez-Castaneda, E.; Pohle, R.V.C.; Cardoso Rodriguez, F.; Sufi, J.; Puigdevall-Costa, P.; Subramaniyam, M.; Kirtsios, E.; Eddaoudi, A.; Wu, S.W.; et al. Activation priming and cytokine polyfunctionality modulate the enhanced functionality of low-affinity CD19 CAR T cells. Blood Adv. 2023; in press. [Google Scholar] [CrossRef]
- Li, R.; Li, D.; Nie, Y. IL-6/gp130 signaling: A key unlocking regeneration. Cell. Regen. 2023, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Ugolini, A.; Nuti, M. CD137+ T-Cells: Protagonists of the Immunotherapy Revolution. Cancers 2021, 13, 456. [Google Scholar] [CrossRef]
- Li, Z.M.; Liu, G.; Gao, Y.; Zhao, M.G. Targeting CDK7 in oncology: The avenue forward. Pharmacol. Ther. 2022, 240, 108229. [Google Scholar] [CrossRef]
- Sobah, M.L.; Liongue, C.; Ward, A.C. SOCS Proteins in Immunity, Inflammatory Diseases, and Immune-Related Cancer. Front. Med. 2021, 8, 727987. [Google Scholar] [CrossRef]
- Rehm, A.; Wirges, A.; Hoser, D.; Fischer, C.; Herda, S.; Gerlach, K.; Sauer, S.; Willimsky, G.; Höpken, U.E. EBAG9 controls CD8+ T cell memory formation responding to tumor challenge in mice. JCI Insight 2022, 7, e155534. [Google Scholar] [CrossRef]
- Zhu, X.; Hu, H.; Xiao, Y.; Li, Q.; Zhong, Z.; Yang, J.; Zou, P.; Cao, Y.; Meng, F.; Li, W.; et al. Tumor-derived extracellular vesicles induce invalid cytokine release and exhaustion of CD19 CAR-T Cells. Cancer Lett. 2022, 536, 215668. [Google Scholar] [CrossRef]
- Loeffler-Wirth, H.; Rade, M.; Arakelyan, A.; Kreuz, M.; Loeffler, M.; Koehl, U.; Reiche, K.; Binder, H. Transcriptional states of CAR-T infusion relate to neurotoxicity—Lessons from high-resolution single-cell SOM expression portraying. Front. Immunol. 2022, 13, 994885. [Google Scholar] [CrossRef]
- Wen, L.; Huang, Y.; Peng, L.; Zhao, K.; Sun, Y.; Lin, Z.; Chen, Y.; Li, Z.; Qian, Q.; Tong, F.; et al. Single-cell RNA-Seq reveals the potential risk of anti-mesothelin CAR T Cell therapy toxicity to different organs in humans. Front. Immunol. 2022, 13, 807915. [Google Scholar] [CrossRef] [PubMed]
- Leipold, A.M.; Werner, R.A.; Düll, J.; Jung, P.; John, M.; Stanojkovska, E.; Zhou, X.; Hornburger, H.; Ruckdeschel, A.; Dietrich, O.; et al. Th17.1 cell driven sarcoidosis-like inflammation after anti-BCMA CAR T cells in multiple myeloma. Leukemia 2023, 37, 650–658. [Google Scholar] [CrossRef]
- Prinzing, B.; Schreiner, P.; Bell, M.; Fan, Y.; Krenciute, G.; Gottschalk, S. MyD88/CD40 signaling retains CAR T cells in a less differentiated state. JCI Insight 2020, 5, e136093. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, N.; Long, M.; Fushimi, A.; Yamamoto, M.; Hata, T.; Hagiwara, M.; Bhattacharya, A.; Hu, Q.; Wong, K.K.; Liu, S.; et al. MUC1-C integrates activation of the IFN-γ pathway with suppression of the tumor immune microenvironment in triple-negative breast cancer. J. Immunother. Cancer 2021, 9, e002115. [Google Scholar] [CrossRef]
- Konduri, V.; Joseph, S.K.; Byrd, T.T.; Nawas, Z.; Vazquez-Perez, J.; Hofferek, C.J.; Halpert, M.M.; Liu, D.; Liang, Z.; Baig, Y.; et al. A subset of cytotoxic effector memory T cells enhances CAR T cell efficacy in a model of pancreatic ductal adenocarcinoma. Sci. Transl. Med. 2021, 13, eabc3196. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Liu, J.Y.; Zheng, Z.Z.; Chen, Y.J.; Ding, J.C.; Hu, Y.H.; Hu, G.S.; Xia, N.S.; Liu, W. BRD4 inhibition boosts the therapeutic effects of epidermal growth factor receptor-targeted chimeric antigen receptor T cells in glioblastoma. Mol. Ther. 2021, 29, 3011–3026. [Google Scholar] [CrossRef]
- Le, Q.; Hadland, B.; Smith, J.L.; Leonti, A.; Huang, B.J.; Ries, R.; Hylkema, T.A.; Castro, S.; Tang, T.T.; McKay, C.N.; et al. CBFA2T3-GLIS2 model of pediatric acute megakaryoblastic leukemia identifies FOLR1 as a CAR T cell target. J. Clin. Invest. 2022, 132, e157101. [Google Scholar] [CrossRef]
- Sottile, R.; Panjwani, M.K.; Lau, C.M.; Daniyan, A.F.; Tanaka, K.; Barker, J.N.; Brentjens, R.J.; Sun, J.C.; Le Luduec, J.B.; Hsu, K.C. Human cytomegalovirus expands a CD8+ T cell population with loss of BCL11B expression and gain of NK cell identity. Sci. Immunol. 2021, 6, eabe6968. [Google Scholar] [CrossRef]
- Cicirò, Y.; Sala, A. MYB oncoproteins: Emerging players and potential therapeutic targets in human cancer. Oncogenesis 2021, 10, 19. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, P.; Ahmad, A.; Fatima, M.; Nasser, M.W. FOXM1: A small fox that makes more tracks for cancer progression and metastasis. Semin. Cancer Biol. 2023, 92, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Spinner, C.A.; Lazarevic, V. Transcriptional regulation of adaptive and innate lymphoid lineage specification. Immunol. Rev. 2021, 300, 65–81. [Google Scholar] [CrossRef]
- Kufe, D.W. MUC1-C in chronic inflammation and carcinogenesis; emergence as a target for cancer treatment. Carcinogenesis 2020, 41, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Nüssing, S.; Sutton, V.R.; Trapani, J.A.; Parish, I.A. Beyond target cell death—Granzyme serine proteases in health and disease. Mol. Aspects Med. 2022, 88, 101152. [Google Scholar] [CrossRef] [PubMed]
- Martella, N.; Pensabene, D.; Varone, M.; Colardo, M.; Petraroia, M.; Sergio, W.; La Rosa, P.; Moreno, S.; Segatto, M. Bromodomain and Extra-Terminal Proteins in Brain Physiology and Pathology: BET-ing on Epigenetic Regulation. Biomedicines 2023, 11, 750. [Google Scholar] [CrossRef]
- Young, O.; Ngo, N.; Lin, L.; Stanbery, L.; Creeden, J.F.; Hamouda, D.; Nemunaitis, J. Folate Receptor as a Biomarker and Therapeutic Target in Solid Tumors. Curr. Probl. Cancer 2023, 47, 100917. [Google Scholar] [CrossRef]
- Charitidis, F.T.; Adabi, E.; Thalheimer, F.B.; Clarke, C.; Buchholz, C.J. Monitoring CAR T cell generation with a CD8-targeted lentiviral vector by single-cell transcriptomics. Mol. Ther. Methods Clin. Dev. 2021, 23, 359–369. [Google Scholar] [CrossRef]
- Deng, Q.; Han, G.; Puebla-Osorio, N.; Ma, M.C.J.; Strati, P.; Chasen, B.; Dai, E.; Dang, M.; Jain, N.; Yang, H.; et al. Characteristics of anti-CD19 CAR T cell infusion products associated with efficacy and toxicity in patients with large B cell lymphomas. Nat. Med. 2020, 26, 1878–1887. [Google Scholar] [CrossRef]
- Castellanos-Rueda, R.; Di Roberto, R.B.; Schlatter, F.S.; Reddy, S.T. Leveraging Single-Cell Sequencing for Chimeric Antigen Receptor T Cell Therapies. Trends Biotechnol. 2021, 39, 1308–1320. [Google Scholar] [CrossRef]
- Tumino, N.; Weber, G.; Besi, F.; Del Bufalo, F.; Bertaina, V.; Paci, P.; Quatrini, L.; Antonucci, L.; Sinibaldi, M.; Quintarelli, C.; et al. Polymorphonuclear myeloid-derived suppressor cells impair the anti-tumor efficacy of GD2.CAR T-cells in patients with neuroblastoma. J. Hematol. Oncol. 2021, 14, 191. [Google Scholar] [CrossRef]
- Shao, L.; Iyer, A.; Zhao, Y.; Somerville, R.; Panch, S.; Pelayo, A.; Stroncek, D.F.; Jin, P. Identification of genomic signatures in bone marrow associated with clinical response of CD19 CAR T-cell therapy. Sci. Rep. 2022, 12, 2830. [Google Scholar] [CrossRef]
- Chen, W.; Shi, H.; Liu, Z.; Yang, F.; Liu, J.; Zhang, L.; Wu, Y.; Xia, Y.; Ou, Y.; Li, R.; et al. Single-cell transcriptomics reveals immune reconstitution in R/R T-ALL/LBL patients with donor-derived CD7 CAR-T therapy. Clin. Cancer Res. 2023; in press. [Google Scholar] [CrossRef]
- Nolan, E.M.; Peet, J.J.Y. Post-translational modifications on the metal-sequestering protein calprotectin. Biometals, 2023; in press. [Google Scholar] [CrossRef]
- Wolf, M.; Joseph, R.; Austermann, J.; Scharrnbeck-Davis, C.; Hermann, S.; Roth, J.; Vogl, T. S100A8/S100A9 Integrates F-Actin and Microtubule Dynamics to Prevent Uncontrolled Extravasation of Leukocytes. Biomedicines 2023, 11, 835. [Google Scholar] [CrossRef] [PubMed]
- Garcia, V.; Perera, Y.R.; Chazin, W.J. A Structural Perspective on Calprotectin as a Ligand of Receptors Mediating Inflammation and Potential Drug Target. Biomolecules 2022, 12, 519. [Google Scholar] [CrossRef] [PubMed]
- Patel, U.; Abernathy, J.; Savani, B.N.; Oluwole, O.; Sengsayadeth, S.; Dholaria, B. CAR T cell therapy in solid tumors: A review of current clinical trials. EJHaem 2021, 3 (Suppl. S1), 24–31. [Google Scholar] [CrossRef]
- Cyske, Z.; Anikiej-Wiczenbach, P.; Wisniewska, K.; Wisniewska, K.; Gaffke, L.; Pierzynowska, K.; Mański, A.; Wegrzyn, G. Sanfilippo Syndrome: Optimizing Care with a Multidisciplinary Approach. J. Multidiscip. Healthc. 2022, 15, 2097–2110. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Process Affected by or in CAR-T Cells | Examples of Genes with Changed Expression | Function(s) of the Gene(s) or Gene Product(s) |
|---|---|---|
| Immunological memory | IL6, STAT3 [12] | Interleukin 6 (IL-6) is a proinflammatory cytokine; STAT3 is a transcription activator, stimulated in response to action of cytokines [22]. |
| Cell cycle and immune response | NRBP1, HIST1H4A, HIST2H4A [13] | Nuclear receptor-binding protein 1 (NRBP1) is a tumor suppressor and pseudokinase [23]; HIST1H4A, HIST2H4A encode histone proteins [24]. |
| Cytotoxicity and proliferation | CCL4, NKG7 [14] | CCL4 is a chemokine [25]; NKG7 is a natural killer cell granule protein [26]. |
| Human leukocyte antigens’ abundance | ENPP2, IL21 [15] | Ectonucleotide pyrophosphatase/phosphodiesterase 2 (ENPP2) indirectly stimulates cell proliferation and chemotaxis [27]; interleukin 21 (IL-21) is a cytokine with immunoregulatory activity [28]. |
| Cell proliferation, cytotoxicity, and signal transduction | TNFRSF4, TUBA1B, RRM2, CCL3, CCL4, CCL5 [17] | TNFRSF4 is a member of the TNF-receptor superfamily which activates NF-κB [29]; TUBA1B codes for tubulin α1b [30]; CCL3, CCL4, and CCL5 are chemokines [25]. |
| Exhaustion | PDCD1, CTLA4, HAVCR2 [19] | Programmed cell death protein 1 (PDCD1) is an immune-inhibitory receptor [31]; cytotoxic T-lymphocyte-associated protein 4 (CTLA4) transmits an inhibitory signal to T cells [32]; HAVCR2 is a cell surface receptor with an inhibitory function [33]. |
| Proliferation and interferon signaling pathway | AIF1, HLA-DP, HLA-DR, HLA-DM, IFNGR2, GBP6, FLNB [20] | Allograft inflammatory factor-1 (AIF1) is a protein that binds actin and calcium, and stimulates proliferation [34]; HLA-DP, HLA-DR, and HLA-DM are proteins from the major histocompatibility complex class II [35]; guanylate-binding protein 6 (GBP6) is induced by interferon and hydrolyzes GTP to both GDP and GMP [36]; filamin B (FLNB) is a protein connecting cell membrane to the actin cytoskeleton [37]. |
| Problem to Be Solved | Principle of the Method | Application(s) |
|---|---|---|
| Identification of potential targets for CAR | Algorithm based on transcriptomic and proteomic data [38,39] | Development of novel CAR-T therapies |
| Obtaining high-quality RNA samples | A procedure involving fixation with aldehyde, permeabilization with a detergent, staining of intracellular cytokines, and sorting, combined with the addition of an RNase inhibitor and high-salt buffer [40] | All transcriptomic approaches, especially for CAR-T studies |
| Performing efficient analysis of transcriptomic data | Reference T cell atlases with an algorithm for reference atlas projection (ProjecTILs) [41]; a machine learning model [43] | Prediction of effects of various cellular dysfunctions; prediction of the age of individual cells |
| Ranking of potential therapeutic targets | A scoring system based on proteomic and transcriptomic data | Identification potential targets for CAR-T therapies |
| Target Process(es)/Approach to Improve the Efficacy | Gene to Be Modulated | Function of the Gene or Gene Product |
|---|---|---|
| T-cell migration and early memory differentiation | IL6ST [45] | IL-6ST is also known as glycoprotein 130 (GP130) which acts as a transmembrane cytokine receptor involved in the IL-6/STAT3 signal transduction pathway [53]. |
| Enhancement of tumor killing activity | CD137L (TNFSF9) [46] | CD137L is a ligand of the CD137 cytokine (from the tumor necrosis factor family) [54]. |
| Overcoming the resistance of cancer cells to EGFR-targeting CAR-T cells | CDK7 [47] | CDK7 is a cyclin-dependent kinase regulating the cell cycle progression [55]. |
| Checkpoint of the adoptive T-cell response | SOCS1 [48] | SOCS1 is a suppressor of cytokine signaling [56]. |
| Estrogen-dependent signal transduction pathway | EBAG9 [51] | EBAG9 is the estrogen receptor-binding fragment-associated antigen 9 which acts as a tumor-associated antigen, expressed with high efficiency in different kind of cancers [57]. |
| Assessed Toxicity | Specific Method | Application |
|---|---|---|
| Neurotoxicity | Single-cell RNA-seq with over 130,000 cells tested [59] | Identification of transcriptomic patterns suggesting molecular mechanisms of neurotoxicity and indicating possible ways to reduce the toxic effects |
| Toxicity to different organs | Single-cell RNA-seq with samples derived from various organs [60] | Assessment of the risk for tested organs to be toxified by CAR-T cells |
| Pulmonary flare-up | Single-cell RNA-seq [61] | Making a reliable diagnosis and distinguishing between the disease relapse and sarcoidosis |
| Target Disease | Target Gene | Function of the Target Gene or Gene Product |
|---|---|---|
| Glioma, B cell leukemia, osteosarcoma | MYB, FOXM1, TBET [62] | MYB is a proto-oncogene, a transcription factor involved in the control of hematopoiesis [68]; FOXM1 [69] and TBET (T-Bet) [70] are transcription factors involved in the regulation of cell proliferation and developmental processes, respectively. |
| Triple-negative breast cancer | MUC1-C [63] | MUC1-C is a protein from the mucin family, involved in protection of epithelia from the external environment [71]. |
| Pancreatic ductal adenocarcinoma | GZM gene family [64] | Granzymes are serine proteases secreted by natural killer cells and cytotoxic T lymphocytes [72]. |
| Glioblastoma | BRD4 [65] | BRD4 (bromodomain containing 4 protein) is capable of chromatin binding and stimulation of transcription of genes involved in cell cycle progression, differentiation, and inflammation [73]. |
| Acute myeloid leukemia | FOLR1 [66] | The FOLR1 gene codes for the folate receptor α that binds folic acid and transports 5-methyltetrahydrofolate into cells [74]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pierzynowska, K.; Gaffke, L.; Zaucha, J.M.; Węgrzyn, G. Transcriptomic Approaches in Studies on and Applications of Chimeric Antigen Receptor T Cells. Biomedicines 2023, 11, 1107. https://doi.org/10.3390/biomedicines11041107
Pierzynowska K, Gaffke L, Zaucha JM, Węgrzyn G. Transcriptomic Approaches in Studies on and Applications of Chimeric Antigen Receptor T Cells. Biomedicines. 2023; 11(4):1107. https://doi.org/10.3390/biomedicines11041107
Chicago/Turabian StylePierzynowska, Karolina, Lidia Gaffke, Jan M. Zaucha, and Grzegorz Węgrzyn. 2023. "Transcriptomic Approaches in Studies on and Applications of Chimeric Antigen Receptor T Cells" Biomedicines 11, no. 4: 1107. https://doi.org/10.3390/biomedicines11041107