
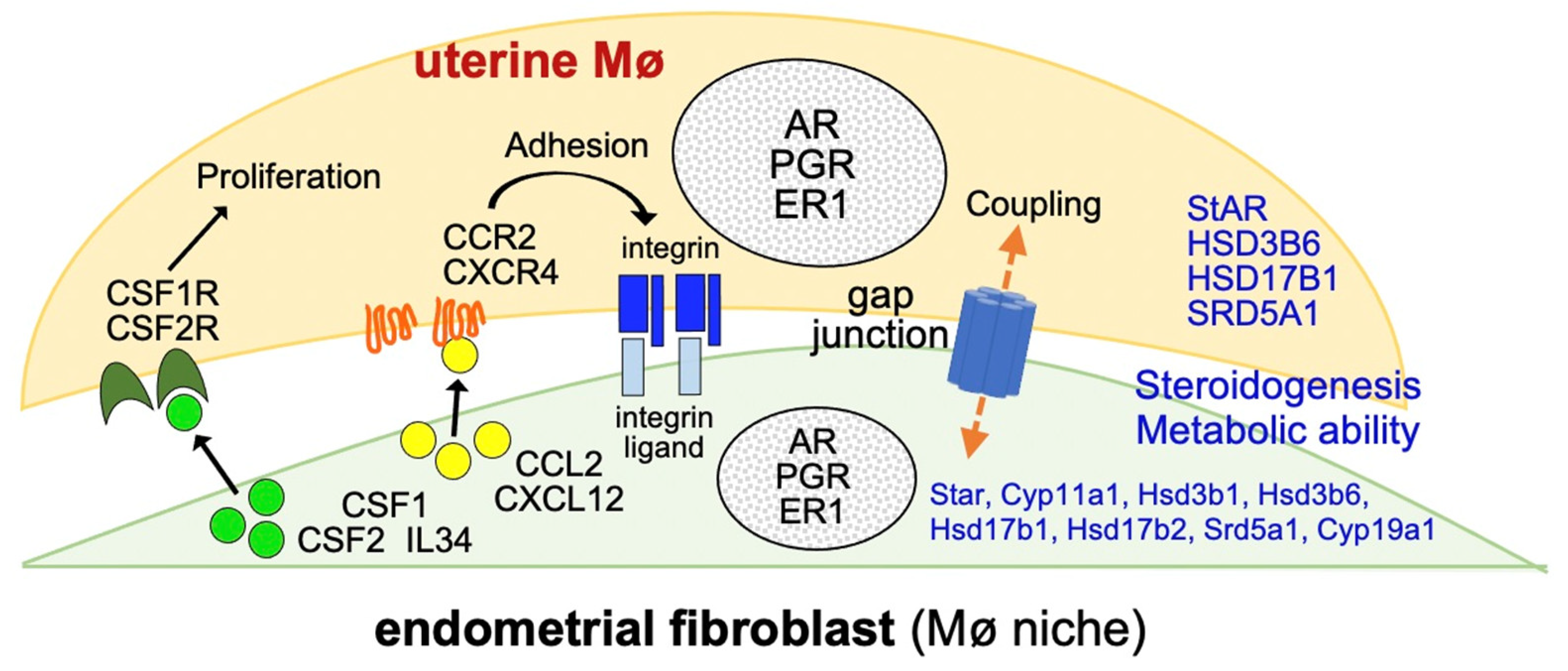
Mixed-Culture Propagation of Uterine-Tissue-Resident Macrophages and Their Expression Properties of Steroidogenic Molecules
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Propagation of Uterine Mø by Mixed Culture with Other Interstitial Cells from the Uterus
2.3. Separation of Uterine Mø Propagated by Mixed Culture with Other Uterine Interstitial Cells
2.4. Separation of Uterine Interstitial Cells Showing Fibroblastic Morphology from Uterine Mø
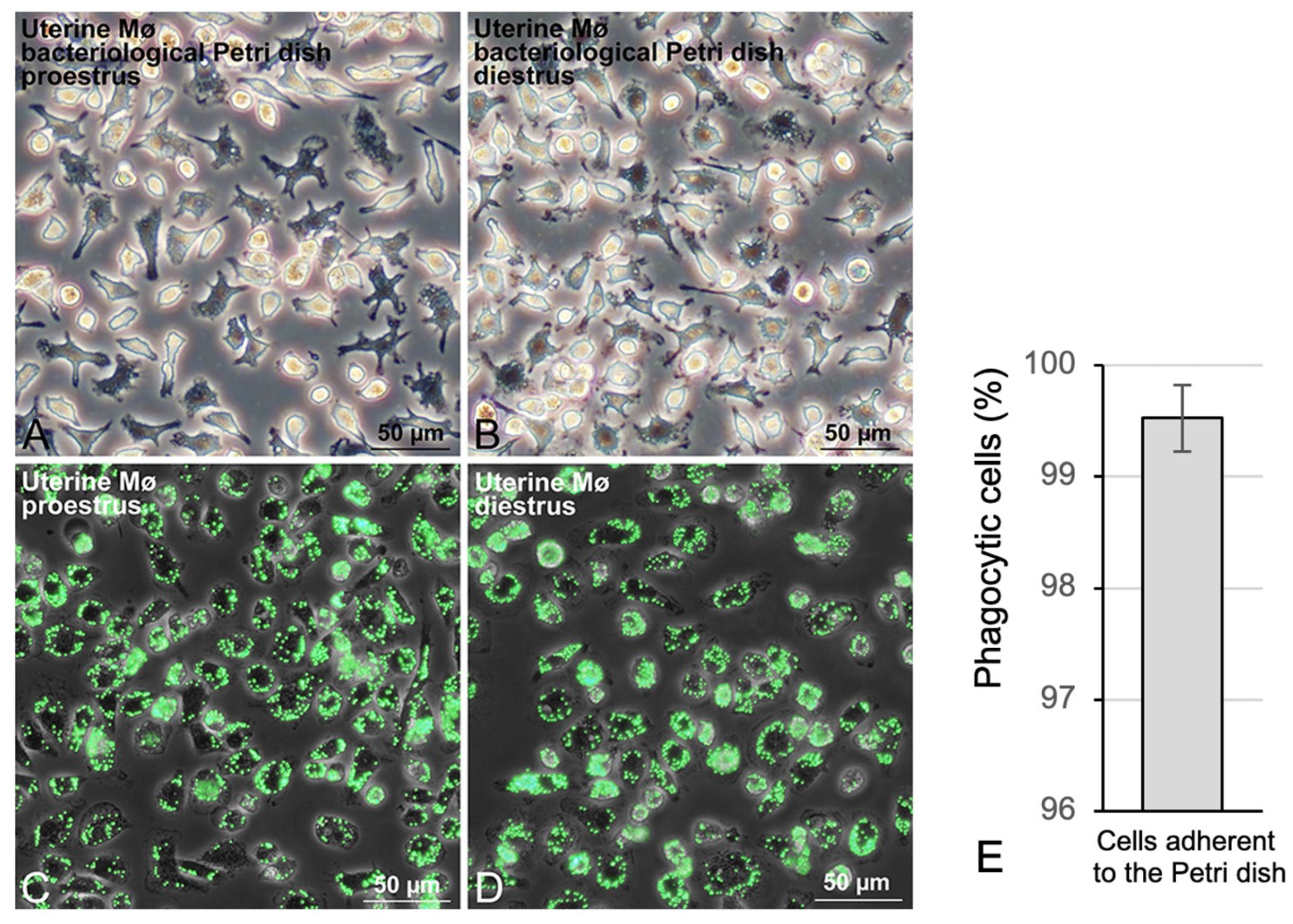
2.5. Phagocytosis Analysis of Uterine Mø with Fluorescent Beads
2.6. Flow Cytometry
2.7. Immunofluorescence Staining
2.8. RT-PCR Analyses
3. Results
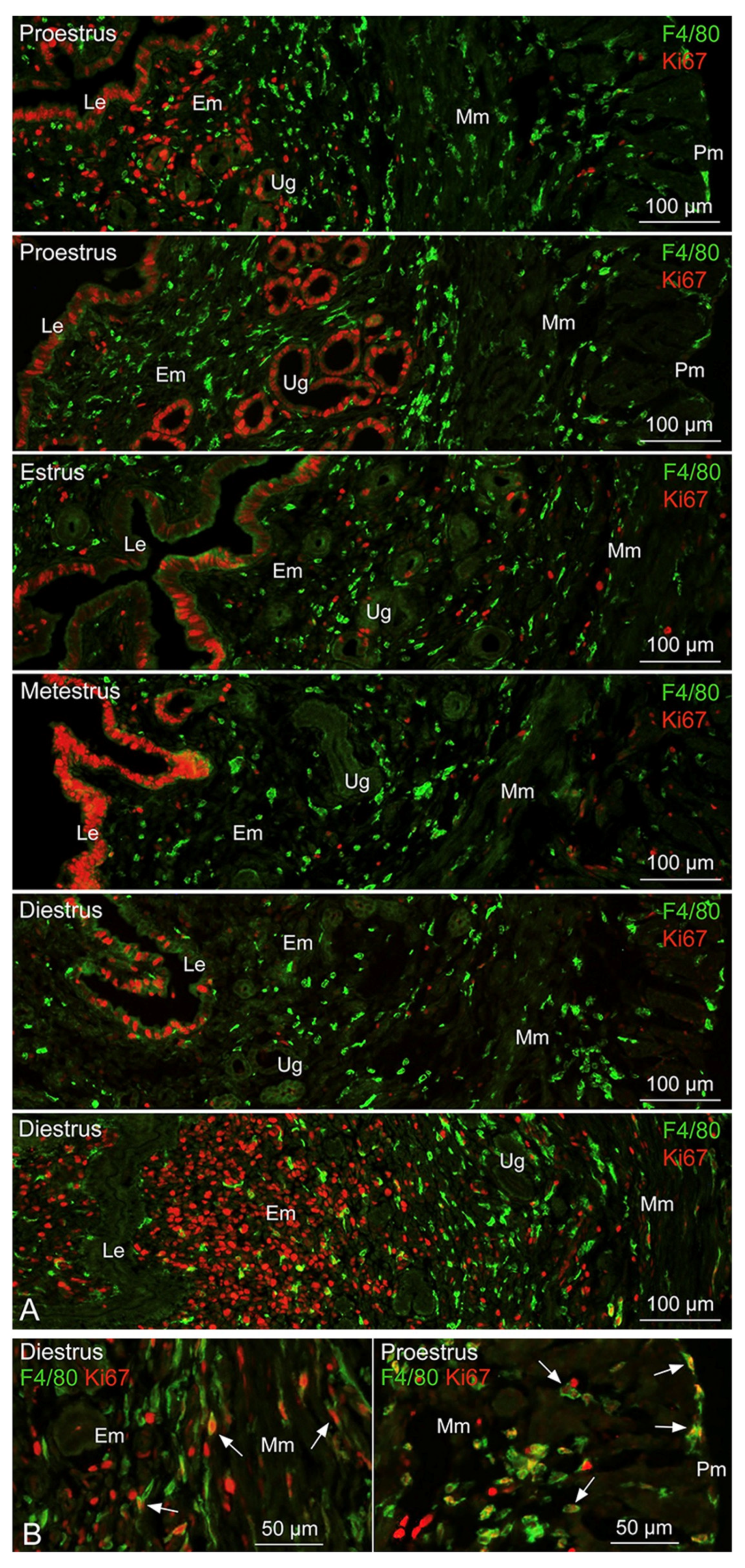
3.1. Distribution and Proliferative Property of Tissue-Resident Mø in the Uterus
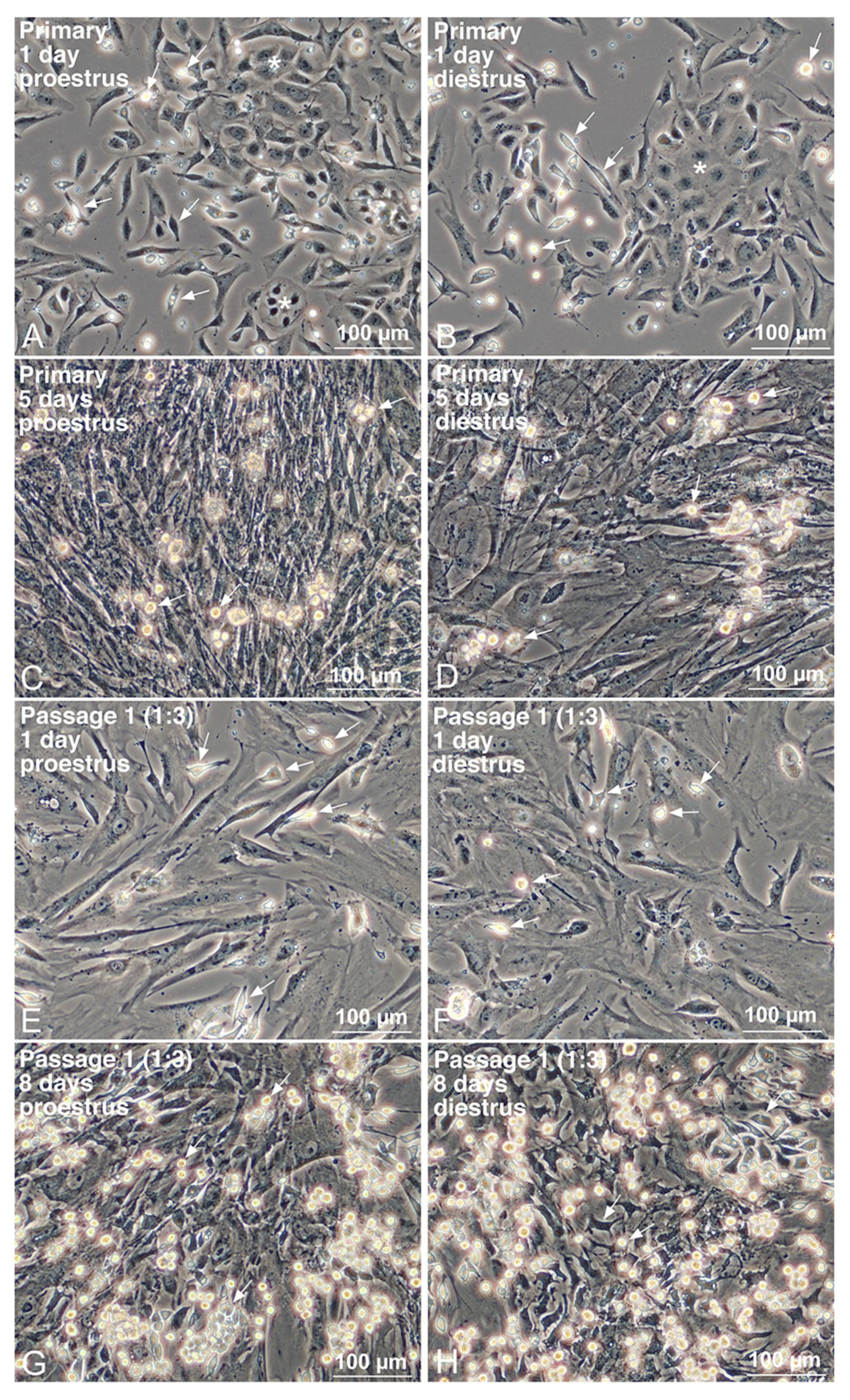
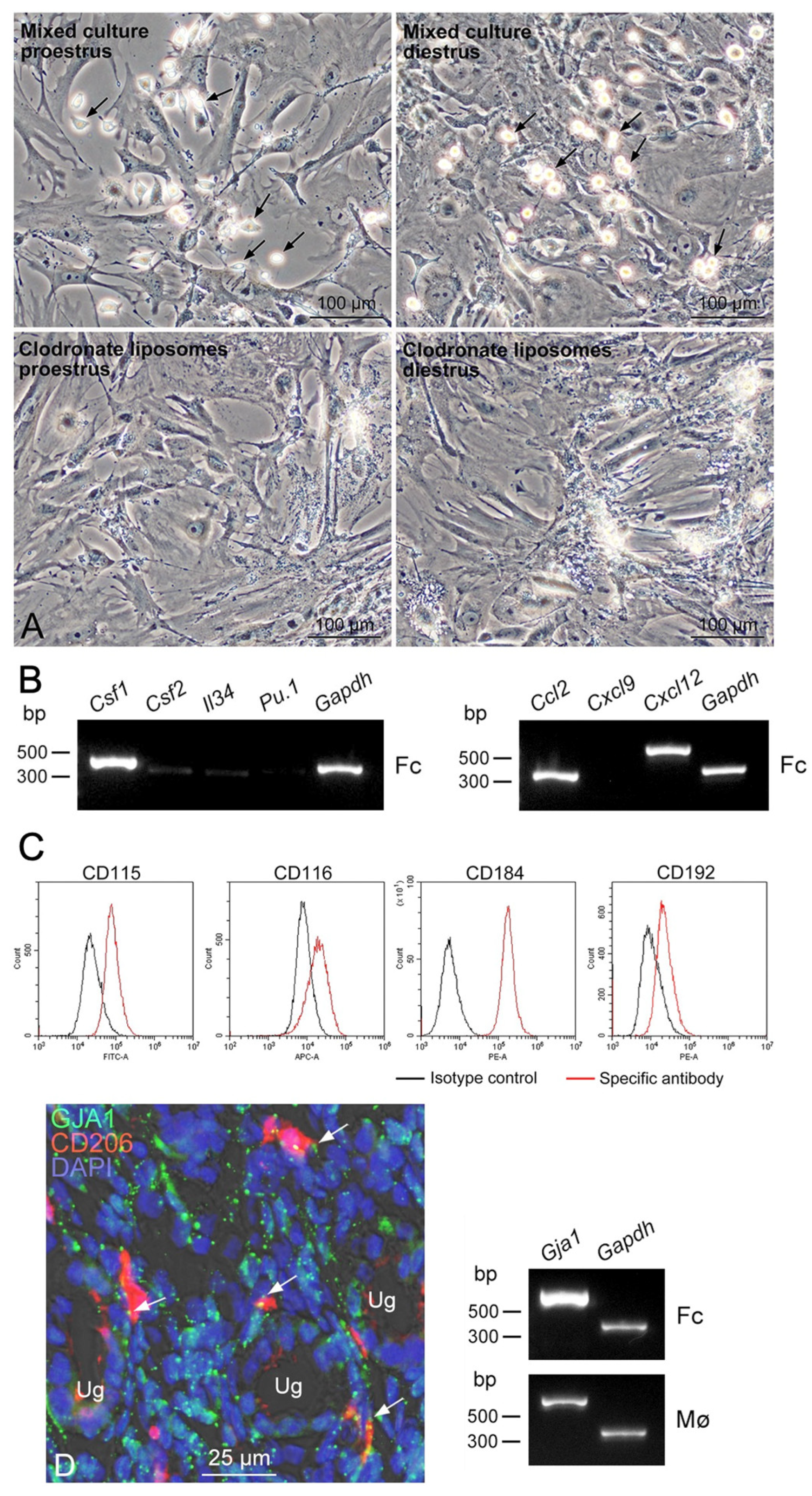
3.2. Propagation of Uterine Mø by Mixed Culture and Their Segregation
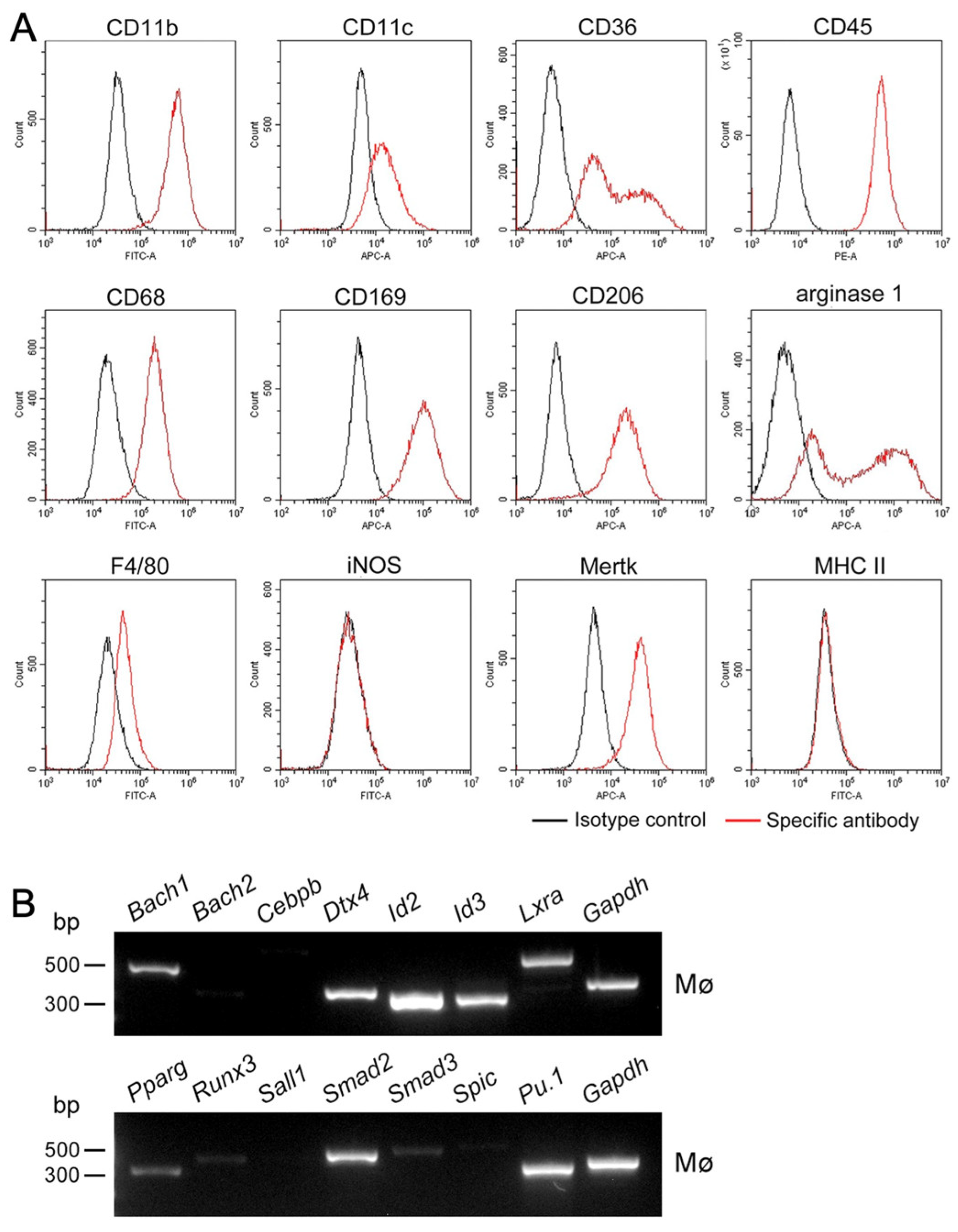
3.3. Expression Profiles of Mø Markers and Those of Transcription Factors Shaping Mø in Propagated Uterine Mø
3.4. Properties of Uterine Fibroblastic Cells as the Niche of Residence in Uterine Mø
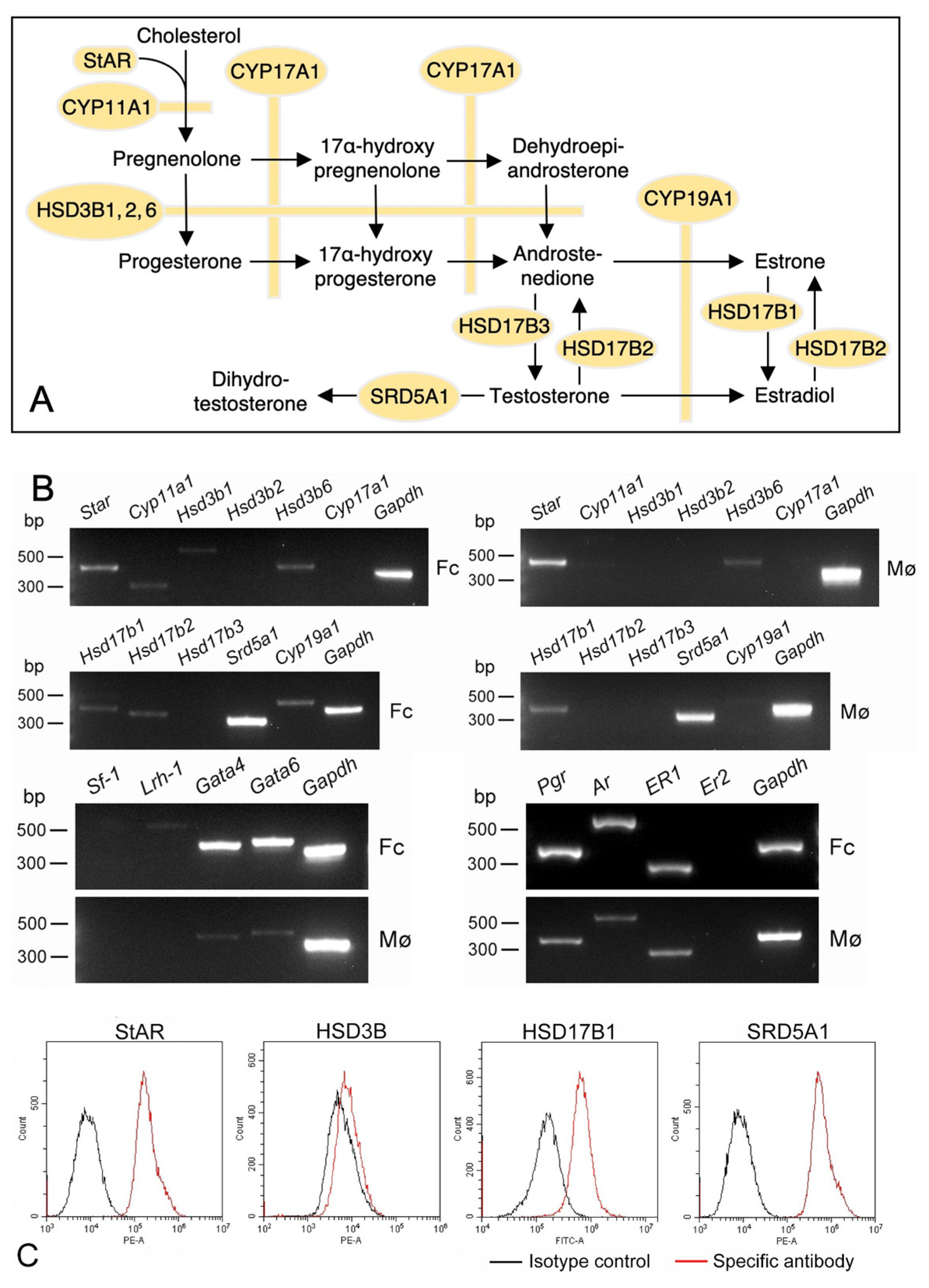
3.5. Expression of Sex-Steroid-Related Molecules in Propagated Uterine Mø and Fibroblastic Cells
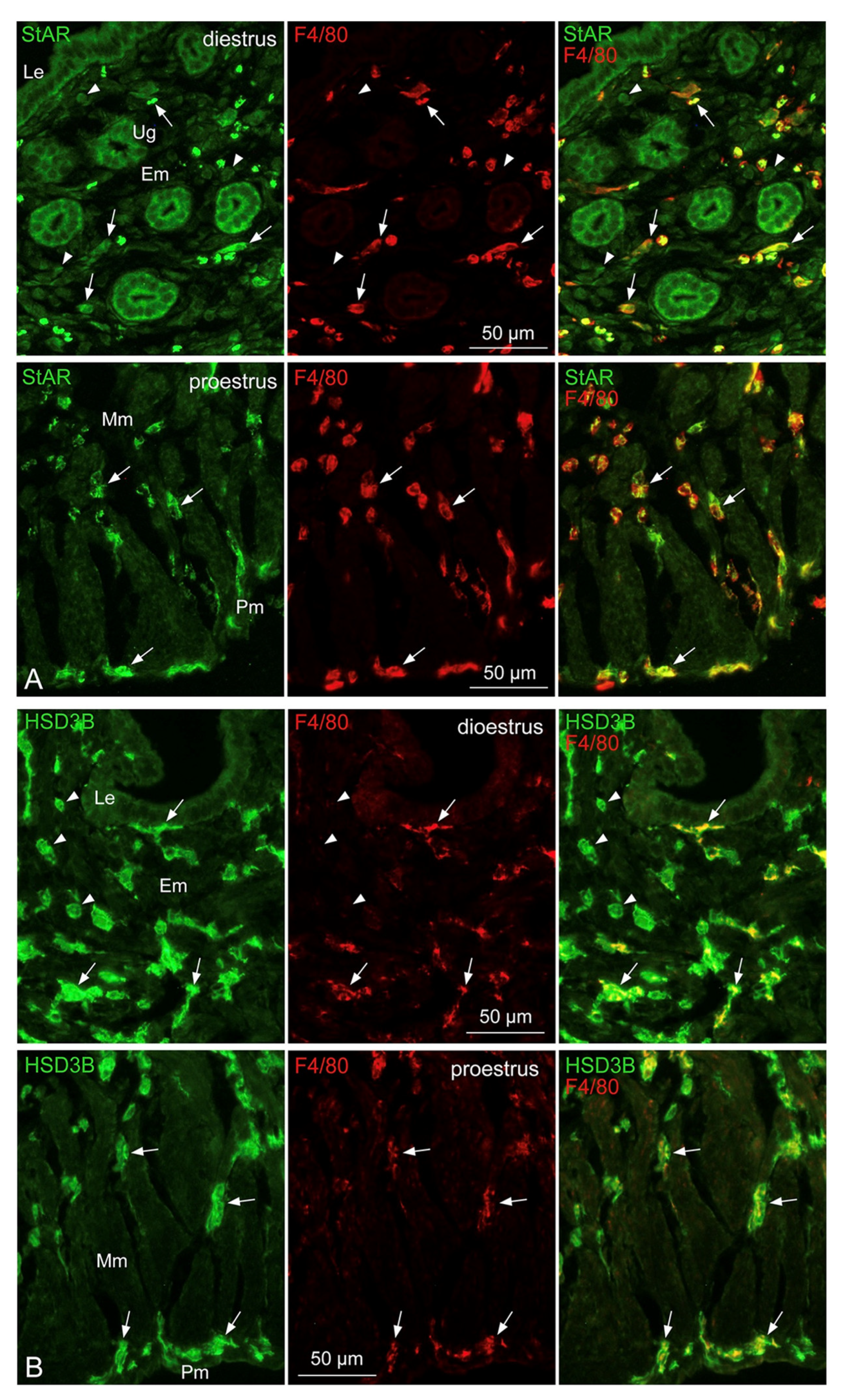
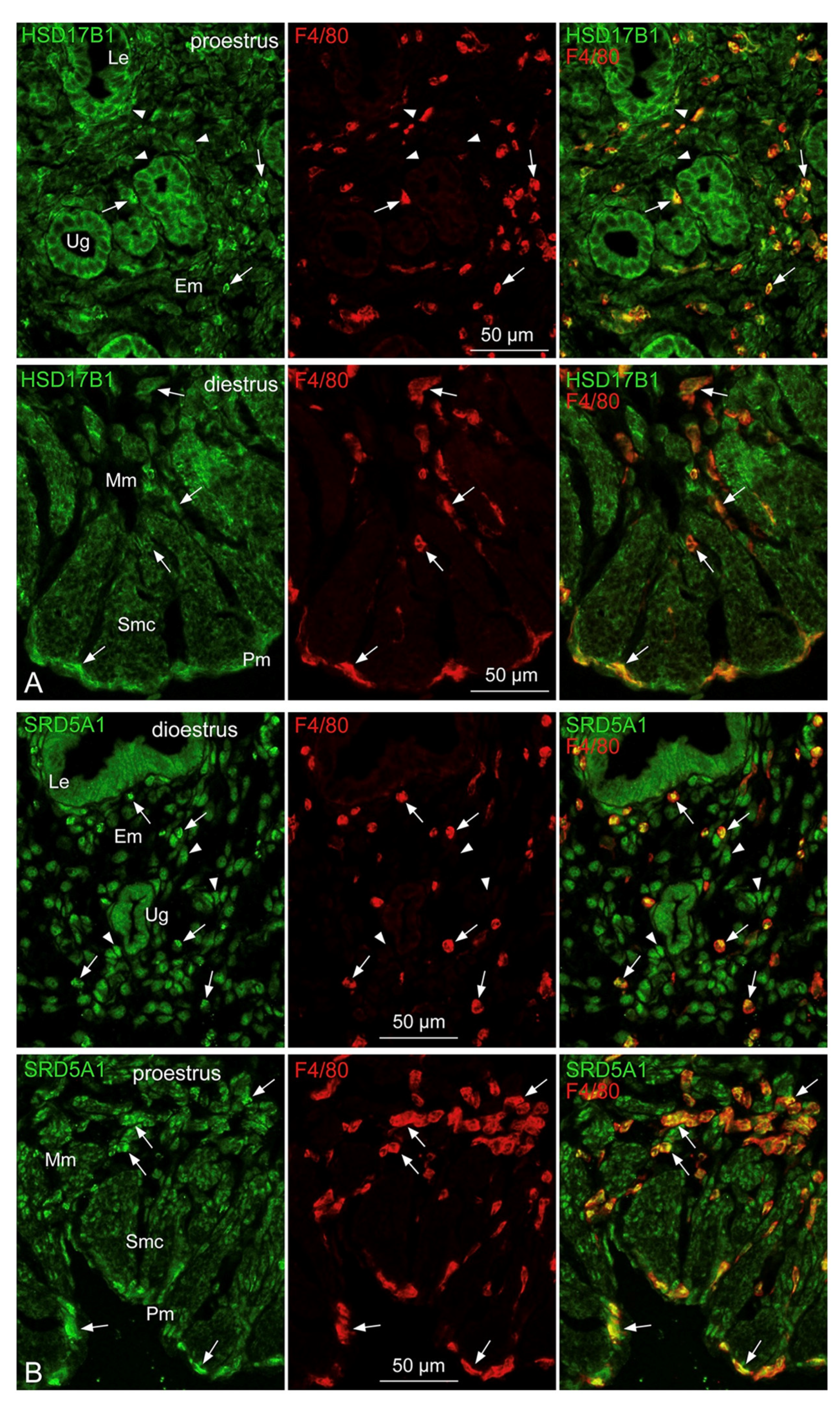
3.6. Localization of StAR, HSD3B, HSD17B1, and SRD5A1 in Tissue-Resident Mø of the Uterus
4. Discussion
4.1. Propagation of Uterine Mø in Mixed Culture
4.2. Metabolic Properties of Sex Steroids in Uterine Mø
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gentek, R.; Molawi, K.; Sieweke, M.H. Tissue macrophage identity and self-renewal. Immunol. Rev. 2014, 262, 56–73. [Google Scholar] [CrossRef]
- Blériot, C.; Chakarov, S.; Ginhoux, F. Determinants of Resident Tissue Macrophage Identity and Function. Immunity 2020, 52, 957–970. [Google Scholar] [CrossRef] [PubMed]
- T’Jonck, W.; Guilliams, M.; Bonnardel, J. Niche signals and transcription factors involved in tissue-resident macrophage development. Cell. Immunol. 2018, 330, 43–53. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, B.; Mosser, D.M.; Zhang, X. TLRs, macrophages, and NK cells: Our understandings of their functions in uterus and ovary. Int. Immunopharmacol. 2011, 11, 1442–1450. [Google Scholar] [CrossRef]
- Jena, M.K.; Nayak, N.; Chen, K.; Nayak, N.R. Role of Macrophages in Pregnancy and Related Complications. Arch. Immunol. Ther. Exp. 2019, 67, 295–309. [Google Scholar] [CrossRef] [PubMed]
- DeLoia, J.A.; Stewart-Akers, A.M.; Brekosky, J.; Kubik, C.J. Effects of exogenous estrogen on uterine leukocyte recruitment. Fertil. Steril. 2002, 77, 548–554. [Google Scholar] [CrossRef] [PubMed]
- De, M.; Wood, G.W. Influence of oestrogen and progesterone on macrophage distribution in the mouse uterus. J. Endocrinol. 1990, 126, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Keenihan, S.N.; Robertson, S.A. Diversity in phenotype and steroid hormone dependence in dendritic cells and macrophages in the mouse uterus. Biol. Reprod. 2004, 70, 1562–1572. [Google Scholar] [CrossRef] [Green Version]
- Cousins, F.L.; Kirkwood, P.M.; Saunders, P.T.; Gibson, D.A. Evidence for a dynamic role for mononuclear phagocytes during endometrial repair and remodelling. Sci. Rep. 2016, 6, 36748. [Google Scholar] [CrossRef] [Green Version]
- Thiruchelvam, U.; Dransfield, I.; Saunders, P.T.; Critchley, H.O. The importance of the macrophage within the human endometrium. J. Leukoc. Biol. 2013, 93, 217–225. [Google Scholar] [CrossRef]
- Sun, F.; Wang, S.; Du, M. Functional regulation of decidual macrophages during pregnancy. J. Reprod. Immunol. 2021, 143, 103264. [Google Scholar] [CrossRef] [PubMed]
- Vallvé-Juanico, J.; Houshdaran, S.; Giudice, L.C. The endometrial immune environment of women with endometriosis. Hum. Reprod. Update 2019, 25, 564–591. [Google Scholar] [CrossRef]
- Bacci, M.; Capobianco, A.; Monno, A.; Cottone, L.; Di Puppo, F.; Camisa, B.; Mariani, M.; Brignole, C.; Ponzoni, M.; Ferrari, S.; et al. Macrophages are alternatively activated in patients with endometriosis and required for growth and vascularization of lesions in a mouse model of disease. Am. J. Pathol. 2009, 175, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Itoh, F.; Komohara, Y.; Takaishi, K.; Honda, R.; Tashiro, H.; Kyo, S.; Katabuchi, H.; Takeya, M. Possible involvement of signal transducer and activator of transcription-3 in cell-cell interactions of peritoneal macrophages and endometrial stromal cells in human endometriosis. Fertil. Steril. 2013, 99, 1705–1713. [Google Scholar] [CrossRef] [PubMed]
- Yellon, S.M.; Greaves, E.; Heuerman, A.C.; Dobyns, A.E.; Norman, J.E. Effects of macrophage depletion on characteristics of cervix remodeling and pregnancy in CD11b-dtr mice. Biol. Reprod. 2019, 100, 1386–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Care, A.S.; Diener, K.R.; Jasper, M.J.; Brown, H.M.; Ingman, W.V.; Robertson, S.A. Macrophages regulate corpus luteum development during embryo implantation in mice. J. Clin. Investig. 2013, 123, 3472–3487. [Google Scholar] [CrossRef] [Green Version]
- Wood, G.W.; Hausmann, E.; Choudhuri, R. Relative role of CSF-1, MCP-1/JE, and RANTES in macrophage recruitment during successful pregnancy. Mol. Reprod. Dev. 1997, 46, 62–69; discussion 69–70. [Google Scholar] [CrossRef]
- Shimada-Hiratsuka, M.; Naito, M.; Kaizu, C.; Shuying, J.; Hasegawa, G.; Shultz, L.D. Defective macrophage recruitment and clearance of apoptotic cells in the uterus of osteopetrotic mutant mice lacking macrophage colony-stimulating factor (M-CSF). J. Submicrosc. Cytol. Pathol. 2000, 32, 297–307. [Google Scholar]
- Hume, D.A. The mononuclear phagocyte system. Curr. Opin. Immunol. 2006, 18, 49–53. [Google Scholar] [CrossRef]
- Dziennis, S.; Van Etten, R.A.; Pahl, H.L.; Morris, D.L.; Rothstein, T.L.; Blosch, C.M.; Perlmutter, R.M.; Tenen, D.G. The CD11b promoter directs high-level expression of reporter genes in macrophages in transgenic mice. Blood 1995, 85, 319–329. [Google Scholar] [CrossRef]
- Ogawa, K.; Tsurutani, M.; Hashimoto, A.; Soeda, M. Simple propagation method for resident macrophages by co-culture and subculture, and their isolation from various organs. BMC Immunol. 2019, 20, 34. [Google Scholar] [CrossRef] [PubMed]
- Tsurutani, M.; Horie, H.; Ogawa, K. Cell Properties of Lung Tissue-Resident Macrophages Propagated by Co-Culture with Lung Fibroblastic Cells from C57BL/6 and BALB/c Mice. Biomedicines 2021, 9, 1241. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, S.; Yamamoto, K.; Ogawa, K. Testicular Macrophages Produce Progesterone De Novo Promoted by cAMP and Inhibited by M1 Polarization Inducers. Biomedicines 2022, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Kohara, S.; Ogawa, K. Eph/Ephrin Promotes the Adhesion of Liver Tissue-Resident Macrophages to a Mimicked Surface of Liver Sinusoidal Endothelial Cells. Biomedicines 2022, 10, 3234. [Google Scholar] [CrossRef] [PubMed]
- Tagliani, E.; Shi, C.; Nancy, P.; Tay, C.S.; Pamer, E.G.; Erlebacher, A. Coordinate regulation of tissue macrophage and dendritic cell population dynamics by CSF-1. J. Exp. Med. 2011, 208, 1901–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, P.E.; Nishimura, K.; Zhu, L.; Pollard, J.W. Macrophages: Important accessory cells for reproductive function. J. Leukoc. Biol. 1999, 66, 765–772. [Google Scholar] [CrossRef]
- Hatayama, H.; Kanzaki, H.; Iwai, M.; Kariya, M.; Fujimoto, M.; Higuchi, T.; Kojima, K.; Nakayama, H.; Mori, T.; Fujita, J. Progesterone enhances macrophage colony-stimulating factor production in human endometrial stromal cells in vitro. Endocrinology 1994, 135, 1921–1927. [Google Scholar] [CrossRef]
- Simitsidellis, I.; Saunders, P.T.K.; Gibson, D.A. Androgens and endometrium: New insights and new targets. Mol. Cell. Endocrinol. 2018, 465, 48–60. [Google Scholar] [CrossRef]
- Ajayi, A.F.; Akhigbe, R.E. Staging of the estrous cycle and induction of estrus in experimental rodents: An update. Fertil. Res. Pract. 2020, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.A.; Simitsidellis, I.; Collins, F.; Saunders, P.T.K. Androgens, oestrogens and endometrium: A fine balance between perfection and pathology. J. Endocrinol. 2020, 246, R75–R93. [Google Scholar] [CrossRef]
- Gibson, D.A.; Simitsidellis, I.; Saunders, P.T. Regulation of androgen action during establishment of pregnancy. J. Mol. Endocrinol. 2016, 57, R35–R47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinsohn, M.C.; Smith, O.E.; Bertolin, K.; Murphy, B.D. The Orphan Nuclear Receptors Steroidogenic Factor-1 and Liver Receptor Homolog-1: Structure, Regulation, and Essential Roles in Mammalian Reproduction. Physiol. Rev. 2019, 99, 1249–1279. [Google Scholar] [CrossRef]
- Tremblay, J.J.; Viger, R.S. Novel roles for GATA transcription factors in the regulation of steroidogenesis. J. Steroid Biochem. Mol. Biol. 2003, 85, 291–298. [Google Scholar] [CrossRef]
- Mei, J.; Zhou, W.J.; Li, S.Y.; Li, M.Q.; Sun, H.X. Interleukin-22 secreted by ectopic endometrial stromal cells and natural killer cells promotes the recruitment of macrophages through promoting CCL2 secretion. Am. J. Reprod. Immunol. 2019, 82, e13166. [Google Scholar] [CrossRef]
- Chao, H.H.; Li, L.; Gao, X.; Wang, C.; Yue, W. CXCL12 expression in aborted mouse uteri induced by IFN-γ: Potential anti-inflammatory effect involves in endometrial restoration after abortion in mice. Gene 2019, 700, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Venkata Suresh, V.; Yahata, Y.; Nakano, M.; Suzuki, S.; Suzuki, S.; Yamada, S.; Kitaura, H.; Mizoguchi, I.; Noiri, Y.; et al. Inhibition of the CXCL9-CXCR3 axis suppresses the progression of experimental apical periodontitis by blocking macrophage migration and activation. Sci. Rep. 2021, 11, 2613. [Google Scholar] [CrossRef]
- Yu, J.; Berga, S.L.; Zou, W.; Yook, D.G.; Pan, J.C.; Andrade, A.A.; Zhao, L.; Sidell, N.; Bagchi, I.C.; Bagchi, M.K.; et al. IL-1β Inhibits Connexin 43 and Disrupts Decidualization of Human Endometrial Stromal Cells Through ERK1/2 and p38 MAP Kinase. Endocrinology 2017, 158, 4270–4285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yona, S.; Kim, K.W.; Wolf, Y.; Mildner, A.; Varol, D.; Breker, M.; Strauss-Ayali, D.; Viukov, S.; Guilliams, M.; Misharin, A.; et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity 2013, 38, 79–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; He, B.; Xu, X.; Wang, J. Comprehensive analysis of leukocytes, vascularization and matrix metalloproteinases in human menstrual xenograft model. PLoS ONE 2011, 6, e16840. [Google Scholar] [CrossRef] [Green Version]
- Mills, C.D. Anatomy of a discovery: m1 and m2 macrophages. Front. Immunol. 2015, 6, 212. [Google Scholar] [CrossRef]
- Buechler, C.; Ritter, M.; Orsó, E.; Langmann, T.; Klucken, J.; Schmitz, G. Regulation of scavenger receptor CD163 expression in human monocytes and macrophages by pro- and antiinflammatory stimuli. J. Leukoc. Biol. 2000, 67, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Quillay, H.; El Costa, H.; Marlin, R.; Duriez, M.; Cannou, C.; Chrétien, F.; Fernandez, H.; Lebreton, A.; Ighil, J.; Schwartz, O.; et al. Distinct characteristics of endometrial and decidual macrophages and regulation of their permissivity to HIV-1 infection by SAMHD1. J. Virol. 2015, 89, 1329–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cominelli, A.; Gaide Chevronnay, H.P.; Lemoine, P.; Courtoy, P.J.; Marbaix, E.; Henriet, P. Matrix metalloproteinase-27 is expressed in CD163+/CD206+ M2 macrophages in the cycling human endometrium and in superficial endometriotic lesions. Mol. Hum. Reprod. 2014, 20, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.L.; Collins, J.; Shipman, E.P.; Wira, C.R.; Guyre, P.M.; Pioli, P.A. A subset of human uterine endometrial macrophages is alternatively activated. Am. J. Reprod. Immunol. 2012, 68, 374–386. [Google Scholar] [CrossRef] [Green Version]
- Sieweke, M.H.; Allen, J.E. Beyond stem cells: Self-renewal of differentiated macrophages. Science 2013, 342, 1242974. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Abram, C.L.; Lowell, C.A. The ins and outs of leukocyte integrin signaling. Annu. Rev. Immunol. 2009, 27, 339–362. [Google Scholar] [CrossRef] [Green Version]
- Kinashi, T. Intracellular signalling controlling integrin activation in lymphocytes. Nat. Rev. Immunol. 2005, 5, 546–559. [Google Scholar] [CrossRef]
- Critchley, H.O.D.; Maybin, J.A.; Armstrong, G.M.; Williams, A.R.W. Physiology of the Endometrium and Regulation of Menstruation. Physiol. Rev. 2020, 100, 1149–1179. [Google Scholar] [CrossRef]
- Mote, P.A.; Arnett-Mansfield, R.L.; Gava, N.; deFazio, A.; Mulac-Jericevic, B.; Conneely, O.M.; Clarke, C.L. Overlapping and distinct expression of progesterone receptors A and B in mouse uterus and mammary gland during the estrous cycle. Endocrinology 2006, 147, 5503–5512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitaker, L.H.; Murray, A.A.; Matthews, R.; Shaw, G.; Williams, A.R.; Saunders, P.T.; Critchley, H.O. Selective progesterone receptor modulator (SPRM) ulipristal acetate (UPA) and its effects on the human endometrium. Hum. Reprod. 2017, 32, 531–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.N.; Masuzaki, H.; Fujishita, A.; Kitajima, M.; Sekine, I.; Matsuyama, T.; Ishimaru, T. Estrogen and progesterone receptor expression in macrophages and regulation of hepatocyte growth factor by ovarian steroids in women with endometriosis. Hum. Reprod. 2005, 20, 2004–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.A.; Bulmer, J.N.; Murdoch, A.P. Endometrial leucocytes: Expression of steroid hormone receptors. J. Clin. Pathol. 1998, 51, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Rubinow, K.B. An intracrine view of sex steroids, immunity, and metabolic regulation. Mol. Metab. 2018, 15, 92–103. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Melnichenko, A.A.; Myasoedova, V.A.; Grechko, A.V.; Orekhov, A.N. Mechanisms of foam cell formation in atherosclerosis. J. Mol. Med. 2017, 95, 1153–1165. [Google Scholar] [CrossRef]
- Sobolev, V.V.; Tchepourina, E.; Korsunskaya, I.M.; Geppe, N.A.; Chebysheva, S.N.; Soboleva, A.G.; Mezentsev, A. The Role of Transcription Factor PPAR-γ in the Pathogenesis of Psoriasis, Skin Cells, and Immune Cells. Int. J. Mol. Sci. 2022, 23, 9708. [Google Scholar] [CrossRef]
- Sodhi, R.K.; Singh, N. Liver X receptors: Emerging therapeutic targets for Alzheimer’s disease. Pharmacol. Res. 2013, 72, 45–51. [Google Scholar] [CrossRef]
- Anuka, E.; Gal, M.; Stocco, D.M.; Orly, J. Expression and roles of steroidogenic acute regulatory (StAR) protein in ‘non-classical’, extra-adrenal and extra-gonadal cells and tissues. Mol. Cell. Endocrinol. 2013, 371, 47–61. [Google Scholar] [CrossRef]
- Nielsen, M.S.; Axelsen, L.N.; Sorgen, P.L.; Verma, V.; Delmar, M.; Holstein-Rathlou, N.H. Gap junctions. Compr. Physiol. 2012, 2, 1981–2035. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Lorena, P.D.; Ng, L.K.; Lim, D.; Shen, L.; Siow, W.Y.; Teh, M.; Reichardt, J.K.; Salto-Tellez, M. Differential expression of steroid 5alpha-reductase isozymes and association with disease severity and angiogenic genes predict their biological role in prostate cancer. Endocr. Relat. Cancer 2010, 17, 757–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swerdloff, R.S.; Dudley, R.E.; Page, S.T.; Wang, C.; Salameh, W.A. Dihydrotestosterone: Biochemistry, Physiology, and Clinical Implications of Elevated Blood Levels. Endocr. Rev. 2017, 38, 220–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogawa, K.; Tanida, T. Mixed-Culture Propagation of Uterine-Tissue-Resident Macrophages and Their Expression Properties of Steroidogenic Molecules. Biomedicines 2023, 11, 985. https://doi.org/10.3390/biomedicines11030985
Ogawa K, Tanida T. Mixed-Culture Propagation of Uterine-Tissue-Resident Macrophages and Their Expression Properties of Steroidogenic Molecules. Biomedicines. 2023; 11(3):985. https://doi.org/10.3390/biomedicines11030985
Chicago/Turabian StyleOgawa, Kazushige, and Takashi Tanida. 2023. "Mixed-Culture Propagation of Uterine-Tissue-Resident Macrophages and Their Expression Properties of Steroidogenic Molecules" Biomedicines 11, no. 3: 985. https://doi.org/10.3390/biomedicines11030985