
Telomere Transcription in MLL-Rearranged Leukemia Cell Lines: Increased Levels of TERRA Associate with Lymphoid Lineage and Are Independent of Telomere Length and Ploidy

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. ChIP Analysis
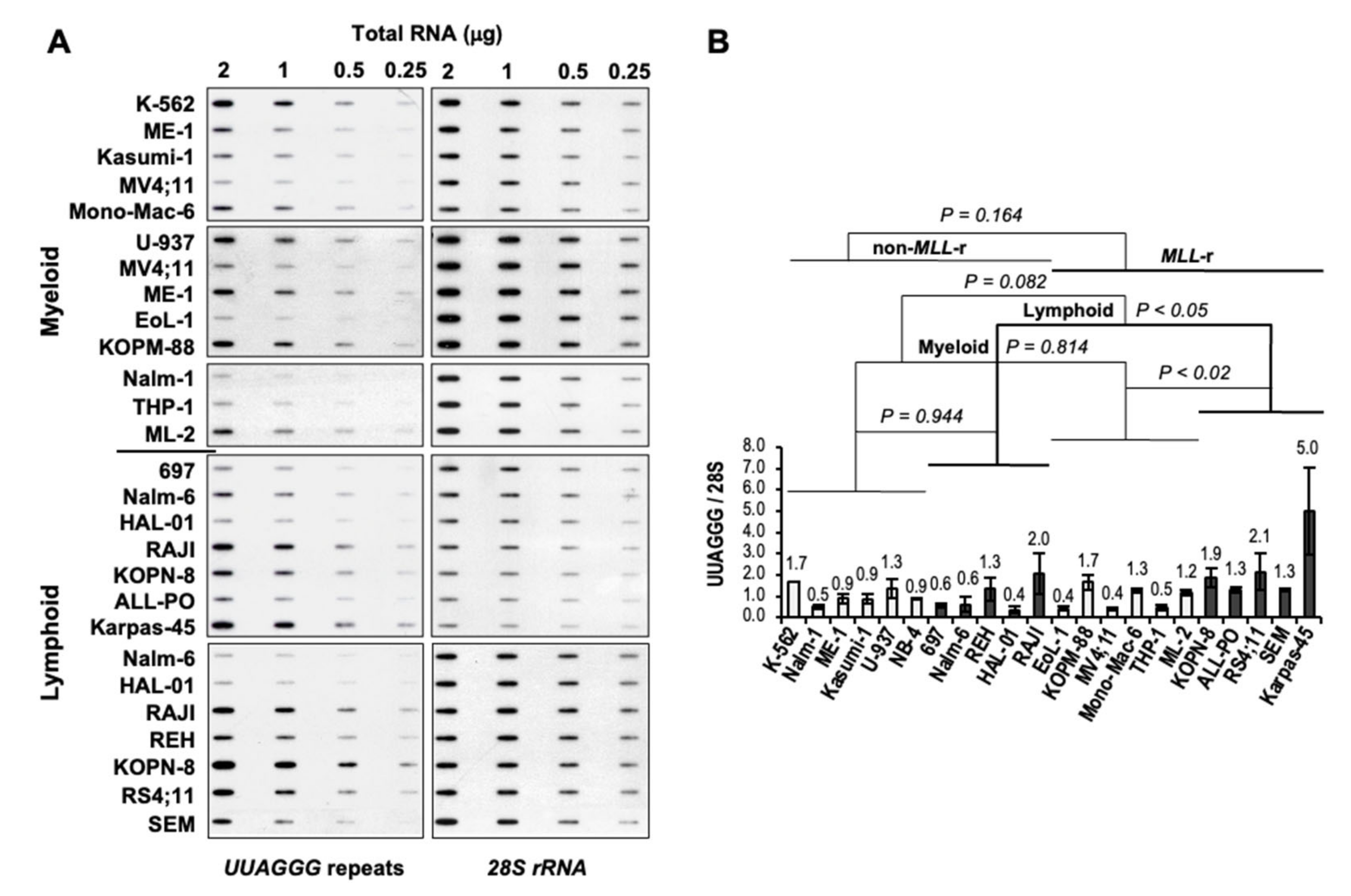
2.3. RNA Slot-Blot Analysis
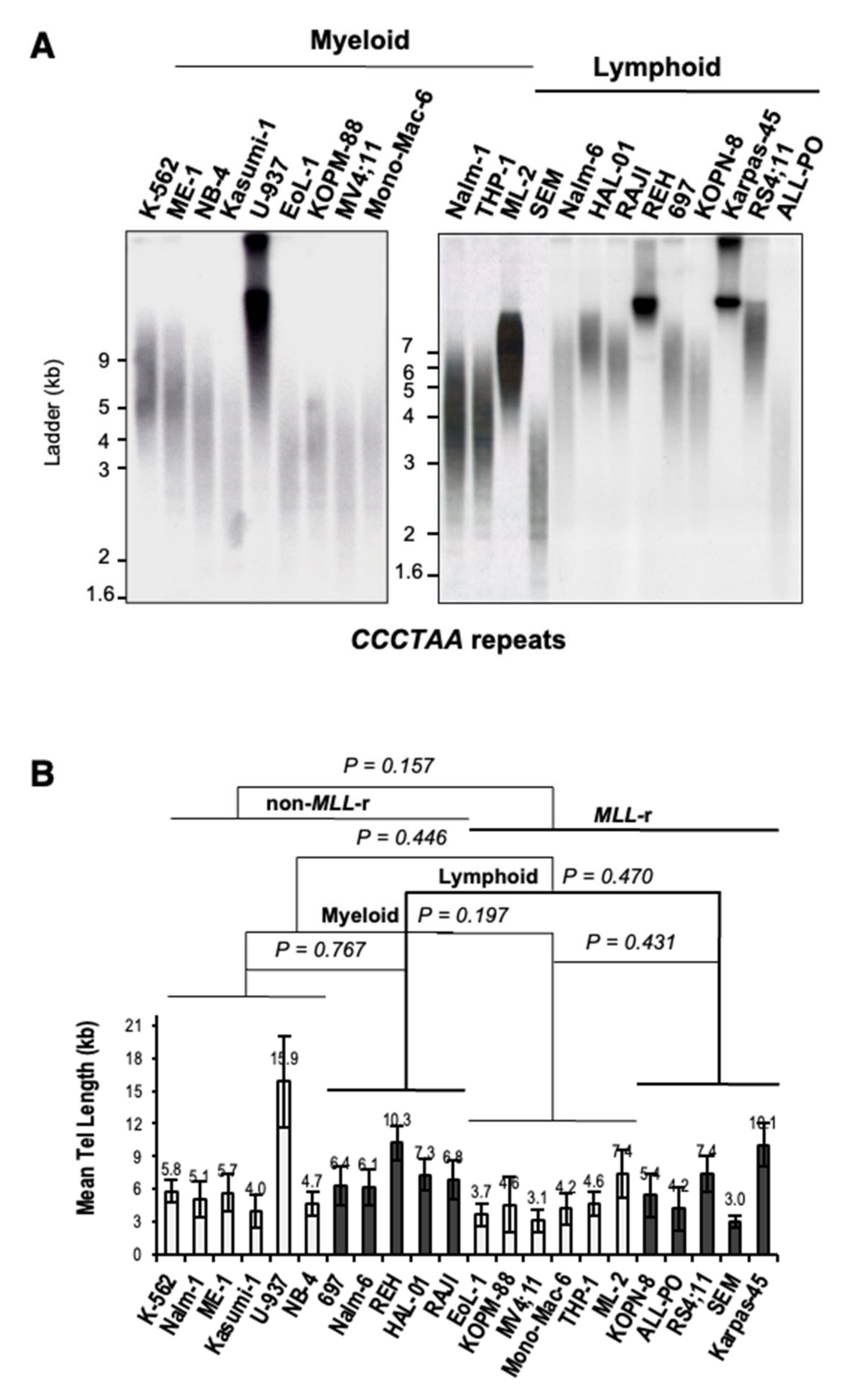
2.4. Terminal Restriction Fragment (TRF) Analysis
2.5. Telomeric-Repeat Amplification Protocol (TRAP)
3. Results
3.1. Telomeric Chromatin Binding of MLL Proteins and Telomerase Activity in MLL-Rearranged Acute Lymphoblastic Leukemia
3.2. Increased Levels of UUAGGG Repeat-Containing RNA in MLL-r ALL
3.3. MLL Rearrangements Do Not Affect Telomere Length in Leukemia
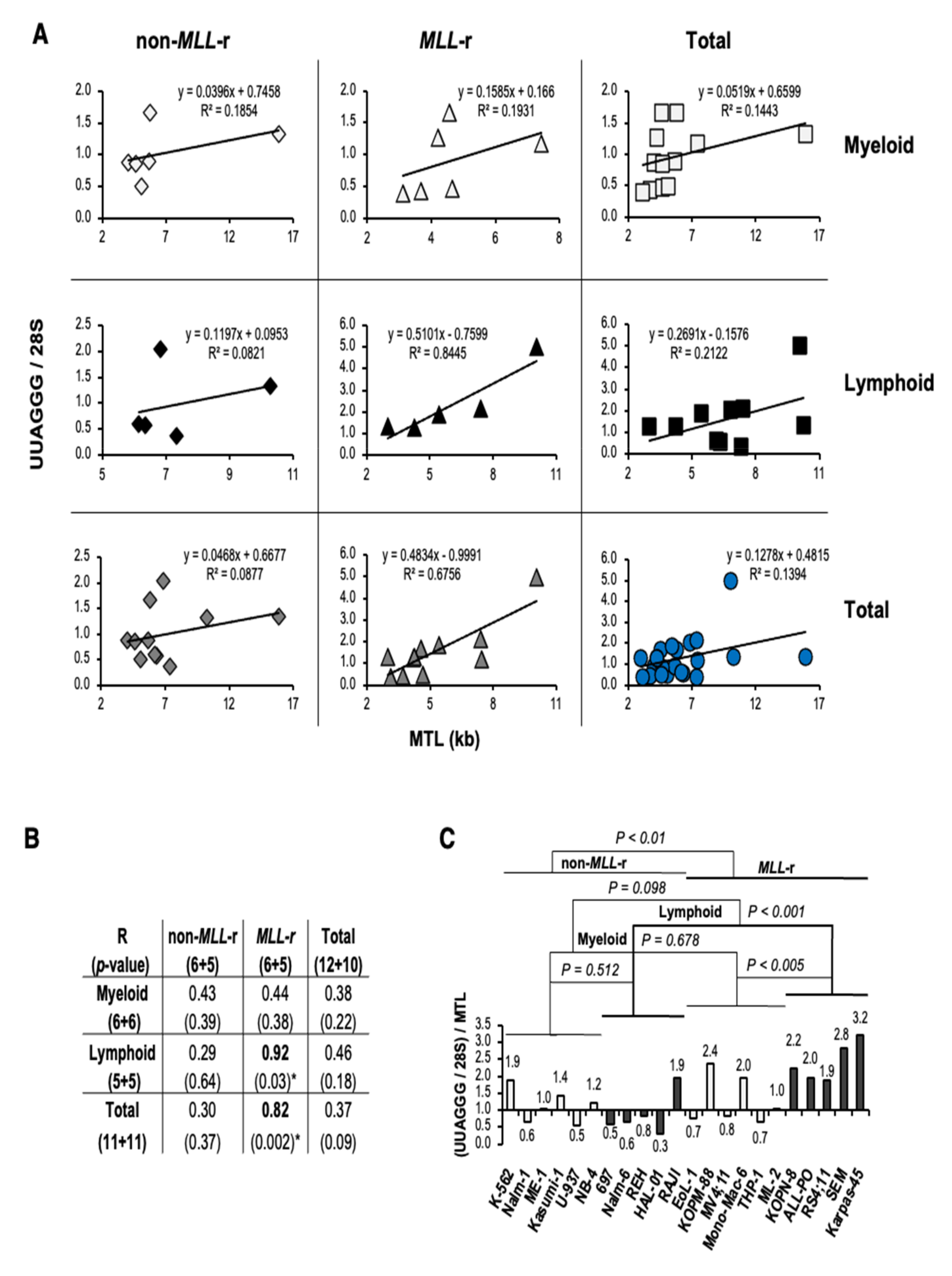
3.4. UUAGGG Repeat-Containing RNA Levels in MLL-r ALL Correlate with Telomere Length and Reflect Increased Levels of TERRA
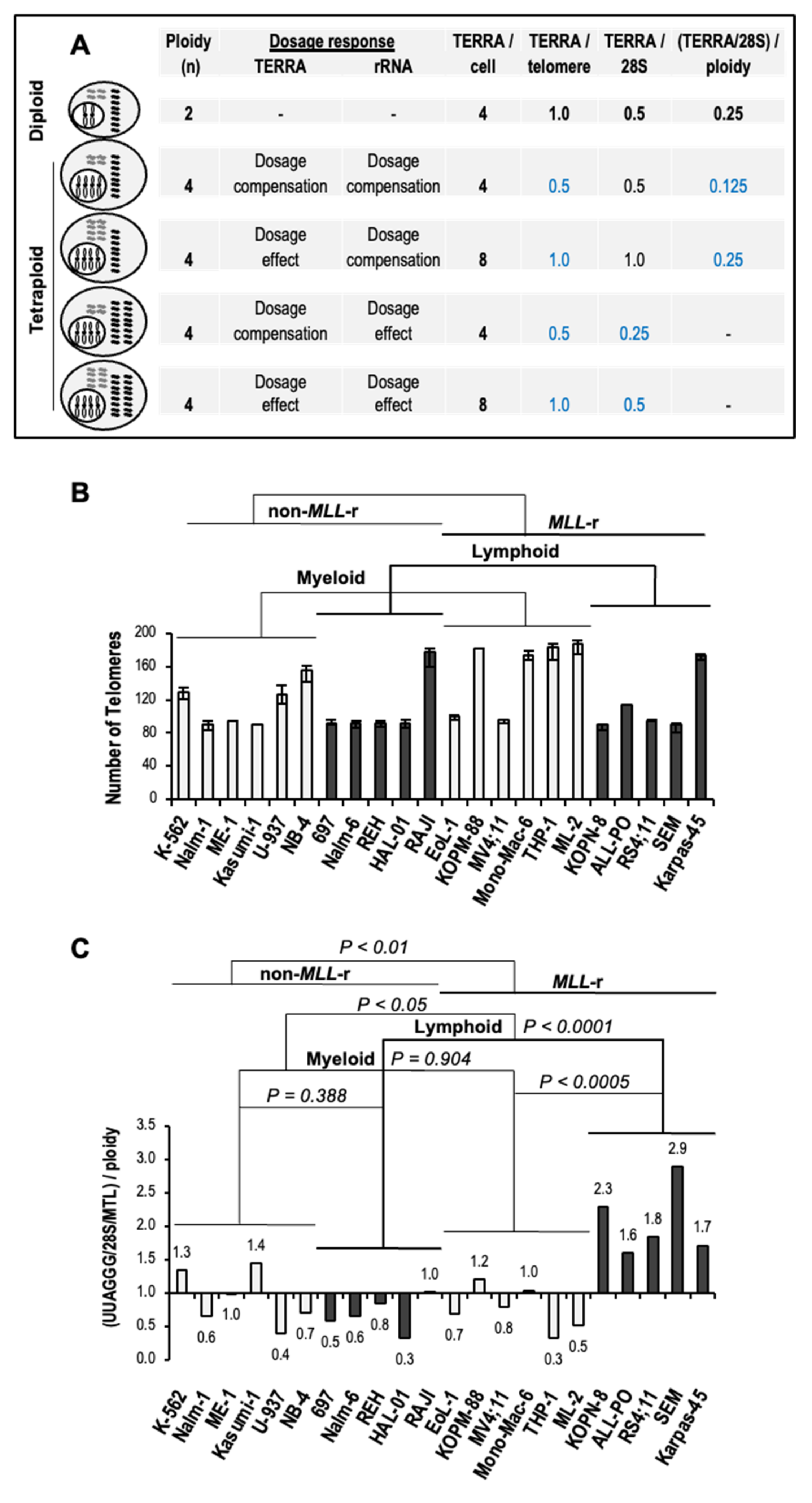
3.5. Increased Levels of TERRA in MLL-r ALL Are Independent of Ploidy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric Repeat Containing RNA and RNA Surveillance Factors at Mammalian Chromosome Ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Schoeftner, S.; Blasco, M.A. Developmentally Regulated Transcription of Mammalian Telomeres by DNA-Dependent RNA Polymerase II. Nat. Cell Biol. 2008, 10, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Farnung, B.O.; Brun, C.M.; Arora, R.; Lorenzi, L.E.; Azzalin, C.M. Telomerase Efficiently Elongates Highly Transcribing Telomeres in Human Cancer Cells. PLoS ONE 2012, 7, e35714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redon, S.; Zemp, I.; Lingner, J. A Three-State Model for the Regulation of Telomerase by TERRA and HnRNPA1. Nucleic Acids Res. 2013, 41, 9117–9128. [Google Scholar] [CrossRef] [Green Version]
- Cusanelli, E.; Romero, C.A.P.; Chartrand, P. Telomeric Noncoding RNA TERRA Is Induced by Telomere Shortening to Nucleate Telomerase Molecules at Short Telomeres. Mol. Cell 2013, 51, 780–791. [Google Scholar] [CrossRef] [Green Version]
- Balk, B.; Maicher, A.; Dees, M.; Klermund, J.; Luke-Glaser, S.; Bender, K.; Luke, B. Telomeric RNA-DNA Hybrids Affect Telomere-Length Dynamics and Senescence. Nat. Struct. Mol. Biol. 2013, 20, 1199–1205. [Google Scholar] [CrossRef]
- Yu, T.-Y.; Kao, Y.; Lin, J.-J. Telomeric Transcripts Stimulate Telomere Recombination to Suppress Senescence in Cells Lacking Telomerase. Proc. Natl. Acad. Sci. USA 2014, 111, 3377–3382. [Google Scholar] [CrossRef] [Green Version]
- Montero, J.J.; López de Silanes, I.; Graña, O.; Blasco, M.A. Telomeric RNAs Are Essential to Maintain Telomeres. Nat. Commun. 2016, 7, 12534. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.P.; Froberg, J.E.; Kesner, B.; Oh, H.J.; Ji, F.; Sadreyev, R.; Pinter, S.F.; Lee, J.T. PAR-TERRA Directs Homologous Sex Chromosome Pairing. Nat. Struct. Mol. Biol. 2017, 24, 620–631. [Google Scholar] [CrossRef] [Green Version]
- De Silanes, I.L.; Graña, O.; De Bonis, M.L.; Dominguez, O.; Pisano, D.G.; Blasco, M.A. Identification of TERRA Locus Unveils a Telomere Protection Role through Association to Nearly All Chromosomes. Nat. Commun. 2014, 5, 4723. [Google Scholar] [CrossRef] [Green Version]
- Viceconte, N.; Loriot, A.; Lona Abreu, P.; Scheibe, M.; Fradera Sola, A.; Butter, F.; De Smet, C.; Azzalin, C.M.; Arnoult, N.; Decottignies, A. PAR-TERRA Is the Main Contributor to Telomeric Repeat-Containing RNA Transcripts in Normal and Cancer Mouse Cells. RNA 2021, 27, 106–121. [Google Scholar] [CrossRef]
- Nergadze, S.G.; Farnung, B.O.; Wischnewski, H.; Khoriauli, L.; Vitelli, V.; Chawla, R.; Giulotto, E.; Azzalin, C.M. CpG-Island Promoters Drive Transcription of Human Telomeres. RNA 2009, 15, 2186–2194. [Google Scholar] [CrossRef] [Green Version]
- Feretzaki, M.; Nunes, P.R.; Lingner, J. Expression and Differential Regulation of Human TERRA at Several Chromosome Ends. RNA 2019, 25, 1470–1480. [Google Scholar] [CrossRef]
- Chu, H.-P.; Cifuentes-Rojas, C.; Kesner, B.; Aeby, E.; Lee, H.; Wei, C.; Oh, H.J.; Boukhali, M.; Haas, W.; Lee, J.T. TERRA RNA Antagonizes ATRX and Protects Telomeres. Cell 2017, 170, 86–101. [Google Scholar] [CrossRef] [Green Version]
- Coate, J.E.; Doyle, J.J. Variation in Transcriptome Size: Are We Getting the Message? Chromosoma 2015, 124, 27–43. [Google Scholar] [CrossRef]
- Schoenfelder, K.P.; Fox, D.T. The Expanding Implications of Polyploidy. J. Cell Biol. 2015, 209, 485–491. [Google Scholar] [CrossRef]
- De Silanes, I.L.; D’Alcontres, M.S.; Blasco, M.A. TERRA Transcripts Are Bound by a Complex Array of RNA-Binding Proteins. Nat. Commun. 2010, 1, 33. [Google Scholar] [CrossRef] [Green Version]
- Flynn, R.L.; Centore, R.C.; O’Sullivan, R.J.; Rai, R.; Tse, A.; Songyang, Z.; Chang, S.; Karlseder, J.; Zou, L. TERRA and HnRNPA1 Orchestrate an RPA-to-POT1 Switch on Telomeric Single-Stranded DNA. Nature 2011, 471, 532–536. [Google Scholar] [CrossRef] [Green Version]
- Flynn, R.L.; Cox, K.E.; Jeitany, M.; Wakimoto, H.; Bryll, A.R.; Ganem, N.J.; Bersani, F.; Pineda, J.R.; Suvà, M.L.; Benes, C.H.; et al. Alternative Lengthening of Telomeres Renders Cancer Cells Hypersensitive to ATR Inhibitors. Science 2015, 347, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Yoshimura, H.; Shimada, R.; Hattori, M.; Eguchi, M.; Fujiwara, T.K.; Kusumi, A.; Ozawa, T. Spatiotemporal Analysis with a Genetically Encoded Fluorescent RNA Probe Reveals TERRA Function around Telomeres. Sci. Rep. 2016, 6, 38910. [Google Scholar] [CrossRef] [Green Version]
- Arora, R.; Lee, Y.; Wischnewski, H.; Brun, C.M.; Schwarz, T.; Azzalin, C.M. RNaseH1 Regulates TERRA-Telomeric DNA Hybrids and Telomere Maintenance in ALT Tumour Cells. Nat. Commun. 2014, 5, 5220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA Binding to TRF2 Facilitates Heterochromatin Formation and ORC Recruitment at Telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnoult, N.; Van Beneden, A.; Decottignies, A. Telomere Length Regulates TERRA Levels through Increased Trimethylation of Telomeric H3K9 and HP1α. Nat. Struct. Mol. Biol. 2012, 19, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Montero, J.J.; López-Silanes, I.; Megías, D.; F Fraga, M.; Castells-García, Á.; Blasco, M.A. TERRA Recruitment of Polycomb to Telomeres Is Essential for Histone Trymethylation Marks at Telomeric Heterochromatin. Nat. Commun. 2018, 9, 1548. [Google Scholar] [CrossRef] [Green Version]
- Ng, L.J.; Cropley, J.E.; Pickett, H.A.; Reddel, R.R.; Suter, C.M. Telomerase Activity Is Associated with an Increase in DNA Methylation at the Proximal Subtelomere and a Reduction in Telomeric Transcription. Nucleic Acids Res. 2009, 37, 1152–1159. [Google Scholar] [CrossRef]
- Episkopou, H.; Draskovic, I.; Van Beneden, A.; Tilman, G.; Mattiussi, M.; Gobin, M.; Arnoult, N.; Londoño-Vallejo, A.; Decottignies, A. Alternative Lengthening of Telomeres Is Characterized by Reduced Compaction of Telomeric Chromatin. Nucleic Acids Res. 2014, 42, 4391–4405. [Google Scholar] [CrossRef]
- Voon, H.P.J.; Collas, P.; Wong, L.H. Compromised Telomeric Heterochromatin Promotes Alternative Lengthening of Telomeres. Trends Cancer 2016, 2, 114–116. [Google Scholar] [CrossRef]
- Yehezkel, S.; Segev, Y.; Viegas-Péquignot, E.; Skorecki, K.; Selig, S. Hypomethylation of Subtelomeric Regions in ICF Syndrome Is Associated with Abnormally Short Telomeres and Enhanced Transcription from Telomeric Regions. Hum. Mol. Genet. 2008, 17, 2776–2789. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Campbell, A.E.; Lieberman, P.M. TERRA, CpG Methylation and Telomere Heterochromatin: Lessons from ICF Syndrome Cells. Cell Cycle 2010, 9, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Sagie, S.; Toubiana, S.; Hartono, S.R.; Katzir, H.; Tzur-Gilat, A.; Havazelet, S.; Francastel, C.; Velasco, G.; Chédin, F.; Selig, S. Telomeres in ICF Syndrome Cells Are Vulnerable to DNA Damage Due to Elevated DNA:RNA Hybrids. Nat. Commun. 2017, 8, 14015. [Google Scholar] [CrossRef] [Green Version]
- Caslini, C.; Connelly, J.A.; Serna, A.; Broccoli, D.; Hess, J.L. MLL Associates with Telomeres and Regulates Telomeric Repeat-Containing RNA Transcription. Mol. Cell. Biol. 2009, 29, 4519–4526. [Google Scholar] [CrossRef] [Green Version]
- Caslini, C. Transcriptional Regulation of Telomeric Non-Coding RNA: Implications on Telomere Biology, Replicative Senescence and Cancer. RNA Biol. 2010, 7, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Caslini, C.; Alarcòn, A.; Hess, J.; Tanaka, R.; Murti, K.; Biondi, A. The Amino Terminus Targets the Mixed Lineage Leukemia (MLL) Protein to the Nucleolus, Nuclear Matrix and Mitotic Chromosomal Scaffolds. Leukemia 2000, 14, 1898–1908. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.C.; Dou, Y. Hijacked in Cancer: The KMT2 (MLL) Family of Methyltransferases. Nat. Rev. Cancer 2015, 15, 334–346. [Google Scholar] [CrossRef] [Green Version]
- Guenther, M.G.; Jenner, R.G.; Chevalier, B.; Nakamura, T.; Croce, C.M.; Canaani, E.; Young, R.A. Global and Hox-Specific Roles for the MLL1 Methyltransferase. Proc. Natl. Acad. Sci. USA 2005, 102, 8603–8608. [Google Scholar] [CrossRef] [Green Version]
- Caslini, C.; Yang, Z.; El-Osta, M.; Milne, T.A.; Slany, R.K.; Hess, J.L. Interaction of MLL Amino Terminal Sequences with Menin Is Required for Transformation. Cancer Res. 2007, 67, 7275–7283. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, A.; Cleary, M.L. Menin Critically Links MLL Proteins with LEDGF on Cancer-Associated Target Genes. Cancer Cell 2008, 14, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Tutton, S.; Azzam, G.A.; Stong, N.; Vladimirova, O.; Wiedmer, A.; Monteith, J.A.; Beishline, K.; Wang, Z.; Deng, Z.; Riethman, H.; et al. Subtelomeric P53 Binding Prevents Accumulation of DNA Damage at Human Telomeres. EMBO J. 2016, 35, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Scheibe, M.; Arnoult, N.; Kappei, D.; Buchholz, F.; Decottignies, A.; Butter, F.; Mann, M. Quantitative Interaction Screen of Telomeric Repeat-Containing RNA Reveals Novel TERRA Regulators. Genome Res. 2013, 23, 2149–2157. [Google Scholar] [CrossRef] [Green Version]
- Fujita, T.; Asano, Y.; Ohtsuka, J.; Takada, Y.; Saito, K.; Ohki, R.; Fujii, H. Identification of Telomere-Associated Molecules by Engineered DNA-Binding Molecule-Mediated Chromatin Immunoprecipitation (EnChIP). Sci. Rep. 2013, 3, 3171. [Google Scholar] [CrossRef] [Green Version]
- Muntean, A.G.; Hess, J.L. The Pathogenesis of Mixed-Lineage Leukemia. Annu. Rev. Pathol. 2012, 7, 283–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, C.; Burmeister, T.; Gröger, D.; Tsaur, G.; Fechina, L.; Renneville, A.; Sutton, R.; Venn, N.C.; Emerenciano, M.; Pombo-de-Oliveira, M.S.; et al. The MLL Recombinome of Acute Leukemias in 2017. Leukemia 2018, 32, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Slany, R.K. The Molecular Biology of Mixed Lineage Leukemia. Haematologica 2009, 94, 984–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monroe, S.C.; Jo, S.Y.; Sanders, D.S.; Basrur, V.; Elenitoba-Johnson, K.S.; Slany, R.K.; Hess, J.L. MLL-AF9 and MLL-ENL Alter the Dynamic Association of Transcriptional Regulators with Genes Critical for Leukemia. Exp. Hematol. 2011, 39, 77–86.e5. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Smith, E.R.; Takahashi, H.; Lai, K.C.; Martin-brown, S.; Florens, L.; Washburn, M.P.; Conaway, J.W.; Conaway, R.C.; Shilatifard, A. AFF4, a Component of the ELL/P-TEFb Elongation Complex and a Shared Subunit of MLL Chimeras, Can Link Transcription Elongation to Leukemia. Mol. Cell 2010, 37, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, A.; Lin, M.; Naresh, A.; Kitabayashi, I.; Cleary, M.L. A Higher-Order Complex Containing AF4 and ENL Family Proteins with P-TEFb Facilitates Oncogenic and Physiologic MLL-Dependent Transcription. Cancer Cell 2010, 17, 198–212. [Google Scholar] [CrossRef] [Green Version]
- Gobbi, A.; Di Berardino, C.; Scanziani, E.; Garofalo, A.; Rivolta, A.; Fontana, G.; Rambaldi, A.; Giavazzi, R.; Biondi, A. A Human Acute Lymphoblastic Leukemia Line with the T(4;11) Translocation as a Model of Minimal Residual Disease in SCID Mice. Leuk. Res. 1997, 21, 1107–1114. [Google Scholar] [CrossRef]
- Grant, J.D.; Broccoli, D.; Muquit, M.; Manion, F.J.; Tisdall, J.; Ochs, M.F. Telometric: A Tool Providing Simplified, Reproducible Measurements of Telomeric DNA from Constant Field Agarose Gels. Biotechniques 2001, 31, 1314–1316, 1318. [Google Scholar] [CrossRef] [Green Version]
- Broccoli, D.; Youngtt, J.W.; De Lange, T. Telomerase Activity in Normal and Malignant Hematopoietic Cells. Cell Biol. 1995, 92, 9082–9086. [Google Scholar] [CrossRef] [Green Version]
- Drexler, H.G.; Quentmeier, H.; MacLeod, R.A.F. Malignant Hematopoietic Cell Lines: In Vitro Models for the Study of MLL Gene Alterations. Leukemia 2004, 18, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Sampl, S.; Pramhas, S.; Stern, C.; Preusser, M.; Marosi, C.; Holzmann, K. Expression of Telomeres in Astrocytoma WHO Grade 2 to 4: TERRA Level Correlates with Telomere Length, Telomerase Activity, and Advanced Clinical Grade. Transl. Oncol. 2012, 5, 56-IN4. [Google Scholar] [CrossRef] [Green Version]
- Vitelli, V.; Falvo, P.; Nergadze, S.G.; Santagostino, M.; Khoriauli, L.; Pellanda, P.; Bertino, G.; Occhini, A.; Benazzo, M.; Morbini, P.; et al. Telomeric Repeat-Containing RNAs (TERRA) Decrease in Squamous Cell Carcinoma of the Head and Neck Is Associated with Worsened Clinical Outcome. Int. J. Mol. Sci. 2018, 19, 274. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Zhai, Y.; Ji, X.; Wang, Y.; Zhao, J.; Xing, J.; An, J.; Ren, T. Noncoding Telomeric Repeat-containing RNA Inhibits the Progression of Hepatocellular Carcinoma by Regulating Telomerase-mediated Telomere Length. Cancer Sci. 2020, 111, 2789–2802. [Google Scholar] [CrossRef]
- Adishesh, M.; Alnafakh, R.; Baird, D.M.; Jones, R.E.; Simon, S.; Button, L.; Kamal, A.M.; Kirwan, J.; DeCruze, S.B.; Drury, J.; et al. Human Endometrial Carcinogenesis Is Associated with Significant Reduction in Long Non-Coding RNA, TERRA. Int. J. Mol. Sci. 2020, 21, 8686. [Google Scholar] [CrossRef]
- Liu, H.; Cheng, E.H.Y.; Hsieh, J.J.D. Bimodal Degradation of MLL by SCFSkp2 and APCCdc20 Assures Cell Cycle Execution: A Critical Regulatory Circuit Lost in Leukemogenic MLL Fusions. Genes Dev. 2007, 21, 2385–2398. [Google Scholar] [CrossRef] [Green Version]
- Porro, A.; Feuerhahn, S.; Reichenbach, P.; Lingner, J. Molecular Dissection of Telomeric Repeat-Containing RNA Biogenesis Unveils the Presence of Distinct and Multiple Regulatory Pathways. Mol. Cell. Biol. 2010, 30, 4808–4817. [Google Scholar] [CrossRef] [Green Version]
- Blobel, G.A.; Kadauke, S.; Wang, E.; Lau, A.W.; Zuber, J.; Chou, M.M.; Vakoc, C.R. A Reconfigured Pattern of MLL Occupancy within Mitotic Chromatin Promotes Rapid Transcriptional Reactivation Following Mitotic Exit. Mol. Cell 2009, 36, 970–983. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, Y.B.; Pirrotta, V. Polycomb Silencing Mechanisms and the Management of Genomic Programmes. Nat. Rev. Genet. 2007, 8, 9–22. [Google Scholar] [CrossRef]
- Yehezkel, S.; Shaked, R.; Sagie, S.; Berkovitz, R.; Shachar-Bener, H.; Segev, Y.; Selig, S. Characterization and Rescue of Telomeric Abnormalities in ICF Syndrome Type I Fibroblasts. Front. Oncol. 2013, 3, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Mullighan, C.G.; Goorha, S.; Radtke, I.; Miller, C.B.; Coustan-Smith, E.; Dalton, J.D.; Girtman, K.; Mathew, S.; Ma, J.; Pounds, S.B.; et al. Genome-Wide Analysis of Genetic Alterations in Acute Lymphoblastic Leukaemia. Nature 2007, 446, 758–764. [Google Scholar] [CrossRef]
- Bardini, M.; Spinelli, R.; Bungaro, S.; Mangano, E.; Corral, L.; Cifola, I.; Fazio, G.; Giordan, M.; Basso, G.; De Rossi, G.; et al. DNA Copy-Number Abnormalities Do Not Occur in Infant ALL with t(4;11)/MLL-AF4. Leukemia 2010, 24, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.K.; Ma, J.; Wang, J.; Chen, X.; Gedman, A.L.; Dang, J.; Nakitandwe, J.; Holmfeldt, L.; Parker, M.; Easton, J.; et al. The Landscape of Somatic Mutations in Infant MLL-Rearranged Acute Lymphoblastic Leukemias. Nat. Genet. 2015, 47, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Beishline, K.; Vladimirova, O.; Tutton, S.; Wang, Z.; Deng, Z.; Lieberman, P.M. CTCF Driven TERRA Transcription Facilitates Completion of Telomere DNA Replication. Nat. Commun. 2017, 8, 2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Cell Line | Clinical Data b | Cytogenetic | Fusion Gene | Karyotype | Ploidy e | |
|---|---|---|---|---|---|---|---|
| non-MLL-rearranged leukemia | |||||||
| 1 | Chronic myeloid leukemia | K-562 | CML, 53 F, PE | t(9;22)(q34;q11) | BCR-ABL d | Hypotriploid—64(61–68)<3n> | 2.8 |
| 2 | Chronic myeloid leukemia | NALM-1 a | CML, 3 F, PB | t(9;22)(q34;q12) c | BCR-ABL | hypodiploid with 5% polyploidy—45(42–47)<2n> | 2.0 |
| 3 | Acute myeloid leukemia | ME-1 a | AML, M4eo, 40 M, PB | inv(16)(p13;q22) | CBFb-MYH11 d | near diploid—47<2n> | 2.0 |
| 4 | Acute myeloid leukemia | KASUMI-1 a | AML, M2, 7 M, PB | t(8;21)(q22;q22) c | AML1-ETO d | Hypodiploid—45<2n> | 2.0 |
| 5 | Acute myeloid leukemia | U-937 | AML, M5, 37 M, PE | t(10;11)(p13;q14) | CALM-AF10 | Hypotriploid—63(58–69)<3n> | 2.7 |
| 6 | APL, Acute promyelocytic leukemia | NB-4 a | AML, M3, 23 F, BM | t(15;17)(q22;q11-12) c | PML-RARa d | hypertriploid with 3% polyploidy—78(71–81)<3n> | 3.4 |
| 7 | B cell precursor leukemia | 697 a | pre-B ALL, 12 M, BM | t(1:19)(q23;p13) c | E2A-PBX d | near diploid—46(45–48)<2n> | 2.0 |
| 8 | B cell precursor leukemia | NALM-6 a | pre-B ALL, 19 M, PB | t(5;12)(q33;p13) c | TEL-PDGFRb | near diploid—46(4–47)<2n> | 2.0 |
| 9 | B cell precursor leukemia | REH a | pre-B ALL, 15 F, PB | t(12;21)(p13;q22) c | TEL-AML1 d | Pseudodiploid—46(44–47)<2n> | 2.0 |
| 10 | B cell precursor leukemia | HAL-01 a | pre-B ALL, 17 F, PB | t(17;19)(q22;p13) c | E2A-HLF d | near diploid with 4% polyploidy; 46(43–48)<2n> | 2.0 |
| 11 | Burkitt lymphoma | RAJI a | Burkitt Lymphoma, 12 M, LM | t(8;14)(q24;q32) c | IgH-cMYC | hypotetraploid with 12% polyploidy—89(80–91)<4n> | 3.9 |
| 12 | T cell leukemia | JURKAT a | T-ALL, 14 M, PB | add(2)(p21)/del(2)(p23)x2 | - | hypotetraploid karyotype with 7.8% polyploidy—87(78–91)<4n> | 3.8 |
| MLL-rearranged leukemia | |||||||
| 13 | Acute myeloid (eosinophilic) leukemia | EOL-1 a | AML, 33 M, PB | del(4)(q12)x2 c | FIP1L1-PDGFRa & MLL-PTD | hyperdiploid with 7.5% polyploidy—50(48–51)<2n> | 2.2 |
| 14 | Acute monocytic leukemia | KOPM-88 | AML, 11 M, PB | MLL-PTD | MLL-PTD | near tetraploid—91<4n> | 4.0 |
| 15 | Acute monocytic leukemia | MV4;11 a | AML, M5, 10 M, PB | t(4;11)(q21;q23) c | MLL-AF4 d | Hyperdiploid—48(46–48)<2n> | 2.1 |
| 16 | Acute monocytic leukemia | MONO-MAC-6 a | AML, M5, 64 M, PB | t(9;11)(p22;q23) c | MLL-AF9 | hypotetraploid with near-diploid (8%) and polyploid (17%) sidelines—84–90<4n> | 3.8 |
| 17 | Acute monocytic leukemia | THP-1 a | AML, 1 M, PB | t(9;11)(p22;q23) c | MLL-AF9 | near-tetraploid—94(88–96)<4n> | 4.1 |
| 18 | Acute myelomonocytic leukemia | ML-2 a | AML, M4, 26 M, PB | t(6;11)(q27;q23) c | MLL-AF6 d | near tetraploid—92(84–94)<4n> | 4.0 |
| 19 | B cell precursor leukemia | KOPN-8 a | pre-B ALL, <1 F, PB | t(11;19)(q23;p13) c | MLL-ENL d | hypodiploid with 4% polyploidy—45(42–45)<2n> | 2.0 |
| 20 | B cell precursor leukemia | ALL-PO | pre-B ALL, <1 F, BM | t(4;11)(q21;q23) | MLL-AF4 | hyperdiploid with 20% polyploidy—57<2n> (*) | 2.5 |
| 21 | B cell precursor leukemia | RS4;11 | pre-B ALL, 32 F, BM | t(4;11)(q21;q23) c | MLL-AF4 d | Hyperdiploid—47/48<2n> | 2.0 |
| 22 | B cell precursor leukemia | SEM a | pre-B ALL, 5 F, PB | t(4;11)(q21;q23) c | MLL-AF4 d | hypodiploid with 1.5% polyploidy—45(40–46)<2n> | 2.0 |
| 23 | T cell leukemia | KARPAS-45 a | T-ALL, 2 M, BM | t(X;11)(q13;q23) c | MLL-AFX1 | hypotetraploid with 8% polyploidy—87(84–88)<4n> | 3.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caslini, C.; Serna, A. Telomere Transcription in MLL-Rearranged Leukemia Cell Lines: Increased Levels of TERRA Associate with Lymphoid Lineage and Are Independent of Telomere Length and Ploidy. Biomedicines 2023, 11, 925. https://doi.org/10.3390/biomedicines11030925
Caslini C, Serna A. Telomere Transcription in MLL-Rearranged Leukemia Cell Lines: Increased Levels of TERRA Associate with Lymphoid Lineage and Are Independent of Telomere Length and Ploidy. Biomedicines. 2023; 11(3):925. https://doi.org/10.3390/biomedicines11030925
Chicago/Turabian StyleCaslini, Corrado, and Amparo Serna. 2023. "Telomere Transcription in MLL-Rearranged Leukemia Cell Lines: Increased Levels of TERRA Associate with Lymphoid Lineage and Are Independent of Telomere Length and Ploidy" Biomedicines 11, no. 3: 925. https://doi.org/10.3390/biomedicines11030925