1. Introduction
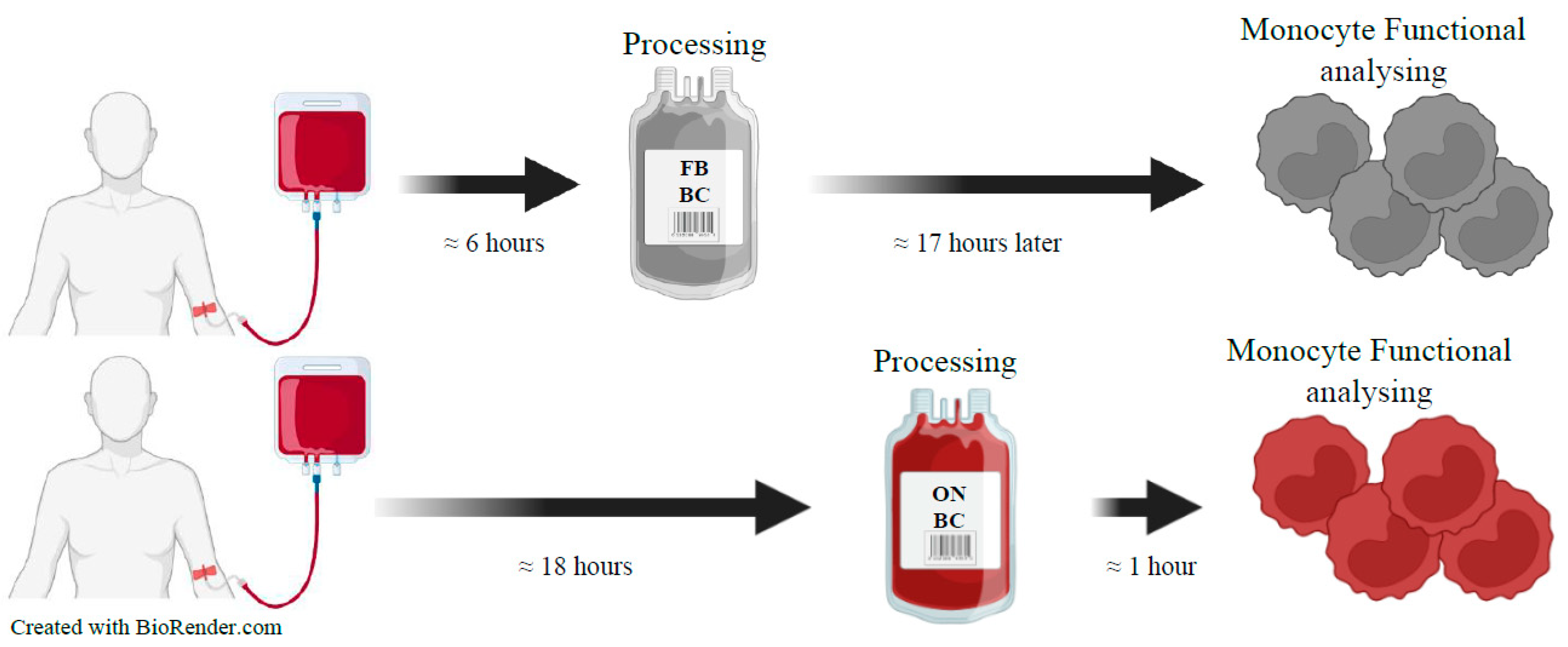
At the blood bank, the processing of whole blood, a 450 mL donation, originates red cells, platelet units, plasma, and a waste product also known as Buffy Coat (BC). BCs may be obtained by two different protocols according to the time interval between blood collection and processing: (1) Fresh Blood BC, in which the processing of the whole blood is performed on the same day as the blood donation and (2) Overnight Blood BC, in which the whole blood is processed in the day after the blood collection (
Figure 1).
The BC is a reliable, consistent, and widely used source of leukocytes, including monocytes, for biomedical research purposes. In healthy donors, reference ranges of different cell populations percentages present in BCs are: granulocytes 42–77%; lymphocytes 20–44%, and monocytes 2–9.5% [
1].
Monocytes are instrumental for several events that happen in the immune system. They are the primary type of mononuclear phagocyte in blood, and they are traditionally looked at as a source of tissue macrophages. However, recent studies underline a more complex role for monocytes in innate and adaptative immunity, particularly as antigen presenting cells [
2].
Monocytes act as antigen-presenting cells to both conventional and unconventional T cells, including invariant NKT (iNKT) cells. iNKT cells are unconventional T cells that are reactive to the nonclassical major histocompatibility complex (MHC) class I-like antigen-presenting molecule CD1d and are characterized by the expression of a semi-invariant TCR, which, in humans, combines a Vα24Jα18 chain paired with a Vβ11 chain [
3,
4]. CD1d is a non-polymorphic molecule facilitating the allogenic use of iNKT cells in immunotherapies [
4,
5]. iNKT cells are an important lymphocyte population that sense a range of endogenous and exogenous lipid antigens presented by CD1d. These cells recognize glycolipid antigens, namely α-Galactosylceramide (α-GalCer), which is the prototype lipid antigen for iNKT cells. Other endogenous and exogenous iNKT cell lipid antigens have been discovered [
3,
4]. The lipid α-Gal-(α1-2)-α-GalCer (α-GalGalCer) becomes an iNKT cell antigen upon internalization and galactose removal by α-galactosidase A in the lysosome [
6]. Upon stimulation, iNKT cells respond rapidly to both innate signals and TCR engagement by promptly producing large amounts of cytokines, which are important in the modulation of several immune responses against infection and cancer [
4,
5].
Monocytes are frequently used in immunological and pharmacological studies, diagnostics, and clinical trials. These assays depend on the physiological state of monocytes. Pre-analytical factors, such as blood collection technique (different tubes and anticoagulants), blood storage, monocyte isolation technique, and freezing may directly interfere with monocyte properties. For instance, monocyte CD14 expression is affected by the type of anticoagulant used in blood collection [
7]. Few studies have been conducted to assess the effect of pre-analytical factors on monocyte properties. Longer delays in blood processing lead to increased granulocyte contamination of Peripheral Blood Mononuclear Cells (PBMCs), a decrease in NK cells’ ability to degranulate, and low cell recovery and affects PBMCs’ proteomic patterns [
8,
9].
In the present study, we investigated whether the antigen-presentation capacity of monocytes to present lipid antigens to iNKT cells is influenced by different BC processing protocols.
2. Materials and Methods
Harvest of the whole blood for both Fresh Blood BC and Overnight Blood BC was completed on the same day from different donors. Whole blood collection units (450 mL) were maintained at room temperature for approximately 6 h for the units that would give rise to Fresh Blood BC and 18 h for the units that would generate Overnight Blood BC. Afterwards, the blood units, from different subjects, were processed into the different products (Red blood cells, plasma, platelets, and BC) at the hospital using the Reveos® system by centrifugation at 2600 or 2900 rpm (depending on the blood product desired) for 7 min. Then, PBMCs were isolated at the same time, in our laboratory, from the BC obtained by the two different protocols: Fresh Blood BC and Overnight Blood BC.
All the experimental protocols were performed using BC kindly provided by the Blood Bank of the Centro Hospitalar Universitário de São João, Porto (Portugal).
2.1. Peripheral Blood Mononuclear Cells (PBMCs) Isolation
PBMCs were isolated from the BC using Histopaque-1077® (Sigma-Aldrich, St. Louis, MO, USA) density centrifugation following the manufacturer’s instructions. After centrifugation, the PBMCs ring, located between the plasma and Histopaque-1077®, was recovered, followed by washing with PBS. Incubation with Red Blood Cell Lysis Buffer (BioLegend, San Diego, CA, USA) was performed to lysate remaining erythrocytes, and, after wash and cell count, PBMCs were used for CD14+ cells isolation.
2.2. Monocytes Isolation through Positive Selection
Monocytes were isolated by positive selection with anti-CD14 magnetic beads using the MACS cell separation system (MiltenyiBiotec, Cologne, Germany) following the manufacturer’s instructions. Succinctly, the PBMCs were incubated with magnetic beads coated with the anti-CD14 antibody and passed through a magnetic column. The magnetic force existing between the beads and the support keeps the labeled cells from eluting. The cells that bind to the beads (monocytes) remain in the column, while non-labeled cells are eluted. When the column is removed from the magnetic support, the CD14+ cells can be eluted. CD14+ cells were used, after purification, for flow cytometry analysis and lipid antigen presentation assays.
2.3. iNKT Cell Line Culture Maintenance and Re-Stimulation
An iNKT cell line was generated and maintained through re-stimulations as previously described [
10]. In short, the iNKT cell line was generated by culturing PBMCs with 100 ng/mL of α-GalCer (KRN7000) and 100 U/mL of IL-2. These PBMCs, and consequently the iNKT cell line, were derived from a healthy individual. After 11 days, CD1d-PBS57 tetramer
+ CD3
+ cells were sorted using FACSAria (BD Biosciences). iNKT cells were maintained in culture at 37 °C, 5% CO
2 by periodic re-stimulation with irradiated PBMCs in the presence of 1 μg/mL of PHA (Thermo Fisher Scientific, Waltham, MA, USA), and 100 U/mL of IL-2.
2.4. Flow Cytometry
Monocyte purity and the basal state of activation of cells were assessed by flow cytometry using the following anti-human monoclonal antibodies: CD14 (61D3, eBiosciences, San Diego, CA, USA), CD1d (51.1, Biolegend San Diego, CA, USA), CD80 (2D10, Biolegend, San Diego, CA, USA), and CD40 (5C3, BioLegend San Diego, CA, USA). Monocyte viability was assessed by flow cytometry using Fixable Viability Dye (FVD—eBiosciences, San Diego, CA, USA). Cells were stained with the previously mentioned antibody cocktails diluted in PBS/2%FBS/1mM EDTA/0.01%NaN3 (flow cytometry solution) for 20 min, at 4 °C, in the dark. After staining, cells were washed with flow cytometry solution and then fixed with PBS 1% formaldehyde. The purity of the iNKT cell line was tested using CD1d-PBS57 tetramers (NIH Tetramer Core Facility, Emory University, Atlanta, GA, USA) and anti-human CD3 (UCHT1, eBioscience, San Diego, CA, USA). Cells were acquired in a FACS Canto II (BD Biosciences, San Diego, CA, USA) using the BD FACS DivaTM software (BD Biosciences). Data analysis was performed with FlowJoR v10 (FlowJo LLC, Ashland, OR, USA).
2.5. Lipid Antigen Presentation Assays
Monocytes were cultured with α-Galactosylceramide (α-GalCer) (at 3.1, 12.5, or 50 ng/mL, KRN700, Sigma) or Gal(α1-2) αGalactosylceramide (α-GalGalCer or PBS 18, provided by Paul B. Savage from Brigham Young University, Provo, UT, USA; PBS 18 stands for the internal classification of the lipid synthetized by Paul B. Savage) (at 50, 150, or 300 ng/mL) and Phytohemagglutinin (PHA, Thermo Fisher Scientific at 1 μg/mL). α-GalGalCer was first dissolved in PBS 0.5% Tween 20 and then diluted in non-supplemented RPMI to have a maximum of 1% vehicle in culture. α-GalCer was resuspended in PBS and directly diluted in non-supplemented RPMI. After 4 h, an iNKT cell line was added, and cells were cocultured for 40 h at 37 °C, 5% CO2. After 40 h, supernatants were collected for cytokine production determination by ELISA. The following antibody pairs from Biolegend were used: purified anti-human GM-CSF (BVD2-23B6) and purified anti-human IL-4 (8D4-8).
2.6. Statistics
An unpaired t-test (normal distribution) was used to compare two groups. p-values lower than 0.05 were considered statistically significant. Along with the comparison of the raw data, and due to the inter-experimental variation, comparison was also performed after normalization of the values of cytokine production for each independent experiment. The values of cytokine production were normalized, considering 100 as the highest cytokine production value within each experiment for the chosen antigen concentration. All the analyses were performed using GraphPad Prism software v6 (GraphPad Software Inc., San Diego, CA, USA).
4. Conclusions
In summary, we show that in lipid antigen presentation assays monocytes from Fresh Blood BC demonstrated a higher capacity to activate iNKT cells than Overnight Blood BC, leading to the conclusion that functional differences exist in monocytes from different time-processed BCs. The BC continues to prove to be a reliable and regular/conventional source of blood cells, therefore this sample is of great importance in research dealing with blood components.
In conclusion, BC-processing protocols influence monocytes’ function in their capacity to present antigens. As such, the protocol for BC processing should be carefully considered in daily immunological activation assay research.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




