
The Potential Role of Serum and Exhaled Breath Condensate miRNAs in Diagnosis and Predicting Exacerbations in Pediatric Asthma


Abstract
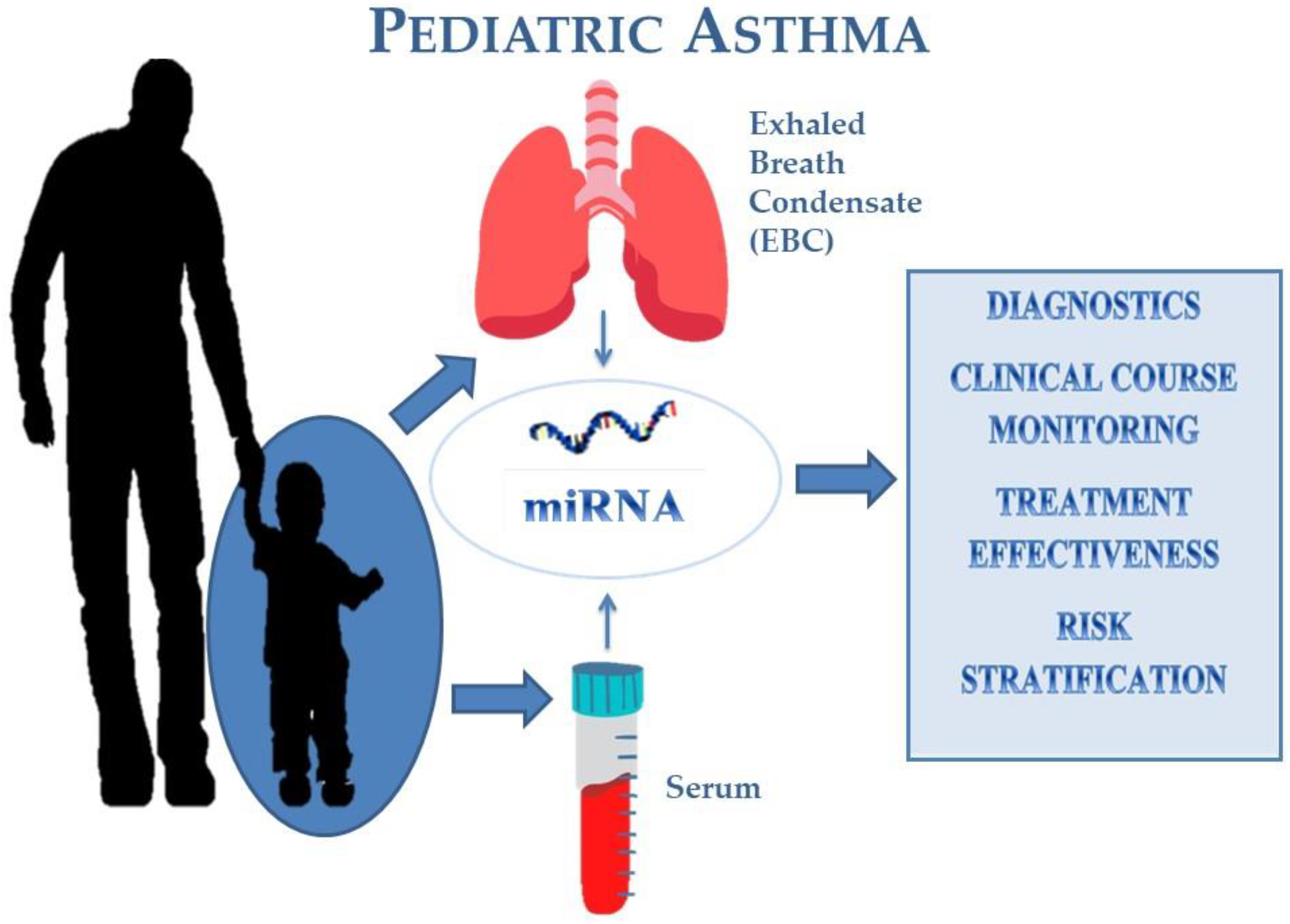
:1. Introduction
2. The Diagnostic Role of miRNAs
3. Asthma Exacerbation and miRNAs
4. Exhaled Breath Condensate
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gutierrez, M.J.; Gomez, J.L.; Perez, G.F.; Pancham, K.; Val, S.; Pillai, D.K.; Giri, M.; Ferrante, S.; Freishtat, R.; Rose, M.C.; et al. Airway Secretory MicroRNAome Changes during Rhinovirus Infection in Early Childhood. PLoS ONE 2016, 11, e0162244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kho, A.T.; McGeachie, M.J.; Moore, K.G.; Sylvia, J.M.; Weiss, S.T.; Tantisira, K.G. Circulating MicroRNAs and Prediction of Asthma Exacerbation in Childhood Asthma. Respir. Res. 2018, 19, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Tiwari, A.; McGeachie, M.J. Recent MiRNA Research in Asthma. Curr. Allergy Asthma Rep. 2022, 22, 231–258. [Google Scholar] [CrossRef] [PubMed]
- Taka, S.; Tzani-Tzanopoulou, P.; Wanstall, H.; Papadopoulos, N.G. MicroRNAs in Asthma and Respiratory Infections: Identifying Common Pathways. Allergy Asthma Immunol. Res. 2020, 12, 4. [Google Scholar] [CrossRef] [PubMed]
- Weidner, J.; Bartel, S.; Kılıç, A.; Zissler, U.M.; Renz, H.; Schwarze, J.; Schmidt-Weber, C.B.; Maes, T.; Rebane, A.; Krauss-Etschmann, S.; et al. Spotlight on MicroRNAs in Allergy and Asthma. Allergy 2021, 76, 1661–1678. [Google Scholar] [CrossRef]
- Lizzo, J.M.; Cortes, S. Pediatric Asthma; Mayo Clinic Press: Rochester, MN, USA, 2022. [Google Scholar]
- Pattarayan, D.; Thimmulappa, R.K.; Ravikumar, V.; Rajasekaran, S. Diagnostic Potential of Extracellular MicroRNA in Respiratory Diseases. Clin. Rev. Allergy Immunol. 2018, 54, 480–492. [Google Scholar] [CrossRef]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.A.; Elia Adil, N.; Ahmed, M.S.E. Diagnosis of Asthma in Childhood Age. Arch. Asthma Allergy Immunol. 2018, 2, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.L.; Gaffin, J.M.; Radhakrishnan, D. Question 3: Can We Diagnose Asthma in Children under the Age of 5 years? Paediatr. Respir. Rev. 2019, 29, 25–30. [Google Scholar] [CrossRef]
- Liu, F.; Qin, H.-B.; Xu, B.; Zhou, H.; Zhao, D.-Y. Profiling of MiRNAs in Pediatric Asthma: Upregulation of MiRNA-221 and MiRNA-485-3p. Mol. Med. Rep. 2012, 6, 1178–1182. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Bai, M.; Deng, L.; Fan, L.; Wang, X. MiRNA-21 Regulates Bronchial Epithelial Cell Proliferation by Activating Tgfβ1/Smad Signaling Pathway and Its Correlation with Asthma Severity in Children. Iran. J. Public Health 2021, 50, 1973–1982. [Google Scholar] [CrossRef] [PubMed]
- Atashbasteh, M.; Mortaz, E.; Mahdaviani, S.A.; Jamaati, H.; Allameh, A. Expression Levels of Plasma Exosomal MiR-124, MiR-125b, MiR-133b, MiR-130a and MiR-125b-1-3p in Severe Asthma Patients and Normal Individuals with Emphasis on Inflammatory Factors. Allergy Asthma Clin. Immunol. 2021, 17, 51. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Lu, W.; Li, C.; Peng, X.; Wang, Y.; Huang, X.; Yao, Z.; Cai, N.; Huang, Y.; Zhang, X.; et al. MiR-3162-3p Is a Novel MicroRNA That Exacerbates Asthma by Regulating β-Catenin. PLoS ONE 2016, 11, e0149257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yang, L.; Li, P.; Huang, H.; Liu, T.; He, H.; Lin, Z.; Jiang, Y.; Ren, N.; Wu, B.; et al. Circulating MicroRNA Signatures Associated with Childhood Asthma. Clin. Lab. 2015, 61, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Januskevicius, A.; Vaitkiene, S.; Gosens, R.; Janulaityte, I.; Hoppenot, D.; Sakalauskas, R.; Malakauskas, K. Eosinophils Enhance WNT-5a and TGF-Β1 Genes Expression in Airway Smooth Muscle Cells and Promote Their Proliferation by Increased Extracellular Matrix Proteins Production in Asthma. BMC Pulm. Med. 2016, 16, 94. [Google Scholar] [CrossRef] [Green Version]
- Cañas, J.A.; Sastre, B.; Rodrigo-Muñoz, J.M.; del Pozo, V. Exosomes: A New Approach to Asthma Pathology. Clin. Chim. Acta 2019, 495, 139–147. [Google Scholar] [CrossRef]
- He, L.; Liu, J.; Wang, X.; Wang, Y.; Zhu, J.; Kang, X. Identifying a Novel Serum MicroRNA Biomarker Panel for the Diagnosis of Childhood Asthma. Exp. Biol. Med. 2022, 247, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, É.; Madore, A.-M.; Boucher-Lafleur, A.-M.; Simon, M.-M.; Kwan, T.; Pastinen, T.; Laprise, C. Eosinophil MicroRNAs Play a Regulatory Role in Allergic Diseases Included in the Atopic March. Int. J. Mol. Sci. 2020, 21, 9011. [Google Scholar] [CrossRef]
- Rodrigo-Muñoz, J.M.; Gil-Martínez, M.; Lorente-Sorolla, C.; García-Latorre, R.; Valverde-Monge, M.; Quirce, S.; Sastre, J.; del Pozo, V. MiR-144-3p Is a Biomarker Related to Severe Corticosteroid-Dependent Asthma. Front. Immunol. 2022, 13, 858722. [Google Scholar] [CrossRef]
- Maes, T.; Cobos, F.A.; Schleich, F.; Sorbello, V.; Henket, M.; de Preter, K.; Bracke, K.R.; Conickx, G.; Mesnil, C.; Vandesompele, J.; et al. Asthma Inflammatory Phenotypes Show Differential MicroRNA Expression in Sputum. J. Allergy Clin. Immunol. 2016, 137, 1433–1446. [Google Scholar] [CrossRef] [Green Version]
- Huo, X.; Zhang, K.; Yi, L.; Mo, Y.; Liang, Y.; Zhao, J.; Zhang, Z.; Xu, Y.; Zhen, G. Decreased Epithelial and Plasma MiR-181b-5p Expression Associates with Airway Eosinophilic Inflammation in Asthma. Clin. Exp. Allergy 2016, 46, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Elnady, H.G.; Sherif, L.S.; Kholoussi, N.M.; Ali Azzam, M.; Foda, A.R.; Helwa, I.; Sabry, R.N.; Eissa, E.; Fahmy, R.F. Aberrant Expression of Immune-Related MicroRNAs in Pediatric Patients with Asthma. Int. J. Mol. Cell Med. 2020, 9, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.; Hobbs, B.D.; Li, J.; Kho, A.T.; Amr, S.; Celedón, J.C.; Weiss, S.T.; Hersh, C.P.; Tantisira, K.G.; McGeachie, M.J. Blood MiRNAs Are Linked to Frequent Asthma Exacerbations in Childhood Asthma and Adult COPD. Noncoding RNA 2022, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.A.; Ramadan, A.; Wahby, A.A.; Draz, I.H.; El Baroudy, N.R.; Abdel Hamid, T.A. Evaluation of MiR-196a2 Expression and Annexin A1 Level in Children with Bronchial AsthmaEvaluation of MiR-196a2 Expression and Annexin A1 Level in Children. Allergol. Immunopathol. 2020, 48, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Lee, P.-H.; Kim, B.-G.; Seo, H.-J.; Baek, A.-R.; Park, J.-S.; Lee, J.-H.; Park, S.-W.; Kim, D.-J.; Park, C.-S.; et al. Annexin A1 in Plasma from Patients with Bronchial Asthma: Its Association with Lung Function. BMC Pulm. Med. 2018, 18, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, M.H.; Toraih, E.A.; Aly, N.M.; Riad, E.; Fawzy, M.S. A Passenger Strand Variant in MiR-196a2 Contributes to Asthma Severity in Children and Adolescents: A Preliminary Study. Biochem. Cell Biol. 2016, 94, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Jartti, T.; Gern, J.E. Role of Viral Infections in the Development and Exacerbation of Asthma in Children. J. Allergy Clin. Immunol. 2017, 140, 895–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laanesoo, A.; Urgard, E.; Periyasamy, K.; Laan, M.; Bochkov, Y.A.; Aab, A.; Magilnick, N.; Pooga, M.; Gern, J.E.; Johnston, S.L.; et al. Dual Role of the MiR-146 Family in Rhinovirus-induced Airway Inflammation and Allergic Asthma Exacerbation. Clin. Transl. Med. 2021, 11, e427. [Google Scholar] [CrossRef]
- Moheimani, F.; Koops, J.; Williams, T.; Reid, A.T.; Hansbro, P.M.; Wark, P.A.; Knight, D.A. Influenza A Virus Infection Dysregulates the Expression of MicroRNA-22 and Its Targets; CD147 and HDAC4, in Epithelium of Asthmatics. Respir. Res. 2018, 19, 145. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wang, W.; Jing, W. Indoor Air Pollution Aggravates Asthma in Chinese Children and Induces the Changes in Serum Level of MiR-155. Int. J. Environ. Health Res. 2019, 29, 22–30. [Google Scholar] [CrossRef]
- Xiao, R.; Noël, A.; Perveen, Z.; Penn, A.L. In Utero Exposure to Second-Hand Smoke Activates pro-Asthmatic and Oncogenic MiRNAs in Adult Asthmatic Mice. Environ. Mol. Mutagen. 2016, 57, 190–199. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, A.; Wang, A.L.; Li, J.; Lutz, S.M.; Kho, A.T.; Weiss, S.T.; Tantisira, K.G.; McGeachie, M.J. Seasonal Variation in MiR-328-3p and Let-7d-3p Are Associated With Seasonal Allergies and Asthma Symptoms in Children. Allergy Asthma Immunol. Res. 2021, 13, 576. [Google Scholar] [CrossRef]
- Liang, J.; Liu, X.-H.; Chen, X.-M.; Song, X.-L.; Li, W.; Huang, Y. Emerging Roles of Non-Coding RNAs in Childhood Asthma. Front. Pharm. 2022, 13, 856104. [Google Scholar] [CrossRef] [PubMed]
- Kho, A.T.; Sharma, S.; Davis, J.S.; Spina, J.; Howard, D.; McEnroy, K.; Moore, K.; Sylvia, J.; Qiu, W.; Weiss, S.T.; et al. Circulating MicroRNAs: Association with Lung Function in Asthma. PLoS ONE 2016, 11, e0157998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadorp, B.; Soreq, H. Predicted Overlapping MicroRNA Regulators of Acetylcholine Packaging and Degradation in Neuroinflammation-Related Disorders. Front. Mol. Neurosci. 2014, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Qi, R.; Xu, J.; Di, Z.; Zheng, H.; Huo, W.; Zhang, L.; Chen, H.; Gao, X. Profiling of Serum and Urinary MicroRNAs in Children with Atopic Dermatitis. PLoS ONE 2014, 9, e115448. [Google Scholar] [CrossRef]
- Tiwari, A.; Li, J.; Kho, A.T.; Sun, M.; Lu, Q.; Weiss, S.T.; Tantisira, K.G.; McGeachie, M.J. COPD-Associated MiR-145-5p Is Downregulated in Early-Decline FEV1 Trajectories in Childhood Asthma. J. Allergy Clin. Immunol. 2021, 147, 2181–2190. [Google Scholar] [CrossRef] [PubMed]
- Mendes, F.C.; Paciência, I.; Ferreira, A.C.; Martins, C.; Rufo, J.C.; Silva, D.; Cunha, P.; Farraia, M.; Moreira, P.; Delgado, L.; et al. Development and Validation of Exhaled Breath Condensate MicroRNAs to Identify and Endotype Asthma in Children. PLoS ONE 2019, 14, e0224983. [Google Scholar] [CrossRef] [Green Version]
- Mendes, F.C.; Paciência, I.; Cavaleiro Rufo, J.; Silva, D.; Delgado, L.; Moreira, A.; Moreira, P. Dietary Acid Load Modulation of Asthma-Related MiRNAs in the Exhaled Breath Condensate of Children. Nutrients 2022, 14, 1147. [Google Scholar] [CrossRef]
- Szymczak, I.; Wieczfinska, J.; Pawliczak, R. Molecular Background of MiRNA Role in Asthma and COPD: An Updated Insight. Biomed Res. Int. 2016, 2016, 7802521. [Google Scholar] [CrossRef] [Green Version]
- Sinha, A.; Yadav, A.K.; Chakraborty, S.; Kabra, S.K.; Lodha, R.; Kumar, M.; Kulshreshtha, A.; Sethi, T.; Pandey, R.; Malik, G.; et al. Exosome-Enclosed MicroRNAs in Exhaled Breath Hold Potential for Biomarker Discovery in Patients with Pulmonary Diseases. J. Allergy Clin. Immunol. 2013, 132, 219–222.e7. [Google Scholar] [CrossRef] [PubMed]
- Roff, A.N.; Craig, T.J.; August, A.; Stellato, C.; Ishmael, F.T. MicroRNA-570-3p Regulates HuR and Cytokine Expression in Airway Epithelial Cells. Am. J. Clin. Exp. Immunol. 2014, 3, 68–83. [Google Scholar] [PubMed]
- Tiotiu, A. Biomarkers in Asthma: State of the Art. Asthma Res. Pract. 2018, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pite, H.; Morais-Almeida, M.; Mensinga, T.; Diamant, Z. Non-Invasive Biomarkers in Asthma: Promises and Pitfalls. In Asthma—From Childhood Asthma to ACOS Phenotypes; InTech: London, UK, 2016. [Google Scholar]
- James, A.; Hedlin, G. Biomarkers for the Phenotyping and Monitoring of Asthma in Children. Curr. Treat Options Allergy 2016, 3, 439–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Study | Population | miRNA | Gene | Biological Function |
|---|---|---|---|---|
| F. Liu et al. [11] | N = 12 (study with control group) Age: 4–6 years SAMPLE: serum | ↑ miRNA-221 ↑ miRNA-485-3p | SPRED2 | Decreased Spred-2 protein. Increased cell proliferation and eosinophilic inflammatory response in the airways through IL-5. |
| Y. Kang et al. [12] | N = 103 (study with control group) STUDY GROUP: ASTHMA Age: 9.3 ± 0.4 years CONTROL GROUP Age 9.6 ± 0.4 years SAMPLE: serum | ↑ miRNA-21 | Smad7 | Decreased Smad7 protein. TGFβ1/Smad pathway. Excessive synthesis of collagen, α-smooth muscle actin, proliferation and differentiation of fibroblasts. Increased airway obstruction. |
| Y. Wang et al. [14] | N = 54 (study with control group) SAMPLE: serum | ↑ miRNA-3162-3p ↑ miRNA-1260a, ↑ miRNA-let-7c-5p ↑ miRNA-494 | CTNNB1 | Decreased β-catenin. Intensification of airway hyperreactivity and inflammation. |
| H. Elnady et al. [23] | N = 50 (study with control group) STUDY GROUP: ASTHMA Age: 10.6 ± 0.7 years CONTROL GROUP Age: 11.0 ± 0.8 years SAMPLE: serum | ↑ miRNA-146a ↑ miRNA-106b | - | Increased IL-5, IL-13. Stimulate inflammatory cell recruitment, epithelial cell and smooth muscle hyperplasia, goblet cell metaplasia, and extracellular matrix deposition in the respiratory tract. |
| L. He et al. [18] | N = 170 (study with control group) SAMPLE: serum | ↑ miRNA-144-3p | GATA3, STAT6, SOCS5, RHOA, NR3C1 i PTEN | Increased eosinophilia, inflammatory process, and remodeling of the airways. |
| A. T. Kho et al. [2] | N = 153 (study with control group) STUDY GROUP: ASTHMA EXACERBATION Age: 8.9 ± 2.2 years CONTROL GROUP: NO EXACERBATION Age: 8.9 ± 2.0 years SAMPLE: serum | ↑ miRNA-146b-5p ↑ miRNA-206 ↑ miRNA-720 | - | Upregulation GSK3 by AKT and downregulation NF-kB pathway. Remodeling the airways and deepening the inflammatory process. |
| A. Tiwari et al. [24] | N = 351 (study with control group) STUDY GROUP: ASTHMA EXACERBATION Age: 9.0 ± 1.9 years CONTROL GROUP: NO EXACERBATION Age: 9.4 ± 1.8 years SAMPLE: serum | ↓ miRNA-451b ↓ miRNA-7-5p ↑ miRNA-532-3p ↑ miRNA-296-5p ↑ miRNA-766-3p) | Many genes involved in the production of more than 20 proteins. | Upregulation MAPK, PI3K-Akt (i.a). Increased eosinophils, neutrophils, IgE immunoglobulin, and the activation of tumor necrosis factor alpha (TNF-a). Excessive production of interleukins and cytokines and intensification of inflammation. |
| A. A. Ibrahim et al. [25] | N = 100 (study with control group) STUDY GROUP: MILD, MODERATE, SEVERE ASTHMA Age: 8.9 ± 1.3 years CONTROL GROUP Age: 8.2 ± 1.4 years SAMPLE: serum | ↑ miRNA-196-2a | ANXA1 | Downregukation Annexin (anti-inflammatory factor). Concentration: moderate asthma > severe asthma. Increased inflammatory reaction. |
| Q. Liu et al. [31] | N = 360 (study with control group) STUDY GROUP: ASTHMA Age: 10.8 ± 3.1 years CONTROL GROUP Age 10.1 ± 2.7 years SAMPLE: serum | ↑ miRNA-155 | - | Responsible for the enhancement of the type 2 immune response. |
| A. T. Kho et al. [35] | N = 360 (study without control group) Age: 8.8 ± 2.1 years SAMPLE: serum | ↑ miRNA-186-5p ↑ miRNA-203 ↑ miRNA-26 | ≈50 | Activation of cholinergic pathway, intensifies the inflammatory process in the airways leading to their obstruction. Hypertrophy of smooth muscles. |
| A. Tiwari et al. [33] | N = 398 (study without control group) Age: 5–12 years SAMPLE: serum | ↑ let-7d-3p ↑ miRNA-328-3p | - | Decreased serum level of IL-13. Exacerbation of symptoms in patients allergic to aspergilus. |
| F. C. Mendes et al. [39] | N = 186 (study with control group) STUDY GROUP: ASTHMA Age: 8.7 ± 0.8 years CONTROL GROUP Age: 8.7 ± 0.8 years SAMPLE: exhaled breath condensate | ↑ miRNA -155 ↑ miR-126-3p ↑ miR-133a-3p ↑ miR-145-5p, ↑ miRNA-423-3p | E.G RUNX3 | GATA-3-upregulation of Th1/Th2 balance. Downregulation of GATA-3. Promotes a lymphocyte Th2 response. Increase in the levels of the IL-13 inflammatory response. |
| F. C. Mendes et al. [40] | N = 150 (study with control group) Age: 7–12 years SAMPLE: exhaled breath condensate | ↑ miR-133a-3p | - | Upregulation of production of IL-13. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kierbiedź-Guzik, N.; Sozańska, B. The Potential Role of Serum and Exhaled Breath Condensate miRNAs in Diagnosis and Predicting Exacerbations in Pediatric Asthma. Biomedicines 2023, 11, 763. https://doi.org/10.3390/biomedicines11030763
Kierbiedź-Guzik N, Sozańska B. The Potential Role of Serum and Exhaled Breath Condensate miRNAs in Diagnosis and Predicting Exacerbations in Pediatric Asthma. Biomedicines. 2023; 11(3):763. https://doi.org/10.3390/biomedicines11030763
Chicago/Turabian StyleKierbiedź-Guzik, Natalia, and Barbara Sozańska. 2023. "The Potential Role of Serum and Exhaled Breath Condensate miRNAs in Diagnosis and Predicting Exacerbations in Pediatric Asthma" Biomedicines 11, no. 3: 763. https://doi.org/10.3390/biomedicines11030763