
The Proteomes of Oral Cells Change during Co-Cultivation with Aggregatibacter actinomycetemcomitans and Eikenella corrodens

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Characteristic Phenotypes of Oral Cells
3.2. Tissue-Specific Characteristics
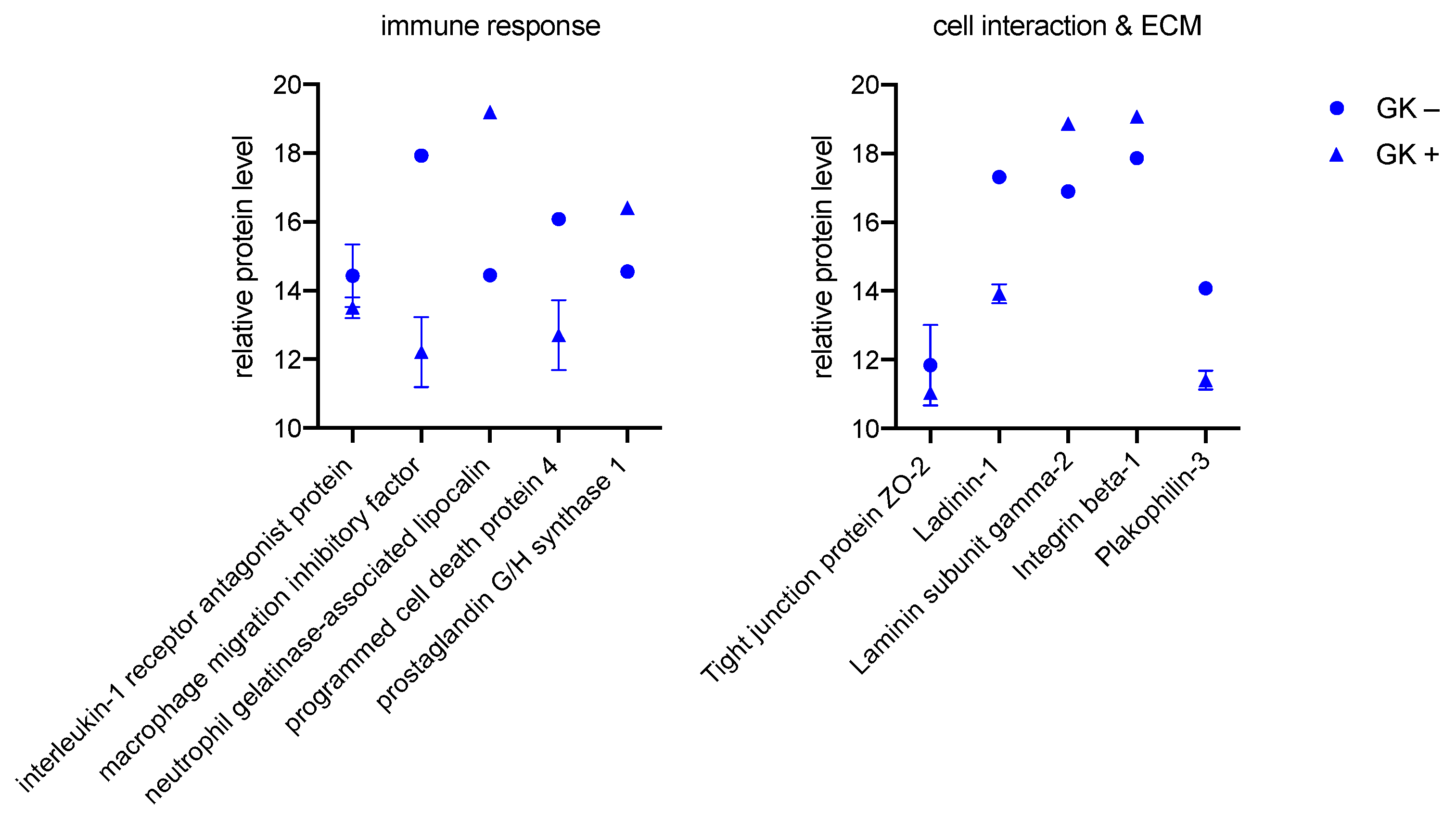
3.2.1. GK
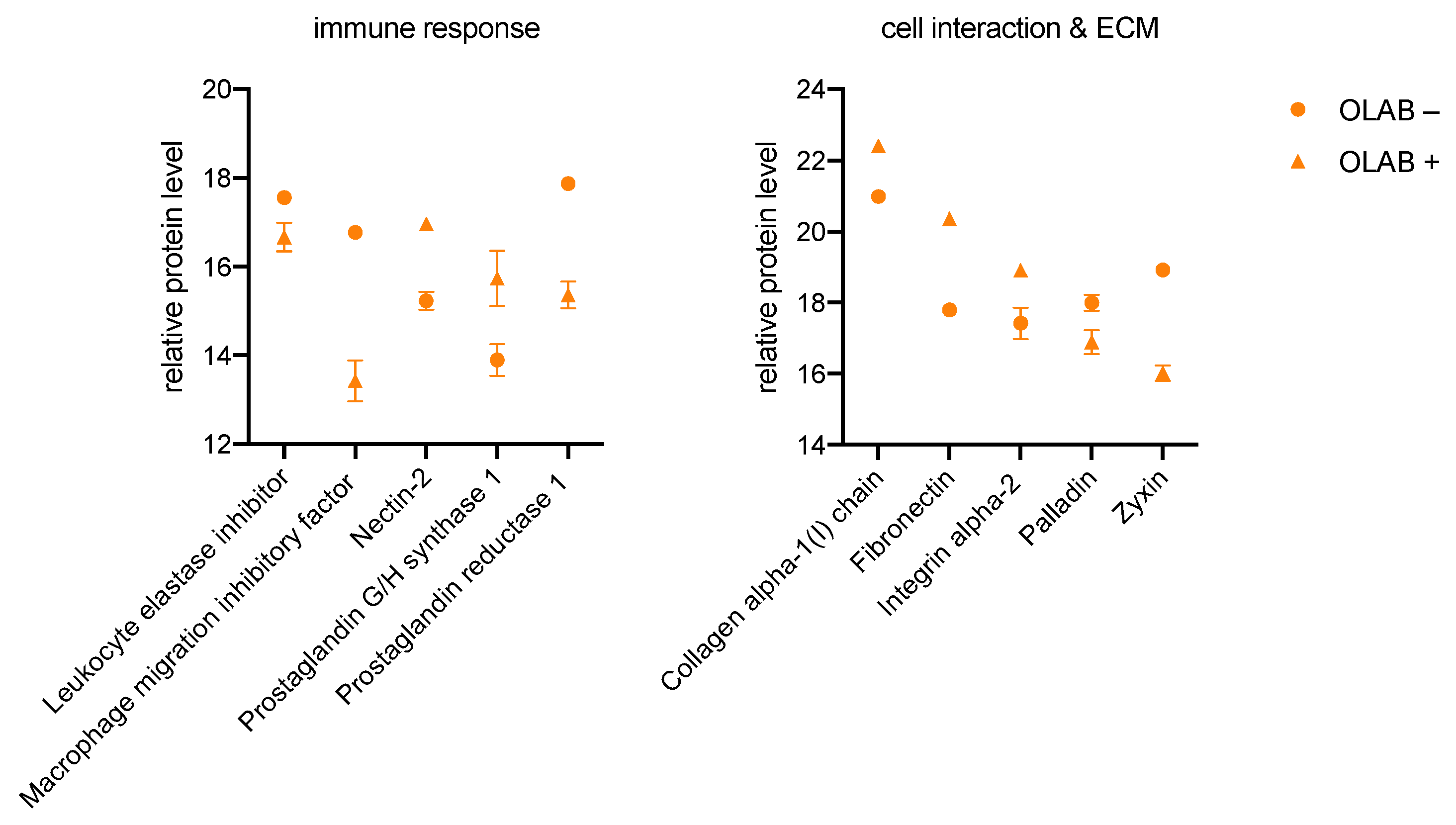
3.2.2. OBLAs
3.2.3. PDLF
3.2.4. CC
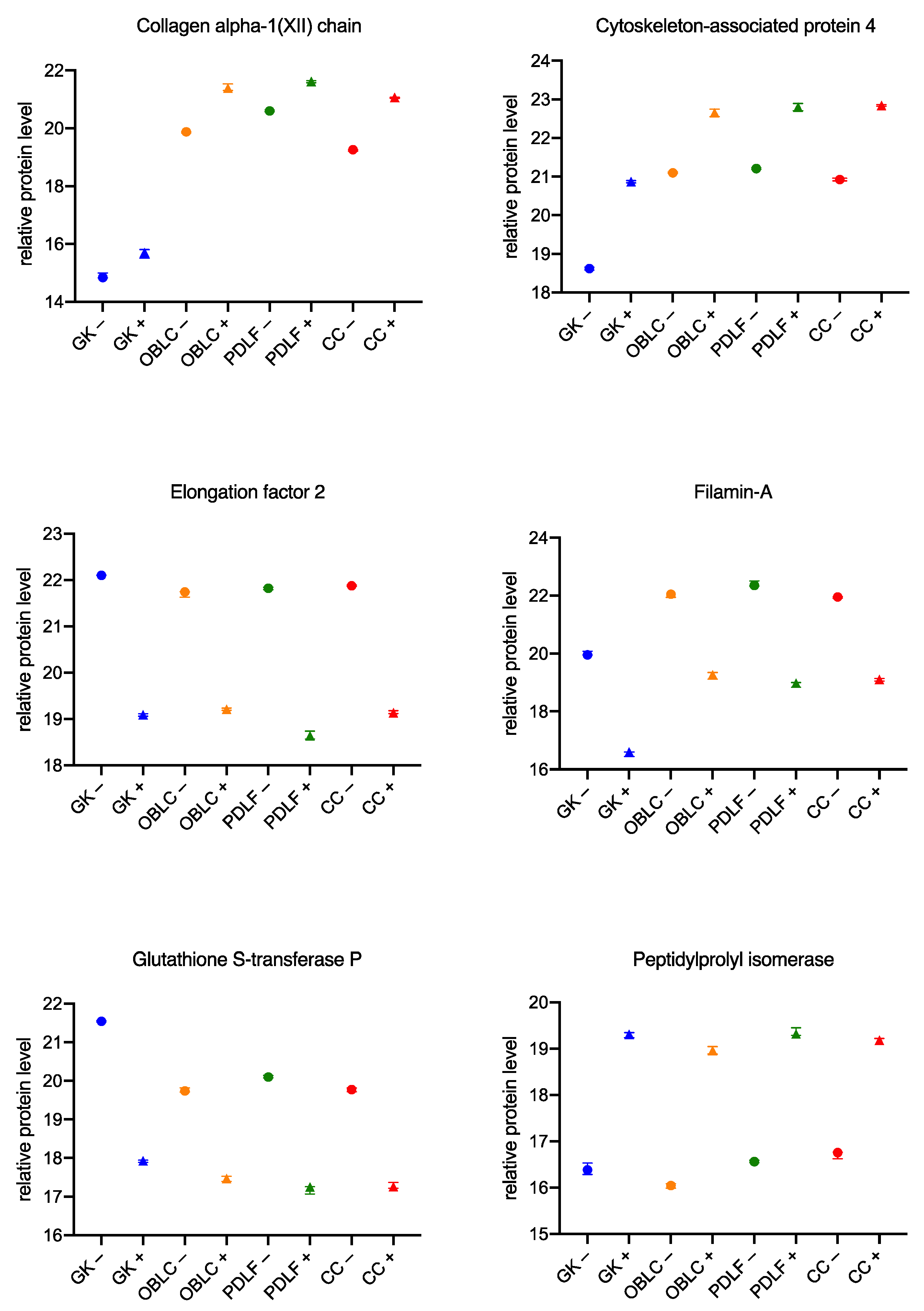
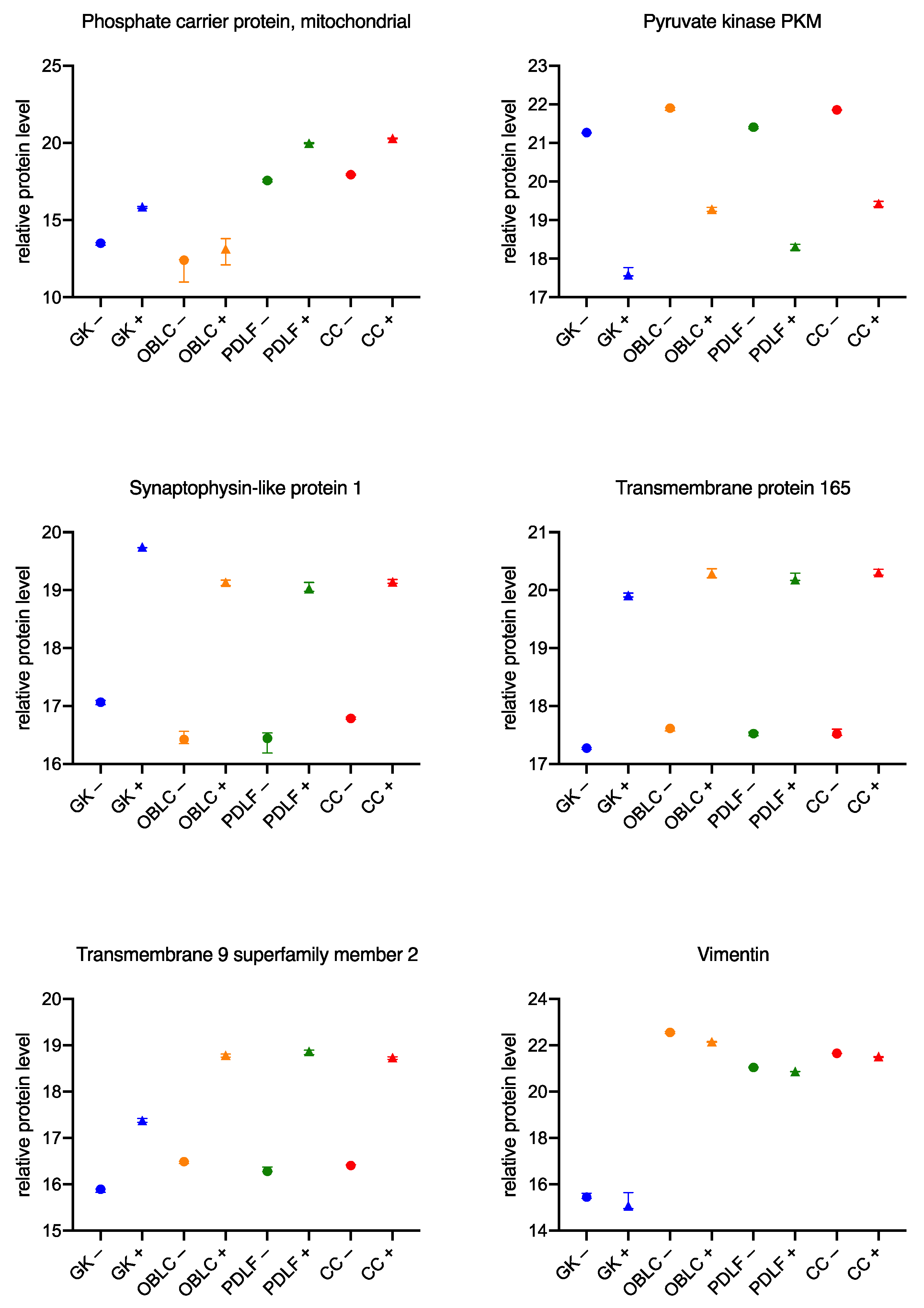
3.3. Similar Changes in the Proteome of the Cell Lines during Culture with the Bacteria
4. Discussion
4.1. The Proteome of Mesodermal Lineage Cells Vary Distinctly from Those of Ectodermal Lineage
4.2. Changes in Keratin Pattern
4.3. Modulation of an Enhanced Immune Response in All Investigated Cells
4.4. Proteins of Cell Interaction and ECM Are Affected by Culture with Bacteria
4.5. Similar Changes in the Proteome across Cell Lines
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Narayanan, A.S.; Bartold, P.M. Biochemistry of Periodontal Connective Tissues and Their Regeneration: A Current Perspective. Connect. Tissue Res. 1996, 34, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Berkovitz, B.K. The Structure of the Periodontal Ligament: An Update. Eur. J. Orthod. 1990, 12, 51–76. [Google Scholar] [CrossRef]
- Tatakis, D.N.; Kumar, P.S. Etiology and Pathogenesis of Periodontal Diseases. Dent. Clin. N. Am. 2005, 49, 491–516. [Google Scholar] [CrossRef] [PubMed]
- Bonewald, L.F. The Amazing Osteocyte. J. Bone Miner. Res. 2011, 26, 229–238. [Google Scholar] [CrossRef]
- Long, F. Building Strong Bones: Molecular Regulation of the Osteoblast Lineage. Nat. Rev. Mol. Cell Biol. 2011, 13, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Birkedal-Hansen, H.; Butler, W.T.; Taylor, R.E. Proteins of the Periodontium: Characterization of the Insoluble Collagens of Bovine Dental Cementum. Calcif. Tissue Res. 1977, 23, 39–44. [Google Scholar] [CrossRef]
- Butler, W.T.; Birkedal-Hansen, H.; Beegle, W.F.; Taylor, R.E.; Chung, E. Proteins of the Periodontium. Identification of Collagens with the [Alpha1(I)]2alpha2 and [Alpha1(III)]3 Structures in Bovine Periodontal Ligament. J. Biol. Chem. 1975, 250, 8907–8912. [Google Scholar] [CrossRef]
- Fullmer, H.M.; Lillie, R.D. The Oxytalan Fiber: A Previously Undescribed Connective Tissue Fiber. J. Histochem. Cytochem. 1958, 6, 425–430. [Google Scholar] [CrossRef]
- Schubert, A.; Schminke, B.; Miosge, N. Fibulins and Matrilins Are Novel Structural Components of the Periodontium in the Mouse. Arch. Oral Biol. 2017, 82, 216–222. [Google Scholar] [CrossRef]
- Lindberg, K.; Rheinwald, J.G. Three Distinct Keratinocyte Subtypes Identified in Human Oral Epithelium by Their Patterns of Keratin Expression in Culture and in Xenografts. Differentiation 1990, 45, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Bartold, P.M.; Wiebkin, O.W.; Thonard, J.C. Glycosaminoglycans of Human Gingival Epithelium and Connective Tissue. Connect. Tissue Res. 1981, 9, 99–106. [Google Scholar] [CrossRef]
- Chen, C.K.; Wilson, M.E. Eikenella Corrodens in Human Oral and Non-Oral Infections: A Review. J. Periodontol. 1992, 63, 941–953. [Google Scholar] [CrossRef]
- Thornton-Evans, G.; Eke, P.; Wei, L.; Palmer, A.; Moeti, R.; Hutchins, S.; Borrell, L.N.; Centers for Disease Control and Prevention (CDC). Periodontitis among Adults Aged ≥30 Years—United States, 2009–2010. MMWR Suppl. 2013, 62, 129–135. [Google Scholar]
- Kilian, M.; Frandsen, E.V.G.; Haubek, D.; Poulsen, K. The Etiology of Periodontal Disease Revisited by Population Genetic Analysis. Periodontology 2000 2006, 42, 158–179. [Google Scholar] [CrossRef]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal Diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.A.; Krauss, J. Neutrophils in Periodontal Inflammation. Front. Oral Biol. 2012, 15, 56–83. [Google Scholar] [CrossRef] [Green Version]
- Yucel-Lindberg, T.; Båge, T. Inflammatory Mediators in the Pathogenesis of Periodontitis. Expert Rev. Mol. Med. 2013, 15, e7. [Google Scholar] [CrossRef] [Green Version]
- Rylev, M.; Kilian, M. Prevalence and Distribution of Principal Periodontal Pathogens Worldwide. J. Clin. Periodontol. 2008, 35, 346–361. [Google Scholar] [CrossRef]
- Frandsen, E.V.; Poulsen, K.; Curtis, M.A.; Kilian, M. Evidence of Recombination in Porphyromonas Gingivalis and Random Distribution of Putative Virulence Markers. Infect. Immun. 2001, 69, 4479–4485. [Google Scholar] [CrossRef] [Green Version]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L. Microbial Complexes in Subgingival Plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef]
- Haffajee, A.D.; Socransky, S.S. Microbiology of Periodontal Diseases: Introduction. Periodontol. 2000 2005, 38, 9–12. [Google Scholar] [CrossRef]
- Balaji, T.M.; Varadarajan, S.; Jagannathan, R.; Mahendra, J.; Fageeh, H.I.; Fageeh, H.N.; Mushtaq, S.; Baeshen, H.A.; Bhandi, S.; Gupta, A.A.; et al. Melatonin as a Topical/Systemic Formulation for the Management of Periodontitis: A Systematic Review. Materials 2021, 14, 2417. [Google Scholar] [CrossRef]
- Guarnieri, R.; Zanza, A.; D’Angelo, M.; Di Nardo, D.; Del Giudice, A.; Mazzoni, A.; Reda, R.; Testarelli, L. Correlation between Peri-Implant Marginal Bone Loss Progression and Peri-Implant Sulcular Fluid Levels of Metalloproteinase-8. J. Pers. Med. 2022, 12, 58. [Google Scholar] [CrossRef]
- Konig, M.F.; Abusleme, L.; Reinholdt, J.; Palmer, R.J.; Teles, R.P.; Sampson, K.; Rosen, A.; Nigrovic, P.A.; Sokolove, J.; Giles, J.T.; et al. Aggregatibacter Actinomycetemcomitans-Induced Hypercitrullination Links Periodontal Infection to Autoimmunity in Rheumatoid Arthritis. Sci. Transl. Med. 2016, 8, 369ra176. [Google Scholar] [CrossRef] [Green Version]
- Sima, C.; Glogauer, M. Diabetes Mellitus and Periodontal Diseases. Curr. Diabetes Rep. 2013, 13, 445–452. [Google Scholar] [CrossRef]
- Lockhart, P.B.; Bolger, A.F.; Papapanou, P.N.; Osinbowale, O.; Trevisan, M.; Levison, M.E.; Taubert, K.A.; Newburger, J.W.; Gornik, H.L.; Gewitz, M.H.; et al. Periodontal Disease and Atherosclerotic Vascular Disease: Does the Evidence Support an Independent Association?: A Scientific Statement from the American Heart Association. Circulation 2012, 125, 2520–2544. [Google Scholar] [CrossRef]
- Schminke, B.; Vom Orde, F.; Gruber, R.; Schliephake, H.; Bürgers, R.; Miosge, N. The Pathology of Bone Tissue during Peri-Implantitis. J. Dent. Res. 2015, 94, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Meyer, D.H.; Lippmann, J.E.; Fives-Taylor, P.M. Invasion of Epithelial Cells by Actinobacillus Actinomycetemcomitans: A Dynamic, Multistep Process. Infect. Immun. 1996, 64, 2988–2997. [Google Scholar] [CrossRef] [Green Version]
- Zapata, A.; Ramirez-Arcos, S. A Comparative Study of McFarland Turbidity Standards and the Densimat Photometer to Determine Bacterial Cell Density. Curr. Microbiol. 2015, 70, 907–909. [Google Scholar] [CrossRef]
- Atanassov, I.; Urlaub, H. Increased Proteome Coverage by Combining PAGE and Peptide Isoelectric Focusing: Comparative Study of Gel-Based Separation Approaches. Proteomics 2013, 13, 2947–2955. [Google Scholar] [CrossRef]
- Zhang, Y.; Bilbao, A.; Bruderer, T.; Luban, J.; Strambio-De-Castillia, C.; Lisacek, F.; Hopfgartner, G.; Varesio, E. The Use of Variable Q1 Isolation Windows Improves Selectivity in LC-SWATH-MS Acquisition. J. Proteome Res. 2015, 14, 4359–4371. [Google Scholar] [CrossRef]
- Lambert, J.-P.; Ivosev, G.; Couzens, A.L.; Larsen, B.; Taipale, M.; Lin, Z.-Y.; Zhong, Q.; Lindquist, S.; Vidal, M.; Aebersold, R.; et al. Mapping Differential Interactomes by Affinity Purification Coupled with Data-Independent Mass Spectrometry Acquisition. Nat. Methods 2013, 10, 1239–1245. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus Computational Platform for Comprehensive Analysis of (Prote)Omics Data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Vizcaíno, J.A.; Deutsch, E.W.; Wang, R.; Csordas, A.; Reisinger, F.; Ríos, D.; Dianes, J.A.; Sun, Z.; Farrah, T.; Bandeira, N.; et al. ProteomeXchange Provides Globally Coordinated Proteomics Data Submission and Dissemination. Nat. Biotechnol. 2014, 32, 223–226. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Doss, M.X.; Gaspar, J.A.; Winkler, J.; Hescheler, J.; Schulz, H.; Sachinidis, A. Specific Gene Signatures and Pathways in Mesodermal Cells and Their Derivatives Derived from Embryonic Stem Cells. Stem Cell Rev. Rep. 2012, 8, 43–54. [Google Scholar] [CrossRef]
- Weinreb, M.; Nemcovsky, C.E. In Vitro Models for Evaluation of Periodontal Wound Healing/Regeneration. Periodontology 2000 2015, 68, 41–54. [Google Scholar] [CrossRef]
- Argelaguet, R.; Clark, S.J.; Mohammed, H.; Stapel, L.C.; Krueger, C.; Kapourani, C.-A.; Imaz-Rosshandler, I.; Lohoff, T.; Xiang, Y.; Hanna, C.W.; et al. Multi-Omics Profiling of Mouse Gastrulation at Single-Cell Resolution. Nature 2019, 576, 487–491. [Google Scholar] [CrossRef]
- Li, S.; Dong, G.; Moschidis, A.; Ortiz, J.; Benakanakere, M.R.; Kinane, D.F.; Graves, D.T.P. Gingivalis Modulates Keratinocytes through FOXO Transcription Factors. PLoS ONE 2013, 8, e78541. [Google Scholar] [CrossRef]
- Huynh, A.H.S.; Veith, P.D.; McGregor, N.R.; Adams, G.G.; Chen, D.; Reynolds, E.C.; Ngo, L.H.; Darby, I.B. Gingival Crevicular Fluid Proteomes in Health, Gingivitis and Chronic Periodontitis. J. Periodontal. Res. 2015, 50, 637–649. [Google Scholar] [CrossRef]
- Sirniö, P.; Väyrynen, J.P.; Mutt, S.J.; Herzig, K.-H.; Walkowiak, J.; Klintrup, K.; Mäkelä, J.; Karttunen, T.J.; Mäkinen, M.J.; Tuomisto, A. Systemic Inflammation Is Associated with Circulating Cell Death Released Keratin 18 Fragments in Colorectal Cancer. Oncoimmunology 2020, 9, 1783046. [Google Scholar] [CrossRef]
- Westerlund, U.; Ingman, T.; Lukinmaa, P.L.; Salo, T.; Kjeldsen, L.; Borregaard, N.; Tjäderhane, L.; Konttinen, Y.T.; Sorsa, T. Human Neutrophil Gelatinase and Associated Lipocalin in Adult and Localized Juvenile Periodontitis. J. Dent. Res. 1996, 75, 1553–1563. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Arend, W.P.; Malyak, M.; Guthridge, C.J.; Gabay, C. Interleukin-1 Receptor Antagonist: Role in Biology. Annu. Rev. Immunol. 1998, 16, 27–55. [Google Scholar] [CrossRef]
- Ortiz-García, Y.M.; García-Iglesias, T.; Morales-Velazquez, G.; Lazalde-Ramos, B.P.; Zúñiga-González, G.M.; Ortiz-García, R.G.; Zamora-Perez, A.L. Macrophage Migration Inhibitory Factor Levels in Gingival Crevicular Fluid, Saliva, and Serum of Chronic Periodontitis Patients. BioMed Res. Int. 2019, 2019, 7850392. [Google Scholar] [CrossRef] [Green Version]
- Simmons, D.L.; Botting, R.M.; Hla, T. Cyclooxygenase Isozymes: The Biology of Prostaglandin Synthesis and Inhibition. Pharmacol. Rev. 2004, 56, 387–437. [Google Scholar] [CrossRef] [Green Version]
- Yokomizo, T.; Ogawa, Y.; Uozumi, N.; Kume, K.; Izumi, T.; Shimizu, T. CDNA Cloning, Expression, and Mutagenesis Study of Leukotriene B4 12-Hydroxydehydrogenase. J. Biol. Chem. 1996, 271, 2844–2850. [Google Scholar] [CrossRef] [Green Version]
- Torriglia, A.; Martin, E.; Jaadane, I. The Hidden Side of SERPINB1/Leukocyte Elastase Inhibitor. Semin. Cell Dev. Biol. 2017, 62, 178–186. [Google Scholar] [CrossRef]
- Hassan, N.J.; Barclay, A.N.; Brown, M.H. Frontline: Optimal T Cell Activation Requires the Engagement of CD6 and CD166. Eur. J. Immunol. 2004, 34, 930–940. [Google Scholar] [CrossRef]
- Deuss, F.A.; Gully, B.S.; Rossjohn, J.; Berry, R. Recognition of Nectin-2 by the Natural Killer Cell Receptor T Cell Immunoglobulin and ITIM Domain (TIGIT). J. Biol. Chem. 2017, 292, 11413–11422. [Google Scholar] [CrossRef] [Green Version]
- Chapoval, A.I.; Ni, J.; Lau, J.S.; Wilcox, R.A.; Flies, D.B.; Liu, D.; Dong, H.; Sica, G.L.; Zhu, G.; Tamada, K.; et al. B7-H3: A Costimulatory Molecule for T Cell Activation and IFN-Gamma Production. Nat. Immunol. 2001, 2, 269–274. [Google Scholar] [CrossRef]
- Courts, F.J.; Boackle, R.J.; Fudenberg, H.H.; Silverman, M.S. Detection of Functional Complement Components in Gingival Crevicular Fluid from Humans with Periodontal Diseases. J. Dent. Res. 1977, 56, 327–331. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms Regulating Intestinal Barrier Integrity and Its Pathological Implications. Exp. Mol. Med. 2018, 50, 103. [Google Scholar] [CrossRef] [Green Version]
- Abé, T.; Maruyama, S.; Yamazaki, M.; Xu, B.; Babkair, H.; Sumita, Y.; Cheng, J.; Yamamoto, T.; Saku, T. Proteomic and Histopathological Characterization of the Interface between Oral Squamous Cell Carcinoma Invasion Fronts and Non-Cancerous Epithelia. Exp. Mol. Pathol. 2017, 102, 327–336. [Google Scholar] [CrossRef]
- Bonné, S.; Gilbert, B.; Hatzfeld, M.; Chen, X.; Green, K.J.; van Roy, F. Defining Desmosomal Plakophilin-3 Interactions. J. Cell Biol. 2003, 161, 403–416. [Google Scholar] [CrossRef]
- Mykkänen, O.M.; Grönholm, M.; Rönty, M.; Lalowski, M.; Salmikangas, P.; Suila, H.; Carpén, O. Characterization of Human Palladin, a Microfilament-Associated Protein. Mol. Biol. Cell 2001, 12, 3060–3073. [Google Scholar] [CrossRef] [Green Version]
- Frenette, P.S.; Wagner, D.D. Adhesion Molecules—Part 1. N. Engl. J. Med. 1996, 334, 1526–1529. [Google Scholar] [CrossRef]
- Fingleton, B. Matrix Metalloproteinases as Regulators of Inflammatory Processes. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2036–2042. [Google Scholar] [CrossRef]
- Timpl, R.; Brown, J.C. Supramolecular Assembly of Basement Membranes. Bioessays 1996, 18, 123–132. [Google Scholar] [CrossRef]
- Larjava, H.; Koivisto, L.; Heino, J.; Häkkinen, L. Integrins in Periodontal Disease. Exp. Cell Res. 2014, 325, 104–110. [Google Scholar] [CrossRef]
- Pankov, R.; Yamada, K.M. Fibronectin at a Glance. J. Cell Sci. 2002, 115, 3861–3863. [Google Scholar] [CrossRef] [Green Version]
- Govindaraju, P.; Todd, L.; Shetye, S.; Monslow, J.; Puré, E. CD44-Dependent Inflammation, Fibrogenesis, and Collagenolysis Regulates Extracellular Matrix Remodeling and Tensile Strength during Cutaneous Wound Healing. Matrix Biol. 2019, 75–76, 314–330. [Google Scholar] [CrossRef]
- Lu, H.; Zhou, Q.; He, J.; Jiang, Z.; Peng, C.; Tong, R.; Shi, J. Recent Advances in the Development of Protein-Protein Interactions Modulators: Mechanisms and Clinical Trials. Signal Transduct. Target Ther. 2020, 5, 213. [Google Scholar] [CrossRef]
- Reichenberger, E.; Baur, S.; Sukotjo, C.; Olsen, B.R.; Karimbux, N.Y.; Nishimura, I. Collagen XII Mutation Disrupts Matrix Structure of Periodontal Ligament and Skin. J. Dent. Res. 2000, 79, 1962–1968. [Google Scholar] [CrossRef]
- Borregaard, N.; Sørensen, O.E.; Theilgaard-Mönch, K. Neutrophil Granules: A Library of Innate Immunity Proteins. Trends Immunol. 2007, 28, 340–345. [Google Scholar] [CrossRef]
- Castello, A.; Fischer, B.; Eichelbaum, K.; Horos, R.; Beckmann, B.M.; Strein, C.; Davey, N.E.; Humphreys, D.T.; Preiss, T.; Steinmetz, L.M.; et al. Insights into RNA Biology from an Atlas of Mammalian MRNA-Binding Proteins. Cell 2012, 149, 1393–1406. [Google Scholar] [CrossRef] [Green Version]
- Savoy, R.M.; Ghosh, P.M. The Dual Role of Filamin A in Cancer: Can’t Live with (Too Much of) It, Can’t Live without It. Endocr. Relat. Cancer 2013, 20, R341–R356. [Google Scholar] [CrossRef] [Green Version]
- Bogaards, J.J.; Venekamp, J.C.; van Bladeren, P.J. Stereoselective Conjugation of Prostaglandin A2 and Prostaglandin J2 with Glutathione, Catalyzed by the Human Glutathione S-Transferases A1-1, A2-2, M1a-1a, and P1-1. Chem. Res. Toxicol. 1997, 10, 310–317. [Google Scholar] [CrossRef]
- Camargo Ortega, V.R.; Bravo López, L.D.; Visoso Salgado, A.; Mejia Sanchez, F.; Castillo Cadena, J. Polymorphisms in Glutathione S-Transferase M1, T1, and P1 in Patients with Chronic Periodontitis: A Pilot Study. Int. Sch. Res. Notices 2014, 2014, 135368. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A.; Horie, T.; Numabe, Y. Investigation of Molecular Biomarker Candidates for Diagnosis and Prognosis of Chronic Periodontitis by Bioinformatics Analysis of Pooled Microarray Gene Expression Datasets in Gene Expression Omnibus (GEO). BMC Oral Health 2019, 19, 52. [Google Scholar] [CrossRef] [Green Version]
- Mayr, J.A.; Merkel, O.; Kohlwein, S.D.; Gebhardt, B.R.; Böhles, H.; Fötschl, U.; Koch, J.; Jaksch, M.; Lochmüller, H.; Horváth, R.; et al. Mitochondrial Phosphate-Carrier Deficiency: A Novel Disorder of Oxidative Phosphorylation. Am. J. Hum. Genet. 2007, 80, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Kurihara-Shimomura, M.; Sasahira, T.; Nakashima, C.; Kuniyasu, H.; Shimomura, H.; Kirita, T. The Multifarious Functions of Pyruvate Kinase M2 in Oral Cancer Cells. Int. J. Mol. Sci. 2018, 19, 2907. [Google Scholar] [CrossRef] [Green Version]
- Craig, T.J.; Anderson, D.; Evans, A.J.; Girach, F.; Henley, J.M. SUMOylation of Syntaxin1A Regulates Presynaptic Endocytosis. Sci. Rep. 2015, 5, 17669. [Google Scholar] [CrossRef] [Green Version]
- Offenbacher, S.; Divaris, K.; Barros, S.P.; Moss, K.L.; Marchesan, J.T.; Morelli, T.; Zhang, S.; Kim, S.; Sun, L.; Beck, J.D.; et al. Genome-Wide Association Study of Biologically Informed Periodontal Complex Traits Offers Novel Insights into the Genetic Basis of Periodontal Disease. Hum. Mol. Genet. 2016, 25, 2113–2129. [Google Scholar] [CrossRef] [Green Version]
- Clark, C.R.; Maile, M.; Blaney, P.; Hellweg, S.R.; Strauss, A.; Durose, W.; Priya, S.; Habicht, J.; Burns, M.B.; Blekhman, R.; et al. Transposon Mutagenesis Screen in Mice Identifies TM9SF2 as a Novel Colorectal Cancer Oncogene. Sci. Rep. 2018, 8, 15327. [Google Scholar] [CrossRef]
- Challa, A.A.; Stefanovic, B. A Novel Role of Vimentin Filaments: Binding and Stabilization of Collagen MRNAs. Mol. Cell. Biol. 2011, 31, 3773–3789. [Google Scholar] [CrossRef] [Green Version]
- Satelli, A.; Li, S. Vimentin in Cancer and Its Potential as a Molecular Target for Cancer Therapy. Cell. Mol. Life Sci. 2011, 68, 3033–3046. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schminke, B.; Kauffmann, P.; Brockmeyer, P.; Miosge, N.; Lenz, C.; Schubert, A. The Proteomes of Oral Cells Change during Co-Cultivation with Aggregatibacter actinomycetemcomitans and Eikenella corrodens. Biomedicines 2023, 11, 700. https://doi.org/10.3390/biomedicines11030700
Schminke B, Kauffmann P, Brockmeyer P, Miosge N, Lenz C, Schubert A. The Proteomes of Oral Cells Change during Co-Cultivation with Aggregatibacter actinomycetemcomitans and Eikenella corrodens. Biomedicines. 2023; 11(3):700. https://doi.org/10.3390/biomedicines11030700
Chicago/Turabian StyleSchminke, Boris, Philipp Kauffmann, Phillipp Brockmeyer, Nicolai Miosge, Christof Lenz, and Andrea Schubert. 2023. "The Proteomes of Oral Cells Change during Co-Cultivation with Aggregatibacter actinomycetemcomitans and Eikenella corrodens" Biomedicines 11, no. 3: 700. https://doi.org/10.3390/biomedicines11030700