

Secretome of hESC-Derived MSC-like Immune and Matrix Regulatory Cells Mitigate Pulmonary Fibrosis through Antioxidant and Anti-Inflammatory Effects
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Secretome Containing hESC-MSC-IMRC Culture Medium (hESC-MSC-IMRC-CM)
2.2. Animal Welfare Statement and Procedures
2.3. Cell Culture and BLM-Induced A549 Alveolar Epithelial Cells Injury
2.4. Lung Index
2.5. Lung Histopathological and Immunohistochemical Analysis
2.6. Hydroxyproline Content in Mice Lung Tissues
2.7. Immunoblotting Analysis
2.8. Measurement of Oxidative Stress and ROS Staining
2.9. ELISA for TNF-α, IL-1β, IL-6
2.10. Statistical Analysis
3. Results
3.1. An Elevated Oxidative Stress and Tlr4/MyD88 Signaling Activity in Lungs of BLM-Induced PF Mice
3.2. BLM Induces Oxidative Stress and NOX4-Mediated ROS Production in Lung Epithelial Cells
3.3. hESC-MSC-IMRC-CM Treatment Suppresses the BLM-Induced Oxidative Stress in Lung Epithelial Cells
3.4. hESC-MSC-IMRCs and hESC-MSC-IMRC-CM Mitigate BLM-Induced PF in Mice
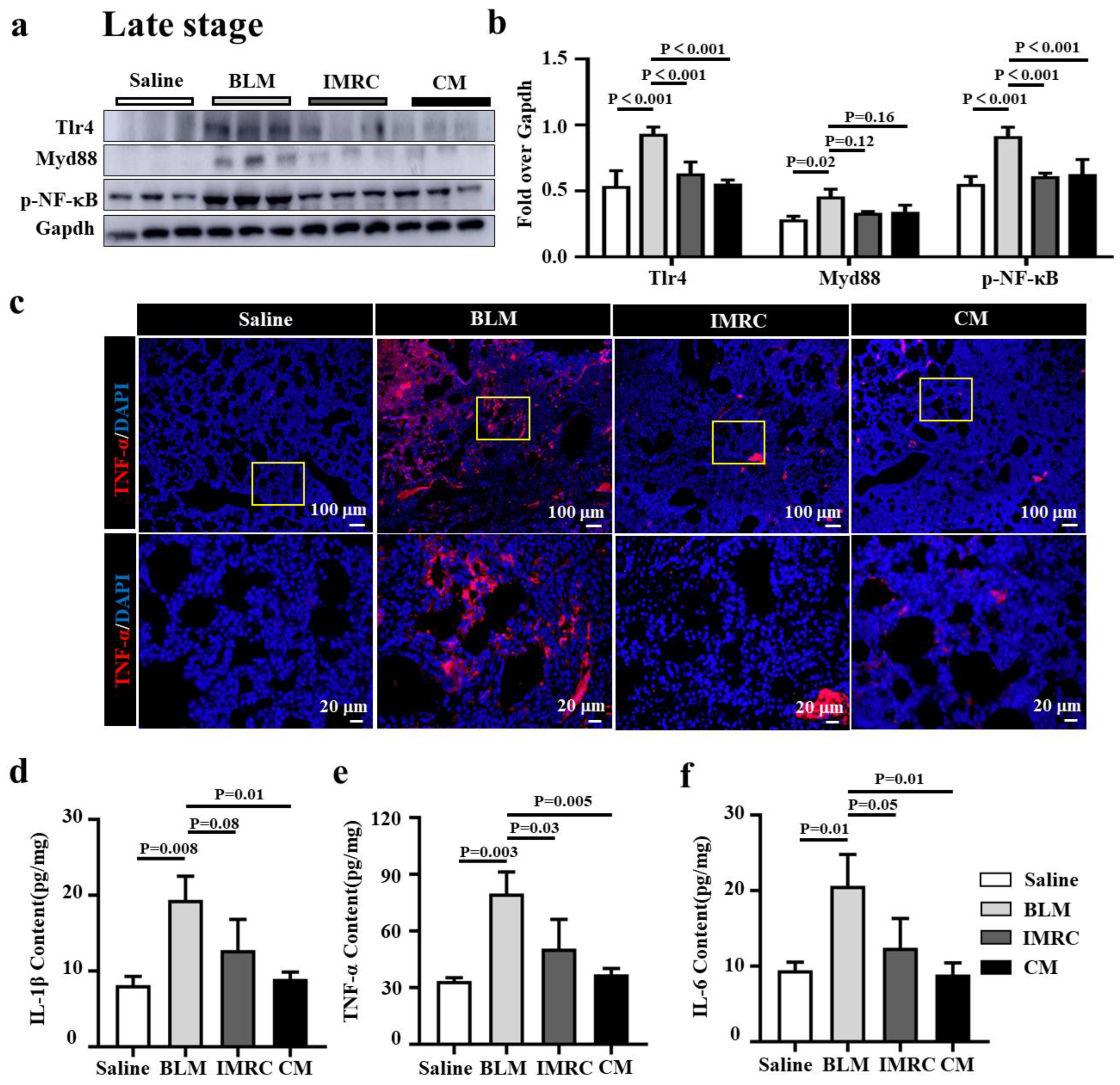
3.5. hESC-MSC-IMRCs or hESC-MSC-IMRC-CM Alleviates Lung Inflammation in PF Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raghu, G. Idiopathic pulmonary fibrosis: Guidelines for diagnosis and clinical management have advanced from consensus-based in 2000 to evidence-based in 2011. Eur. Respir. J. 2011, 37, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Fois, A.G.; Paliogiannis, P.; Sotgia, S.; Mangoni, A.A.; Zinellu, E.; Pirina, P.; Carru, C.; Zinellu, A. Evaluation of oxidative stress biomarkers in idiopathic pulmonary fibrosis and therapeutic applications: A systematic review. Respir. Res. 2018, 19, 51. [Google Scholar] [CrossRef] [PubMed]
- Desai, O.; Winkler, J.; Minasyan, M.; Herzog, E.L. The Role of Immune and Inflammatory Cells in Idiopathic Pulmonary Fibrosis. Front. Med. (Lausanne) 2018, 5, 43. [Google Scholar] [CrossRef]
- Liu, R.M.; Liu, G. Cell senescence and fibrotic lung diseases. Exp. Gerontol. 2020, 132, 110836. [Google Scholar] [CrossRef] [PubMed]
- Bolourani, S.; Brenner, M.; Wang, P. The interplay of DAMPs, TLR4, and proinflammatory cytokines in pulmonary fibrosis. J. Mol. Med. 2021, 99, 1373–1384. [Google Scholar] [CrossRef]
- Estornut, C.; Milara, J.; Bayarri, M.A.; Belhadj, N.; Cortijo, J. Targeting Oxidative Stress as a Therapeutic Approach for Idiopathic Pulmonary Fibrosis. Front. Pharmacol. 2022, 12, 3873. [Google Scholar] [CrossRef]
- Osborn-Heaford, H.L.; Ryan, A.J.; Murthy, S.; Racila, A.M.; He, C.; Sieren, J.C.; Spitz, D.R.; Carter, A.B. Mitochondrial Rac1 GTPase import and electron transfer from cytochrome c are required for pulmonary fibrosis. J. Biol. Chem. 2012, 287, 3301–3312. [Google Scholar] [CrossRef]
- Hecker, L.; Cheng, J.; Thannickal, V.J. Targeting NOX enzymes in pulmonary fibrosis. Cell. Mol. Life Sci. 2012, 69, 2365–2371. [Google Scholar] [CrossRef]
- Murthy, S.; Adamcakova-Dodd, A.; Perry, S.S.; Tephly, L.A.; Keller, R.M.; Metwali, N.; Meyerholz, D.K.; Wang, Y.; Glogauer, M.; Thorne, P.S.; et al. Modulation of reactive oxygen species by Rac1 or catalase prevents asbestos-induced pulmonary fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L846–L855. [Google Scholar] [CrossRef]
- Xia, C.; Dai, Z.; Jin, Y.; Chen, P. Emerging Antioxidant Paradigm of Mesenchymal Stem Cell-Derived Exosome Therapy. Front. Endocrinol. (Lausanne) 2021, 12, 727272. [Google Scholar] [CrossRef]
- Stavely, R.; Nurgali, K. The emerging antioxidant paradigm of mesenchymal stem cell therapy. Stem Cells Transl. Med. 2020, 9, 985–1006. [Google Scholar] [CrossRef] [PubMed]
- Gadkari, R.; Zhao, L.; Teklemariam, T.; Hantash, B.M. Human embryonic stem cell derived-mesenchymal stem cells: An alternative mesenchymal stem cell source for regenerative medicine therapy. Regen. Med. 2014, 9, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Billing, A.M.; Dib, S.S.; Bhagwat, A.M.; da Silva, I.T.; Drummond, R.D.; Hayat, S.; Al-Mismar, R.; Ben-Hamidane, H.; Goswami, N.; Engholm-Keller, K.; et al. A Systems-level Characterization of the Differentiation of Human Embryonic Stem Cells into Mesenchymal Stem Cells. Mol. Cell. Proteom. 2019, 18, 1950–1966. [Google Scholar] [CrossRef]
- Yang, S.; Liu, P.; Gao, T.; Song, D.; Zhao, X.; Li, Y.; Wu, J.; Wang, L.; Wang, Z.; Hao, J.; et al. Every road leads to Rome: Therapeutic effect and mechanism of the extracellular vesicles of human embryonic stem cell-derived immune and matrix regulatory cells administered to mouse models of pulmonary fibrosis through different routes. Stem Cell Res. Ther. 2022, 13, 163. [Google Scholar] [CrossRef]
- Bahrehbar, K.; Rezazadeh Valojerdi, M.; Esfandiari, F.; Fathi, R.; Hassani, S.N.; Baharvand, H. Human embryonic stem cell-derived mesenchymal stem cells improved premature ovarian failure. World J. Stem Cells 2020, 12, 857–878. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Song, D.; Li, Z.; Guo, B.; Xiao, Y.; Liu, W.; Liang, L.; Feng, C.; Gao, T.; Chen, Y.; et al. Immunity-and-matrix-regulatory cells derived from human embryonic stem cells safely and effectively treat mouse lung injury and fibrosis. Cell Res. 2020, 30, 794–809. [Google Scholar] [CrossRef]
- Ma, Z.J.; Yang, J.J.; Lu, Y.B.; Liu, Z.Y.; Wang, X.X. Mesenchymal stem cell-derived exosomes: Toward cell-free therapeutic strategies in regenerative medicine. World J. Stem Cells 2020, 12, 814–840. [Google Scholar] [CrossRef] [PubMed]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal Stem Cell Secretome: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef]
- Li, X.; An, G.; Wang, Y.; Liang, D.; Zhu, Z.; Tian, L. Targeted migration of bone marrow mesenchymal stem cells inhibits silica-induced pulmonary fibrosis in rats. Stem Cell Res. Ther. 2018, 9, 335. [Google Scholar] [CrossRef]
- Kawara, A.; Mizuta, R.; Fujisawa, M.; Ito, T.; Li, C.; Nakamura, K.; Sun, C.; Kuwabara, M.; Kitabatake, M.; Yoshimura, T.; et al. Spred2-deficiency enhances the proliferation of lung epithelial cells and alleviates pulmonary fibrosis induced by bleomycin. Sci. Rep. 2020, 10, 16490. [Google Scholar] [CrossRef]
- Kuroda, J.; Ago, T.; Matsushima, S.; Zhai, P.; Schneider, M.D.; Sadoshima, J. NADPH oxidase 4 (Nox4) is a major source of oxidative stress in the failing heart. Proc. Natl. Acad. Sci. USA 2010, 107, 15565–15570. [Google Scholar] [CrossRef]
- Li, Z.M.; Xu, S.Y.; Feng, Y.Z.; Cheng, Y.R.; Xiong, J.B.; Zhou, Y.; Guan, C.X. The role of NOX4 in pulmonary diseases. J. Cell. Physiol. 2021, 236, 1628–1637. [Google Scholar] [CrossRef] [PubMed]
- Harijith, A.; Basa, P.; Ha, A.; Thomas, J.; Jafri, A.; Fu, P.; MacFarlane, P.M.; Raffay, T.M.; Natarajan, V.; Sudhadevi, T. NOX4 Mediates Epithelial Cell Death in Hyperoxic Acute Lung Injury Through Mitochondrial Reactive Oxygen Species. Front. Pharmacol. 2022, 13, 880878. [Google Scholar] [CrossRef] [PubMed]
- Larson-Casey, J.L.; Gu, L.; Kang, J.; Dhyani, A.; Carter, A.B. NOX4 regulates macrophage apoptosis resistance to induce fibrotic progression. J. Biol. Chem. 2021, 297, 100810. [Google Scholar] [CrossRef]
- Jiang, J.; Huang, K.; Xu, S.; Garcia, J.G.N.; Wang, C.; Cai, H. Targeting NOX4 alleviates sepsis-induced acute lung injury via attenuation of redox-sensitive activation of CaMKII/ERK1/2/MLCK and endothelial cell barrier dysfunction. Redox Biol. 2020, 36, 101638. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, P.; Jiang, Y.; Wang, Z.; Dai, H.; Wang, C. Therapeutic Applications of Mesenchymal Stem Cells in Idiopathic Pulmonary Fibrosis. Front. Cell. Dev. Biol. 2021, 9, 639657. [Google Scholar] [CrossRef]
- Atanasova, E.; Milosevic, D.; Bornschlegl, S.; Krucker, K.P.; Jacob, E.K.; Carmona Porquera, E.M.; Anderson, D.K.; Egan, A.M.; Limper, A.H.; Dietz, A.B. Normal ex vivo mesenchymal stem cell function combined with abnormal immune profiles sets the stage for informative cell therapy trials in idiopathic pulmonary fibrosis patients. Stem Cell Res. Ther. 2022, 13, 45. [Google Scholar] [CrossRef]
- He, C.; Murthy, S.; McCormick, M.L.; Spitz, D.R.; Ryan, A.J.; Carter, A.B. Mitochondrial Cu,Zn-superoxide dismutase mediates pulmonary fibrosis by augmenting H2O2 generation. J. Biol. Chem. 2011, 286, 15597–15607. [Google Scholar] [CrossRef]
- Hecker, L.; Vittal, R.; Jones, T.; Jagirdar, R.; Luckhardt, T.R.; Horowitz, J.C.; Pennathur, S.; Martinez, F.J.; Thannickal, V.J. NADPH oxidase-4 mediates myofibroblast activation and fibrogenic responses to lung injury. Nat. Med. 2009, 15, 1077–1081. [Google Scholar] [CrossRef]
- Weinberg, F.; Hamanaka, R.; Wheaton, W.W.; Weinberg, S.; Joseph, J.; Lopez, M.; Kalyanaraman, B.; Mutlu, G.M.; Budinger, G.R.; Chandel, N.S. Mitochondrial metabolism and ROS generation are essential for Kras-mediated tumorigenicity. Proc. Natl. Acad. Sci. USA 2010, 107, 8788–8793. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, A.R.; Dos Santos, N.B.; Scavone, C.; Munhoz, C.D. Nrf2/ARE Pathway Modulation by Dietary Energy Regulation in Neurological Disorders. Front. Pharmacol. 2019, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Tsubouchi, K.; Araya, J.; Yoshida, M.; Sakamoto, T.; Koumura, T.; Minagawa, S.; Hara, H.; Hosaka, Y.; Ichikawa, A.; Saito, N.; et al. Involvement of GPx4-Regulated Lipid Peroxidation in Idiopathic Pulmonary Fibrosis Pathogenesis. J. Immunol. 2019, 203, 2076–2087. [Google Scholar] [CrossRef]
- Bedard, K.; Lardy, B.; Krause, K.-H. NOX family NADPH oxidases: Not just in mammals. Biochimie 2007, 89, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Hecker, L. NADPH oxidases: Pathophysiology and therapeutic potential in age-associated pulmonary fibrosis. Redox Biol. 2020, 33, 101541. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Cai, Q.; Yang, D.; Yang, J.; Xue, J.; Yu, M.; Liu, Y.; Ma, F.; Li, F.; Liu, X. A Positive Feed Forward Loop between Wnt/beta-Catenin and NOX4 Promotes Silicon Dioxide-Induced Epithelial-Mesenchymal Transition of Lung Epithelial Cells. Oxid. Med. Cell. Longev. 2020, 2020, 3404168. [Google Scholar] [CrossRef] [PubMed]
- Sang, L.; Guo, X.; Fan, H.; Shi, J.; Hou, S.; Lv, Q. Mesenchymal Stem Cell-Derived Extracellular Vesicles as Idiopathic Pulmonary Fibrosis Microenvironment Targeted Delivery. Cells 2022, 11, 2322. [Google Scholar] [CrossRef] [PubMed]
- Kandhare, A.D.; Bodhankar, S.L.; Mohan, V.; Thakurdesai, P.A. Effect of glycosides based standardized fenugreek seed extract in bleomycin-induced pulmonary fibrosis in rats: Decisive role of Bax, Nrf2, NF-kappaB, Muc5ac, TNF-alpha and IL-1beta. Chem. Biol. Interact. 2015, 237, 151–165. [Google Scholar] [CrossRef]
- Li, F.; Han, F.; Li, H.; Zhang, J.; Qiao, X.; Shi, J.; Yang, L.; Dong, J.; Luo, M.; Wei, J.; et al. Human placental mesenchymal stem cells of fetal origins-alleviated inflammation and fibrosis by attenuating MyD88 signaling in bleomycin-induced pulmonary fibrosis mice. Mol. Immunol. 2017, 90, 11–21. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, W.; Yang, J.; Xue, J.; Ma, J.; Wu, S.; Wang, J.; Xu, R.; Wei, J.; Wang, Y.; Wang, S.; et al. Secretome of hESC-Derived MSC-like Immune and Matrix Regulatory Cells Mitigate Pulmonary Fibrosis through Antioxidant and Anti-Inflammatory Effects. Biomedicines 2023, 11, 463. https://doi.org/10.3390/biomedicines11020463
Hu W, Yang J, Xue J, Ma J, Wu S, Wang J, Xu R, Wei J, Wang Y, Wang S, et al. Secretome of hESC-Derived MSC-like Immune and Matrix Regulatory Cells Mitigate Pulmonary Fibrosis through Antioxidant and Anti-Inflammatory Effects. Biomedicines. 2023; 11(2):463. https://doi.org/10.3390/biomedicines11020463
Chicago/Turabian StyleHu, Wenfeng, Jiali Yang, Jing Xue, Jia Ma, Shuang Wu, Jing Wang, Ranran Xu, Jun Wei, Yujiong Wang, Shuyan Wang, and et al. 2023. "Secretome of hESC-Derived MSC-like Immune and Matrix Regulatory Cells Mitigate Pulmonary Fibrosis through Antioxidant and Anti-Inflammatory Effects" Biomedicines 11, no. 2: 463. https://doi.org/10.3390/biomedicines11020463