
Brain-Type Creatine Kinase Release from Cultured Osteoclasts Exposed to Neridronate in Children Affected by Osteogenesis Imperfecta Type 1
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Cells and Culture Conditions
2.3. Biochemical Assessments
2.4. Bone Quality Measurements
2.5. Statistical Analyses
3. Results
3.1. Bone Status in OI Patients
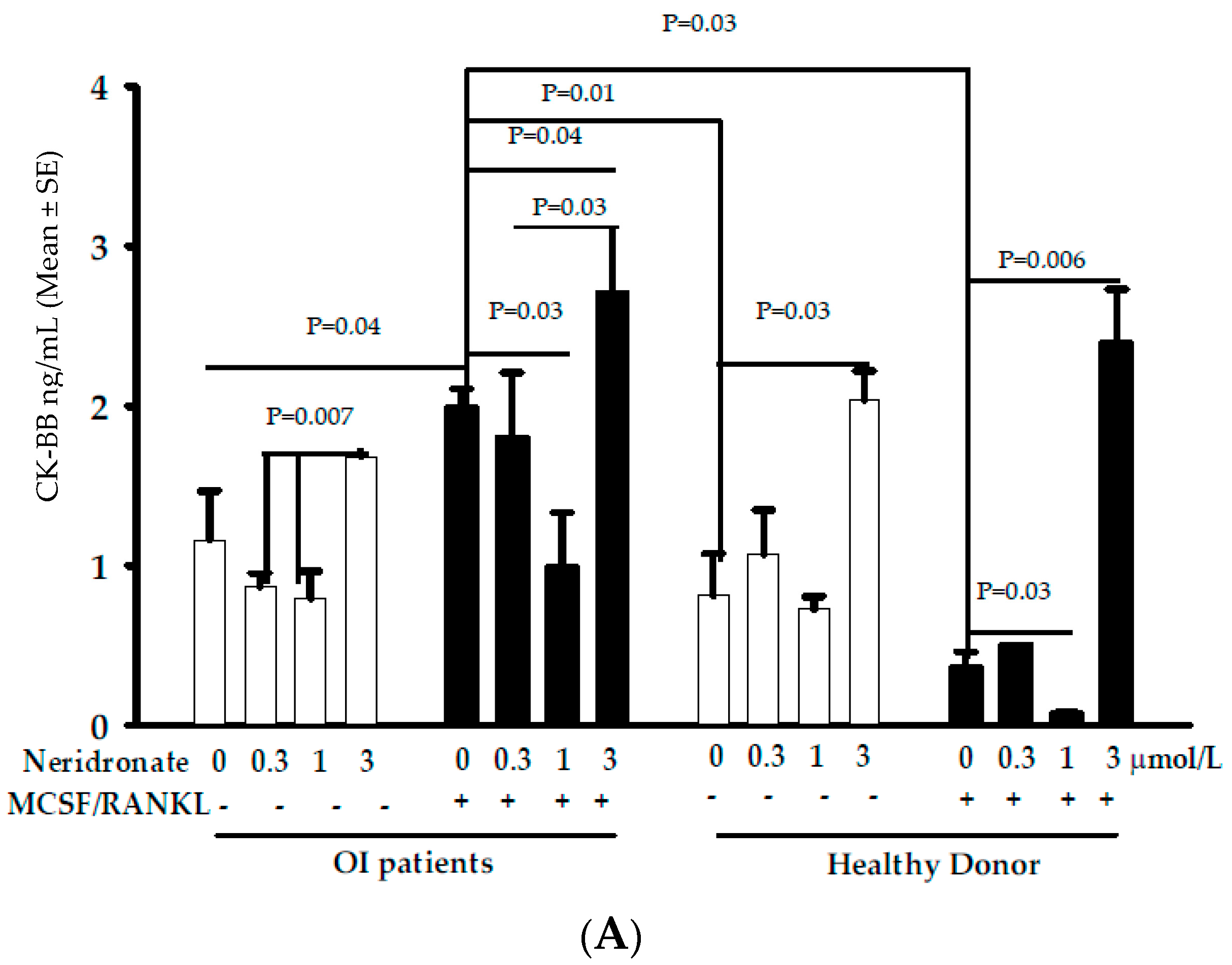
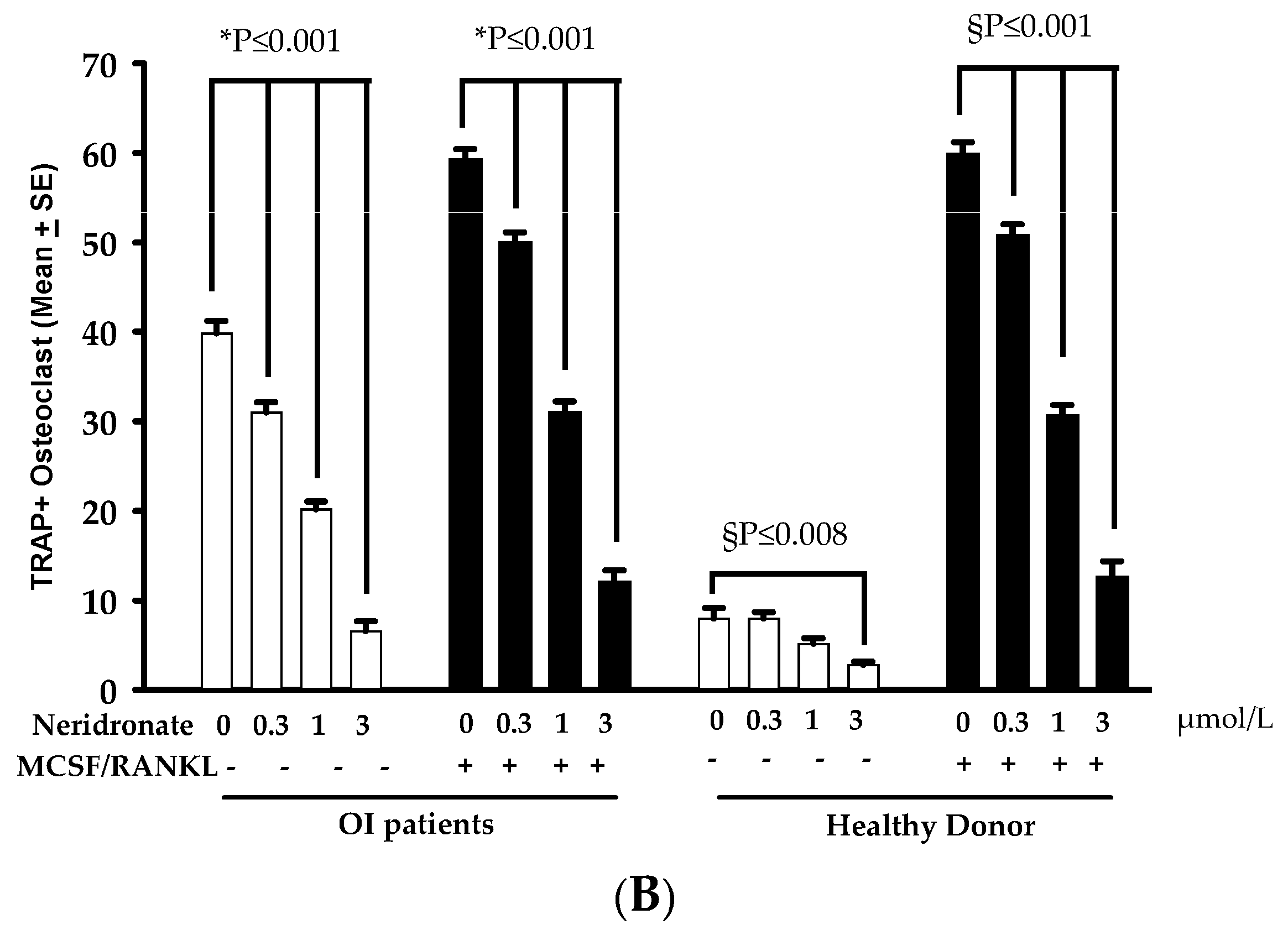
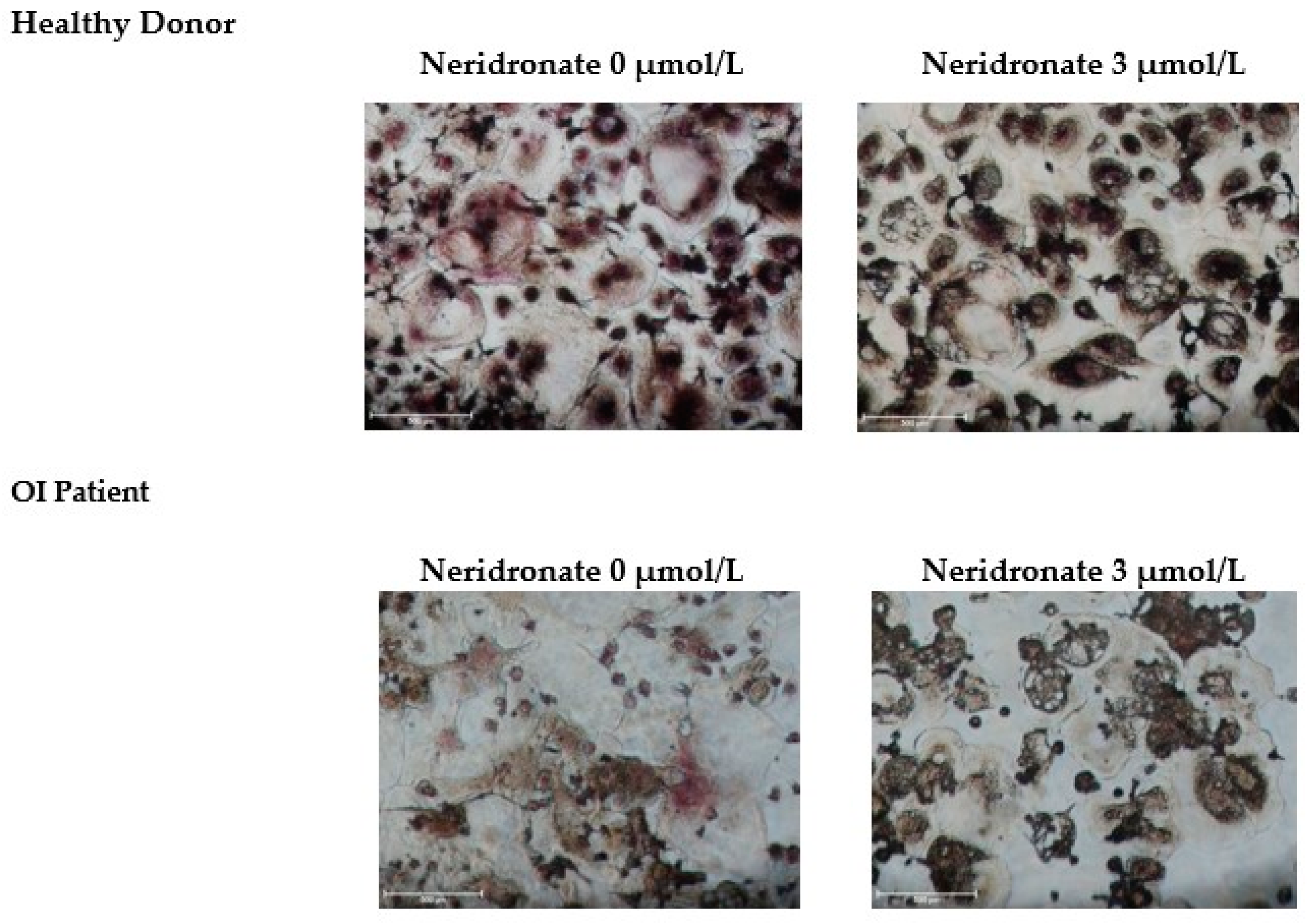
3.2. CK-BB Levels in Osteoclast Culture from OI Patients and Controls
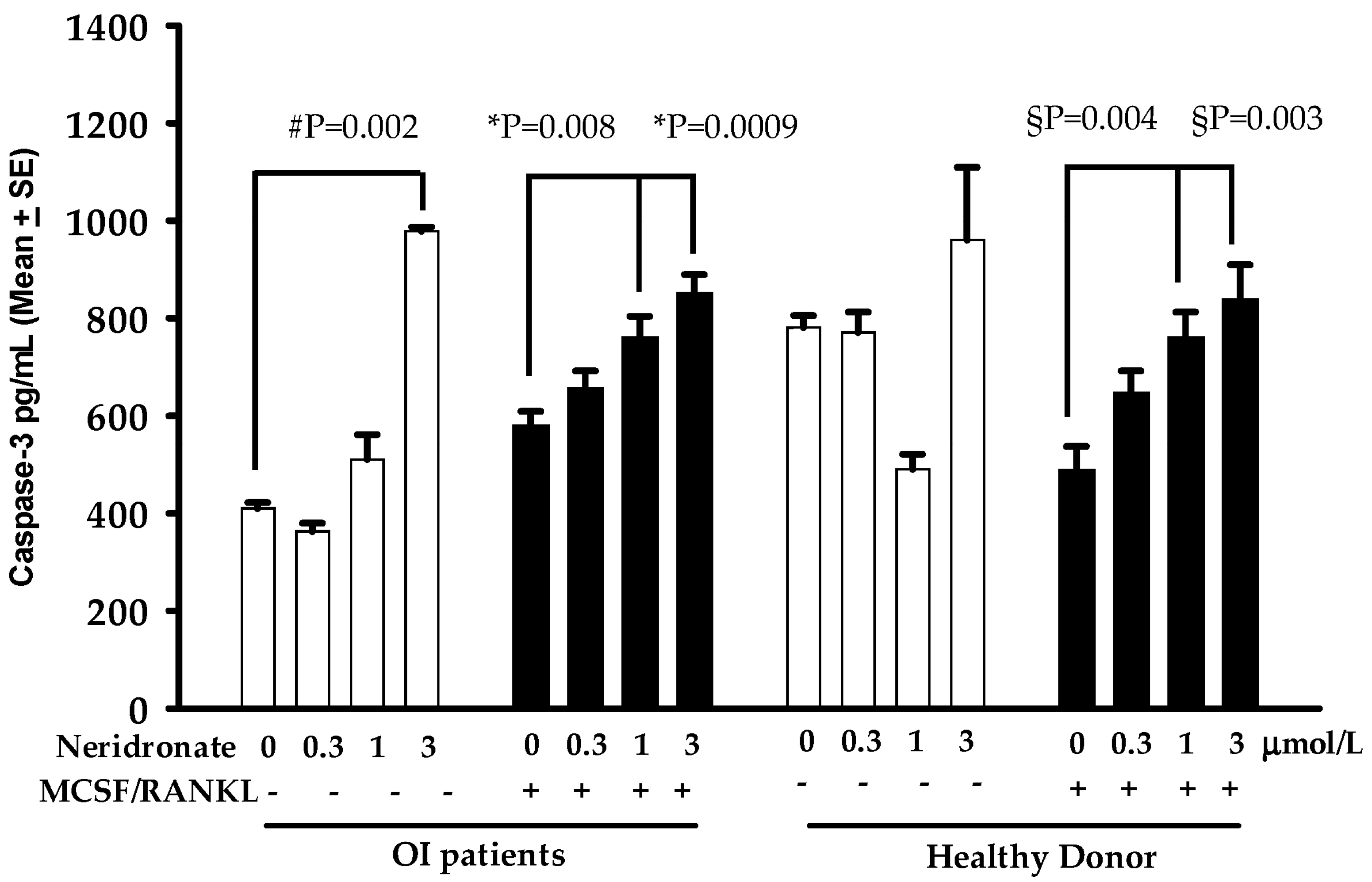
3.3. Caspase-3 Mediated Apoptosis in Neridronate-Treated Cultures
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wallimann, T.; Tokarska-Schlattner, M.; Schlattner, U. The creatine kinase system and pleiotropic effects of creatine. Amino Acids 2011, 40, 1271–1296. [Google Scholar] [CrossRef] [PubMed]
- Eppenberger, H.M.; Dawson, D.M.; Kaplan, N.O. The comparative enzymology of creatine kinases. I. Isolation and characterization from chicken and rabbit tissues. J. Biol. Chem. 1967, 242, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, T.; Wyss, M.; Brdiczka, D.; Nicolay, K.; Eppenberger, H.M. Intracellular compartmentation, structure and function of creatine kinase isoenzymes in tissues with high and fluctuating energy demands: The ‘phosphocreatine circuit’ for cellular energy homeostasis. Biochem. J. 1992, 281, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Hazama, R.; Qu, X.; Yokoyama, K.; Tanaka, C.; Kinoshita, E.; He, J.; Takahashi, S.; Tohyama, K.; Yamamura, H.; Tohyama, Y. ATP-induced osteoclast function: The formation of sealing-zone like structure and the secretion of lytic granules via microtubule-deacetylation under the control of Syk. Genes Cells 2009, 14, 871–884. [Google Scholar] [CrossRef]
- Sakai, D.; Tong, H.S.; Minkin, C. Osteoclast molecular phenotyping by random cDNA sequencing. Bone 1995, 17, 111–119. [Google Scholar] [CrossRef]
- Chen, J.; Sun, Y.; Mao, X.; Liu, Q.; Wu, H.; Chen, Y. RANKL up-regulates brain-type creatine kinase via poly(ADP-ribose) po-lymerase-1 during osteoclastogenesis. J. Biol. Chem. 2010, 285, 36315–36321. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.J.; Ha, J.; Oerlemans, F.; Lee, Y.J.; Lee, S.W.; Ryu, J.; Kim, H.J.; Lee, Y.; Kim, H.M.; Choi, J.Y.; et al. Brain-type creatine kinase has a crucial role in osteoclast-mediated bone resorption. Nat. Med. 2008, 14, 966–972. [Google Scholar] [CrossRef]
- Yoneyama, T.; Fowler, H.L.; Pendleton, J.W.; Sforza, P.P.; Lui, C.Y.; Iranmanesh, A.; Gerard, R.D. Elevated levels of creatine kinase BB isoenzyme in three patients with adult osteopetrosis. N. Engl. J. Med. 1989, 320, 1284–1285. [Google Scholar] [CrossRef]
- Gram, J.; Antonsen, S.; Hørder, M.; Bollerslev, J. Elevated serum levels of creatine kinase BB in autosomal dominant osteopetrosis type II. Calcif. Tissue Int. 1991, 48, 438–439. [Google Scholar] [CrossRef]
- Yoneyama, T.; Fowler, H.L.; Pendleton, J.W.; Sforza, P.P.; Gerard, R.D.; Lui, C.Y.; Eldridge, T.H.; Iranmanesh, A. Elevated serum levels of creatine kinase BB in autosomal dominant osteopetrosis type II--a family study. Clin. Genet. 1992, 42, 39–42. [Google Scholar] [CrossRef]
- Whyte, M.P.; Wenkert, D.; Clements, K.L.; McAlister, W.H.; Mumm, S. Bisphosphonate-induced osteopetrosis. N. Engl. J. Med. 2003, 349, 457–463. [Google Scholar] [CrossRef]
- D’Eufemia, P.; Finocchiaro, R.; Villani, C.; Zambrano, A.; Lodato, V.; Palombaro, M.; Properzi, E.; Celli, M. Serum brain-type creatine kinase increases in children with osteogenesis imperfecta during neridronate treatment. Pediatr. Res. 2014, 75, 626–630. [Google Scholar] [CrossRef] [PubMed]
- D’Eufemia, P.; Finocchiaro, R.; Zambrano, A.; Lodato, V.; Celli, L.; Finocchiaro, S.; Persiani, P.; Turchetti, A.; Celli, M. Serum creatine kinase isoenzymes in children with osteogenesis imperfecta. Osteoporos. Int. 2017, 28, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Rauch, F.; Glorieux, F.H. Osteogenesis imperfecta. Lancet 2004, 363, 1377–1385. [Google Scholar] [CrossRef]
- Maruotti, N.; Corrado, A.; Neve, A.; Cantatore, F.P. Bisphosphonates: Effects on osteoblast. Eur. J. Clin. Pharmacol. 2012, 68, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, F.H. The use of bisphosphonates in children with osteogenesis imperfecta. J. Pediatr. Endocrinol. Metab. 2001, 14 (Suppl. S6), 1491–1495. [Google Scholar] [PubMed]
- Whyte, M.P.; McAlister, W.H.; Novack, D.V.; Clements, K.L.; Schoenecker, P.L.; Wenkert, D. Bisphosphonate-induced osteopetrosis: Novel bone modeling defects, metaphyseal osteopenia, and osteosclerosis fractures after drug exposure ceases. J. Bone Miner. Res. 2008, 23, 1698–1707. [Google Scholar] [CrossRef]
- Tanaka, M.; Mori, H.; Kayasuga, R.; Kawabata, K. Induction of creatine kinase release from cultured osteoclasts via the pharmacological action of aminobisphosphonates. Springerplus 2015, 4, 59. [Google Scholar] [CrossRef] [PubMed]
- Sillence, D.O.; Senn, A.; Danks, D.M. Genetic heterogeneity in osteogenesis imperfecta. J. Med. Genet 1979, 16, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Manuals and User Guides for OsteoSys SONOST 3000. Available online: https://www.manualslib.com/products/Osteosys-Sonost-3000-8710830.html (accessed on 6 November 2022).
- Tanner, J.M.; Whitehouse, R.H. Clinical longitudinal standards for height, weight, height velocity, weight velocity, and stages of puberty. Arch. Dis. Child. 1976, 51, 170–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunetti, G.; Papadia, F.; Tummolo, A.; Fischetto, R.; Nicastro, F.; Piacente, L.; Ventura, A.; Mori, G.; Oranger, A.; Gigante, I.; et al. Impaired bone remodeling in children with osteogenesis imperfecta treated and untreated with bisphosphonates: The role of DKK1, RANKL, and TNF-α. Osteoporos. Int. 2016, 27, 2355–2365. [Google Scholar] [CrossRef]
- Brunetti, G.; D’Amato, G.; Chiarito, M.; Tullo, A.; Colaianni, G.; Colucci, S.; Grano, M.; Faienza, M.F. An update on the role of RANKL-RANK/osteoprotegerin and WNT-ß-catenin signaling pathways in pediatric diseases. World J. Pediatr. 2019, 15, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Galeone, A.; Brunetti, G.; Rotunno, C.; Oranger, A.; Colucci, S.; de Luca Tupputi Schinosa, L.; Zallone, A.; Grano, M.; Paparella, D. Activation of the receptor activator of the nuclear factor-κB ligand pathway during coronary bypass surgery: Comparison between on- and off-pump coronary artery bypass surgery procedures. Eur. J. Cardiothorac. Surg. 2013, 44, e141–e147. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jiang, X.; Delaney, J.; Franceschetti, T.; Bilic-Curcic, I.; Kalinovsky, J.; Lorenzo, J.A.; Grcevic, D.; Rowe, D.W.; Kalajzic, I. Immature osteoblast lineage cells increase osteoclastogenesis in osteogenesis imperfecta murine. Am. J. Pathol. 2010, 176, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Forlino, A.; Marini, J.C. Osteogenesis imperfecta. Lancet 2016, 387, 1657–1671. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.J. New insights into the molecular mechanisms of action of bisphosphonates. Curr. Pharm. Des. 2003, 9, 2643–2658. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.G.; Watts, N.B.; Ebetino, F.H.; Rogers, M.J. Mechanisms of action of bisphosphonates: Similarities and differences and their potential influence on clinical efficacy. Osteoporos. Int. 2008, 19, 733–759. [Google Scholar] [CrossRef]
- Ebetino, F.H.; Hogan, A.M.; Sun, S.; Tsoumpra, M.K.; Duan, X.; Triffitt, J.T.; Kwaasi, A.A.; Dunford, J.E.; Barnett, B.L.; Lundy, M.W.; et al. The relationship between the chemistry and biological activity of the bisphosphonates. Bone 2011, 49, 20–33. [Google Scholar] [CrossRef]
- Nishida, S.; Kikuichi, S.; Haga, H.; Yoshioka, S.; Tsubaki, M.; Fujii, K.; Irimajiri, K. Apoptosis-inducing effect of a new bisphosphonate, YM529, on various hematopoietic tumor cell lines. Biol. Pharm. Bull. 2003, 26, 96–100. [Google Scholar] [CrossRef]
- Coxon, F.P.; Helfrich, M.H.; Larijani, B.; Muzylak, M.; Dunford, J.E.; Marshall, D.; McKinnon, A.D.; Nesbitt, S.A.; Horton, M.A.; Seabra, M.C.; et al. Identification of a novel phosphonocarboxylate inhibitor of Rab geranylgeranyl transferase that specifically prevents Rab prenylation in osteoclasts and macrophages. J. Biol. Chem. 2001, 276, 48213–48222. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, R.; Hagino, H.; Ito, M.; Sone, T.; Nakamura, T.; Mizunuma, H.; Fukunaga, M.; Shiraki, M.; Nishizawa, Y.; Ohashi, Y.; et al. Efficacy and safety of monthly oral minodronate in patients with involutional osteoporosis. Osteoporos. Int. 2012, 23, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Tai, T.W.; Chen, C.Y.; Su, F.C.; Tu, Y.K.; Tsai, T.T.; Lin, C.F.; Jou, I.M. Reactive oxygen species are required for zoledronic acid-induced apoptosis in osteoclast precursors and mature osteoclast-like cells. Sci. Rep. 2017, 7, 44245. [Google Scholar] [CrossRef] [PubMed]
- Rauch, F.; Travers, R.; Parfitt, A.M.; Glorieux, F.H. Static and dynamic bone histomorphometry in children with osteogenesis imperfecta. Bone 2000, 26, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Zeitlin, L.; Fassier, F.; Glorieux, F.H. Modern approach to children with osteogenesis imperfecta. J. Pediatr. Orthop. B 2003, 12, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Doty, S.B.; Hughes, C.; Dempster, D.; Camacho, N.P. Increased resorptive activity and accompanying morphological alterations in osteoclasts derived from the oim/oim mouse model of osteogenesis imperfecta. J. Cell. Biochem. 2007, 102, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Uveges, T.E.; Collin-Osdoby, P.; Cabral, W.A.; Ledgard, F.; Goldberg, L.; Bergwitz, C.; Forlino, A.; Osdoby, P.; Gronowicz, G.A.; Marini, J.C. Cellular mechanism of decreased bone in Brtl mouse model of OI: Imbalance of decreased osteoblast function and increased osteoclasts and their precursors. J. Bone Miner. Res. 2008, 23, 1983–1994. [Google Scholar] [CrossRef] [PubMed]
- Sistermans, E.A.; de Kok, Y.J.; Peters, W.; Ginsel, L.A.; Jap, P.H.; Wieringa, B. Tissue- and cell-specific distribution of creatine kinase B: A new and highly specific monoclonal antibody for use in immunohistochemistry. Cell Tissue Res. 1995, 280, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Somjen, D.; Katzburg, S.; Sharon, O.; Grafi-Cohen, M.; Knoll, E.; Stern, N. The effects of estrogen receptors α- and β-specific agonists and antagonists on cell proliferation and energy metabolism in human bone cell line. J. Cell. Biochem. 2011, 112, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Decking, U.K.; Alves, C.; Wallimann, T.; Wyss, M.; Schrader, J. Functional aspects of creatine kinase isoenzymes in endothelial cells. Am. J. Physiol. Cell Physiol. 2001, 281, C320–C328. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient 1 | Patient 2 | Patient 3 | Patient 4 | Patient 5 | |
|---|---|---|---|---|---|
| Gender | M | F | F | F | M |
| Age (years) | 8.5 | 5.9 | 14.2 | 18.5 | 7.5 |
| Height SDS | 1.71 | 3.04 | −2.32 | −2.11 | −0.93 |
| Weight SDS | 0.95 | 1.98 | 0.27 | −0.76 | 1.22 |
| BMI SDS | 0.07 | 0.89 | 1.27 | 0.37 | 1.97 |
| Tanner stage [21] | I | I | IV | V | I |
| Blu sclera (Yes/no) | no | yes | yes | yes | yes |
| Dentinogenesis imperfecta (yes/no) | no | no | yes | no | yes |
| Number of fractures | 1 | 1 | 4 | 1 | 2 |
| Scoliosis (yes/no) | no | no | yes | no | yes |
| Limited mobility (yes/no) | no | no | no | no | yes |
| Leg deformities (yes/no) | Tibial valgus | Tibial valgus | Lower limb dysmetria (left > right) | No | Coxa and femoral valgus; bilateral clubfoot; lower limb dysmetria (right > left) |
| QUS parameters | |||||
| BUA (dB/MHz) | 81 | 36.1 | 56.2 | 117.6 | 34.8 |
| SOS (m/s) | 1486.2 | 1477.2 | 1478.8 | 1540 | 1490.3 |
| BQI SDS | −1.6 | −2.6 | −3.5 | 0.1 | −2.6 |
| CTX-I (0.034–0.635 ng/mL) | 1.2 | 1.5 | 0.293 | 0.335 | 1.308 |
| TRAcP5b (1.2–4.8 U/L) | 2.6 | 2.3 | 2.8 | 2.4 | 2.1 |
| sRANKL (<1000 pmol/L) | 36.4 | 48.7 | 4 | 32.4 | 87.2 |
| OPG (4.37–5.03 pmol/L) | 3.15 | 3.21 | 4.14 | 2.82 | 1.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faienza, M.F.; Tummolo, A.; Celli, M.; Finocchiaro, R.; Piacente, L.; Di Serio, F.; Nicchia, G.P.; Brunetti, G.; D’Eufemia, P. Brain-Type Creatine Kinase Release from Cultured Osteoclasts Exposed to Neridronate in Children Affected by Osteogenesis Imperfecta Type 1. Biomedicines 2023, 11, 458. https://doi.org/10.3390/biomedicines11020458
Faienza MF, Tummolo A, Celli M, Finocchiaro R, Piacente L, Di Serio F, Nicchia GP, Brunetti G, D’Eufemia P. Brain-Type Creatine Kinase Release from Cultured Osteoclasts Exposed to Neridronate in Children Affected by Osteogenesis Imperfecta Type 1. Biomedicines. 2023; 11(2):458. https://doi.org/10.3390/biomedicines11020458
Chicago/Turabian StyleFaienza, Maria Felicia, Albina Tummolo, Mauro Celli, Roberto Finocchiaro, Laura Piacente, Francesca Di Serio, Grazia Paola Nicchia, Giacomina Brunetti, and Patrizia D’Eufemia. 2023. "Brain-Type Creatine Kinase Release from Cultured Osteoclasts Exposed to Neridronate in Children Affected by Osteogenesis Imperfecta Type 1" Biomedicines 11, no. 2: 458. https://doi.org/10.3390/biomedicines11020458