
Chimeric Newcastle Disease Virus Vectors Expressing Human IFN-γ Mediate Target Immune Responses and Enable Multifaceted Treatments
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Recombinant Chimeric NDV Genome Construction and Rescue Experiment
2.3. Immunofluorescence Assay
2.4. Viral Replication, Neutralization, and IFN-γ RNA Expression Assays
2.5. Cytotoxicity Assay
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Killing Assay
2.8. Statistical Analysis
3. Results
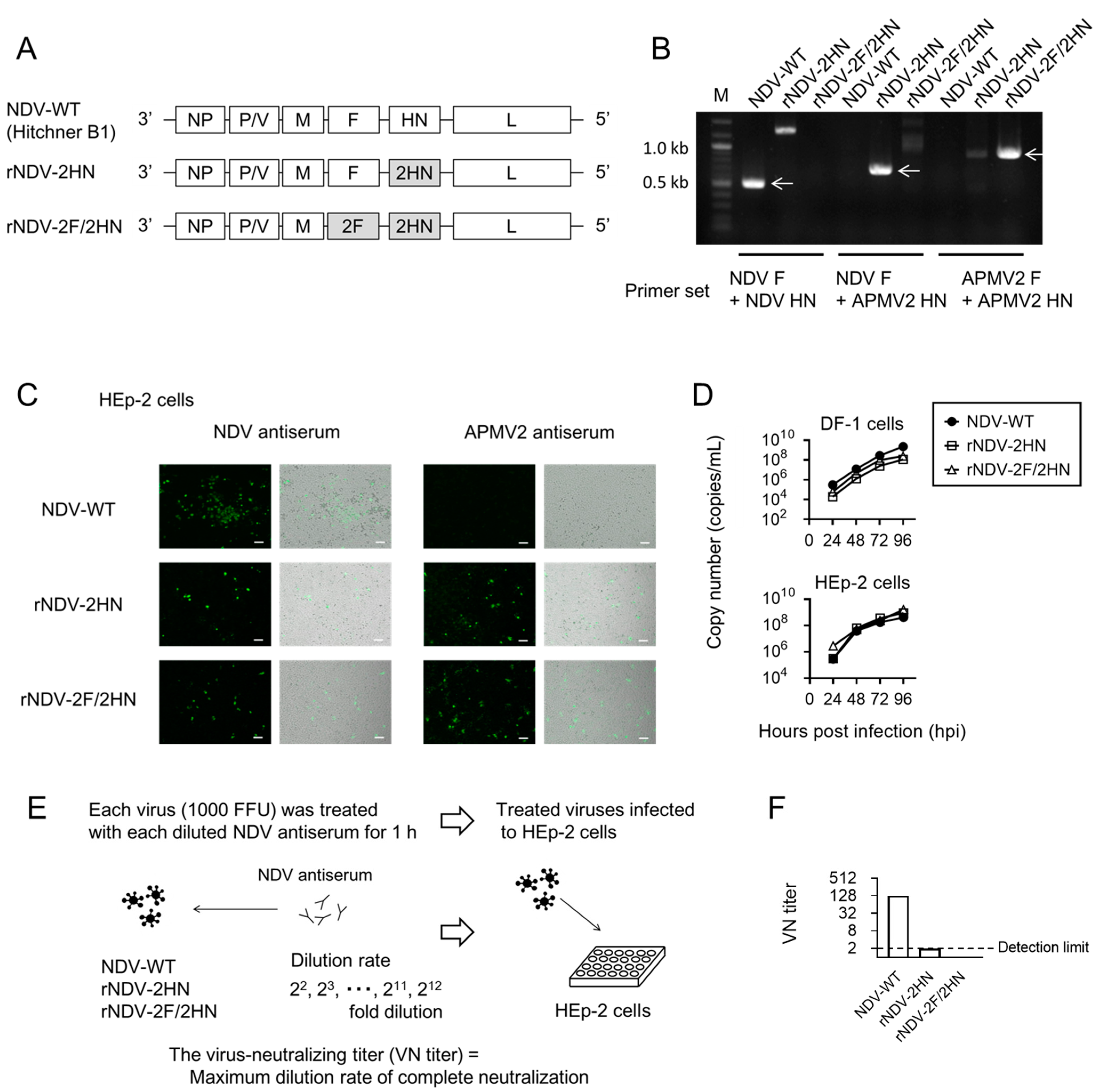
3.1. Two Types of Infectious Chimeric NDVs Were Constructed
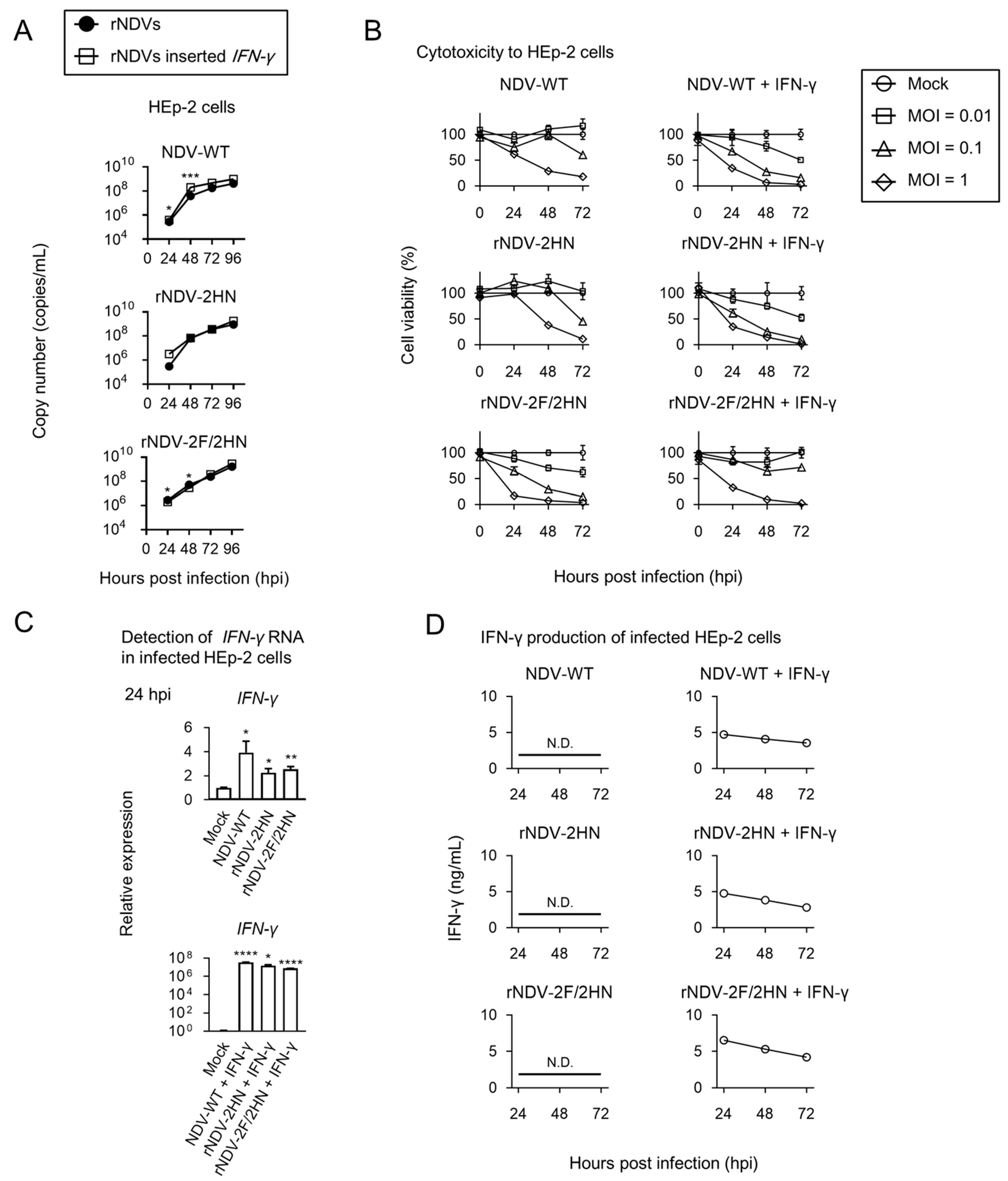
3.2. Recombinant NDVs with Inserted Human IFN-γ Gene Were Constructed and Indicated Low Pathogenicity
3.3. Recombinant NDVs + IFN-γ Induced production of Sufficient IFN-γ in the Infected HEp-2 Cells
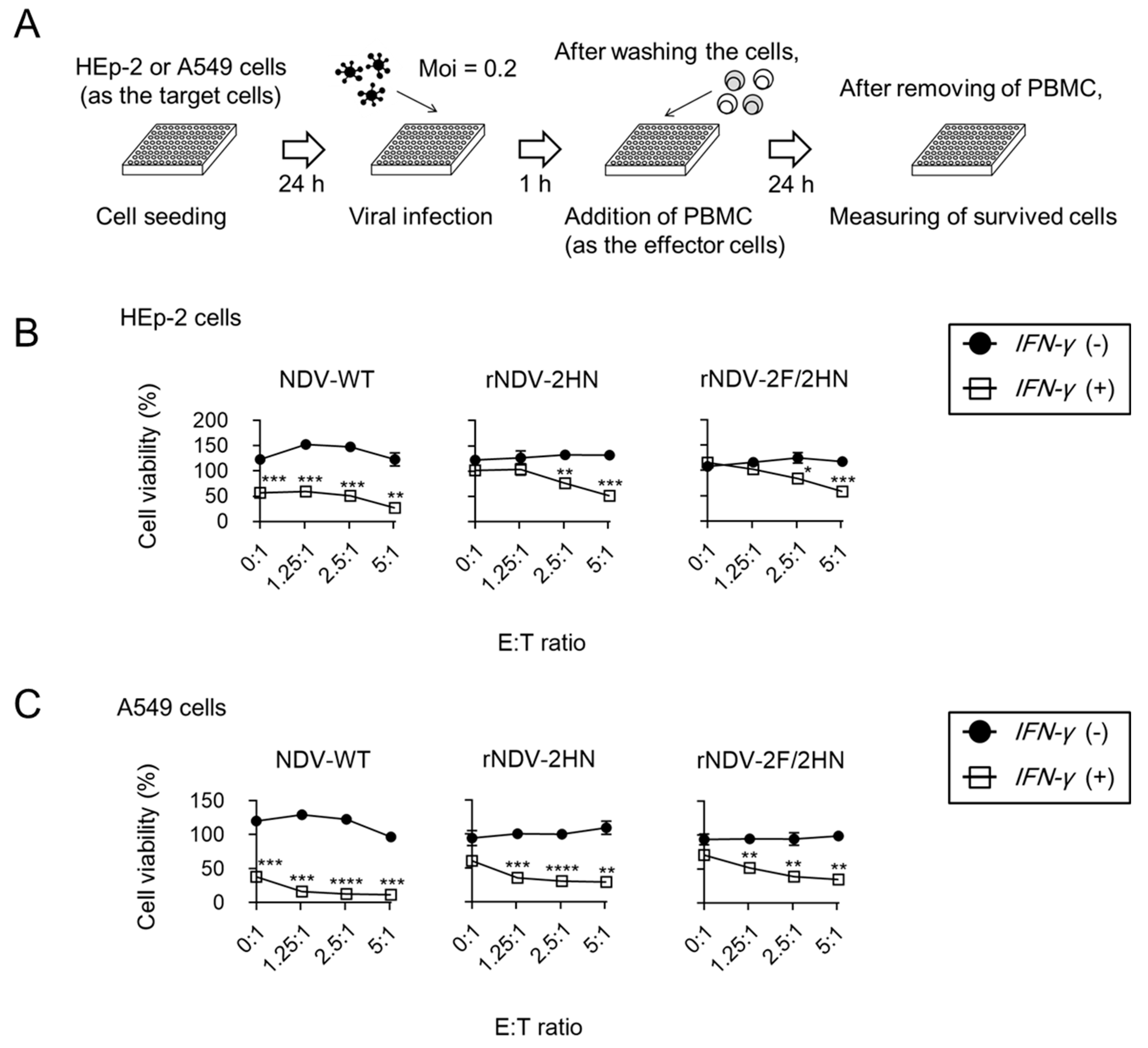
3.4. Recombinant NDVs + IFN-γ Lead to Anti-tumor Responses of PBMC
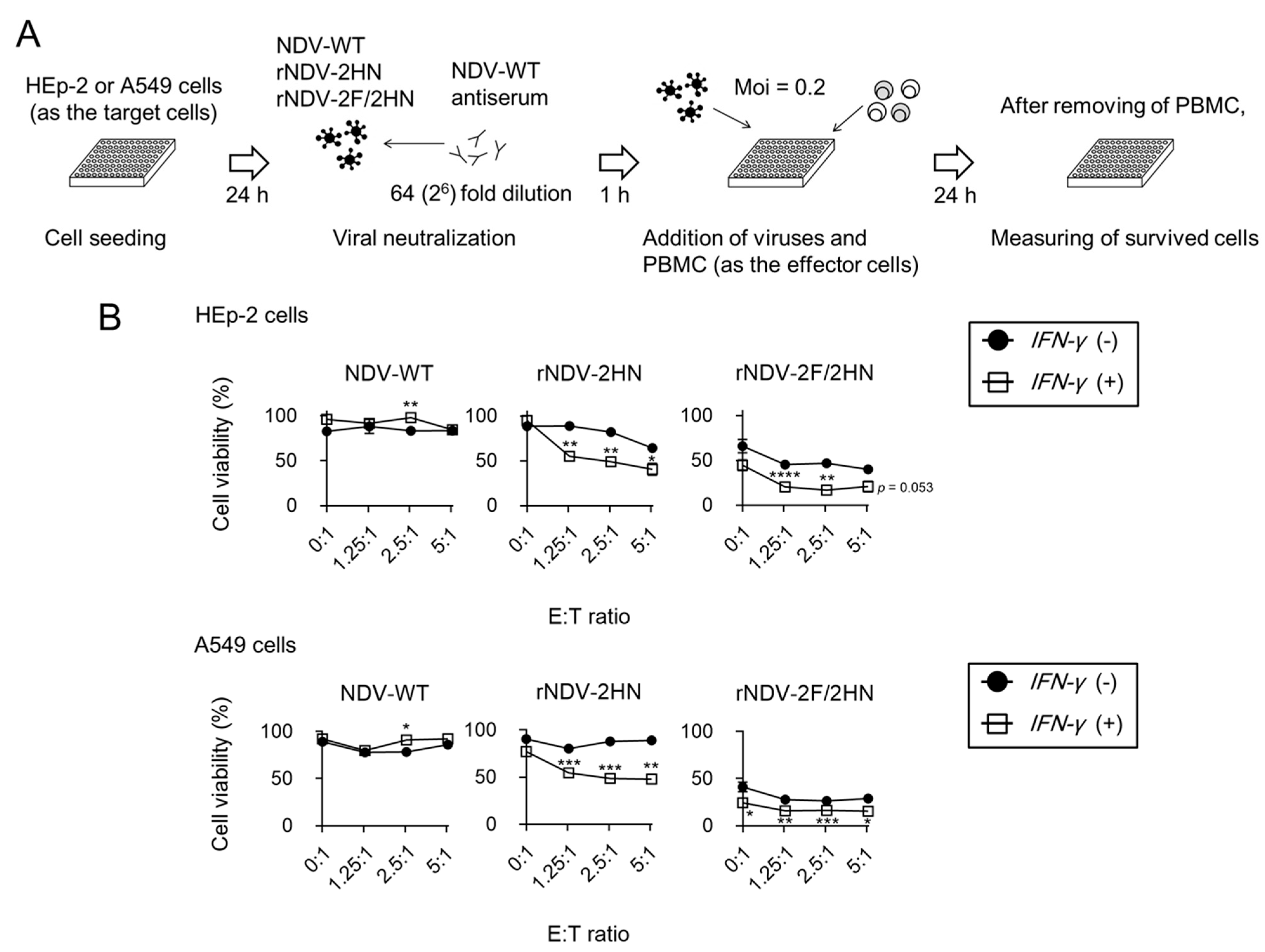
3.5. Infection of rNDV-2HN and 2F/2HN + IFN-γ Induced Anti-tumor Responses in the Presence of Anti-NDV Antiserum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mayers, J.; Mansfield, K.L.; Brown, I.H. The role of vaccination in risk mitigation and control of Newcastle disease in poultry. Vaccine 2017, 35, 5974–5980. [Google Scholar] [CrossRef] [PubMed]
- Gogoi, P.; Ganar, K.; Kumar, S. Avian Paramyxovirus: A Brief Review. Transbound. Emerg. Dis. 2017, 64, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J. Newcastle disease and other avian paramyxoviruses. Rev. Sci. Tech. 2000, 19, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Fiola, C.; Peeters, B.; Fournier, P.; Arnold, A.; Bucur, M.; Schirrmacher, V. Tumor selective replication of Newcastle disease virus: Association with defects of tumor cells in antiviral defence. Int. J. Cancer 2006, 119, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Zamarin, D.; Palese, P. Oncolytic Newcastle disease virus for cancer therapy: Old challenges and new directions. Future Microbiol. 2012, 7, 347–367. [Google Scholar] [CrossRef] [PubMed]
- Syed Najmuddin, S.U.F.; Amin, Z.M.; Tan, S.W.; Yeap, S.K.; Kalyanasundram, J.; Veerakumarasivam, A.; Chan, S.C.; Chia, S.L.; Yusoff, K.; Alitheen, N.B. Oncolytic effects of the recombinant Newcastle disease virus, rAF-IL12, against colon cancer cells in vitro and in tumor-challenged NCr-Foxn1nu nude mice. PeerJ 2020, 8, e9761. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S. Newcastle disease virus vectored vaccines as bivalent or antigen delivery vaccines. Clin. Exp. Vaccine Res. 2017, 6, 72–82. [Google Scholar] [CrossRef]
- Fulber, J.P.C.; Kamen, A.A. Development and Scalable Production of Newcastle Disease Virus-Vectored Vaccines for Human and Veterinary Use. Viruses 2022, 14, 975. [Google Scholar] [CrossRef]
- Spackman, E.; Pantin-Jackwood, M.J. Practical aspects of vaccination of poultry against avian influenza virus. Vet. J. 2014, 202, 408–415. [Google Scholar] [CrossRef]
- Steglich, C.; Grund, C.; Ramp, K.; Breithaupt, A.; Hoper, D.; Keil, G.; Veits, J.; Ziller, M.; Granzow, H.; Mettenleiter, T.C.; et al. Chimeric newcastle disease virus protects chickens against avian influenza in the presence of maternally derived NDV immunity. PLoS ONE 2013, 8, e72530. [Google Scholar] [CrossRef] [Green Version]
- Tsunekuni, R.; Hikono, H.; Saito, T. Evaluation of avian paramyxovirus serotypes 2 to 10 as vaccine vectors in chickens previously immunized against Newcastle disease virus. Vet. Immunol. Immunopathol. 2014, 160, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Paldurai, A.; Samal, S.K. A novel chimeric Newcastle disease virus vectored vaccine against highly pathogenic avian influenza virus. Virology 2017, 503, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xue, L.; Hu, S.; Cheng, H.; Deng, Y.; Hu, Z.; Wang, X.; Liu, X. Chimeric Newcastle disease virus-vectored vaccine protects chickens against H9N2 avian influenza virus in the presence of pre-existing NDV immunity. Arch. Virol. 2018, 163, 3365–3371. [Google Scholar] [CrossRef] [PubMed]
- Earl, P.L.; Cooper, N.; Wyatt, L.S.; Moss, B.; Carroll, M.W. Preparation of cell cultures and vaccinia virus stocks. Curr. Protoc. Mol. Biol. 2001, 43, 16.16.1–16.16.13. [Google Scholar] [CrossRef]
- Nishioka, K.; Daidoji, T.; Nakaya, T. Downregulation of calcium-regulated heat stable protein 1 expression by low-temperature stimulation causes reduction of interferon-beta expression and sensitivity to influenza viral infection. Virus Res. 2022, 309, 198659. [Google Scholar] [CrossRef]
- Brosius, J. Superpolylinkers in cloning and expression vectors. DNA 1989, 8, 759–777. [Google Scholar] [CrossRef]
- Nakaya, T.; Cros, J.; Park, M.S.; Nakaya, Y.; Zheng, H.; Sagrera, A.; Villar, E.; Garcia-Sastre, A.; Palese, P. Recombinant Newcastle disease virus as a vaccine vector. J. Virol. 2001, 75, 11868–11873. [Google Scholar] [CrossRef]
- Hanson, R.P.; Brandly, C.A. Identification of vaccine strains of Newcastle disease virus. Science 1955, 122, 156–157. [Google Scholar] [CrossRef]
- Yan, Y.; Rout, S.N.; Kim, S.H.; Samal, S.K. Role of untranslated regions of the hemagglutinin-neuraminidase gene in replication and pathogenicity of newcastle disease virus. J. Virol. 2009, 83, 5943–5946. [Google Scholar] [CrossRef]
- Kim, S.H.; Yan, Y.; Samal, S.K. Role of the cytoplasmic tail amino acid sequences of Newcastle disease virus hemagglutinin-neuraminidase protein in virion incorporation, cell fusion, and pathogenicity. J. Virol. 2009, 83, 10250–10255. [Google Scholar] [CrossRef] [Green Version]
- Hiam-Galvez, K.J.; Allen, B.M.; Spitzer, M.H. Systemic immunity in cancer. Nat. Rev. Cancer 2021, 21, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Duverge, A.; Negroni, M. Pseudotyping Lentiviral Vectors: When the Clothes Make the Virus. Viruses 2020, 12, 1311. [Google Scholar] [CrossRef] [PubMed]
- Whitt, M.A. Generation of VSV pseudotypes using recombinant DeltaG-VSV for studies on virus entry, identification of entry inhibitors, and immune responses to vaccines. J. Virol. Methods 2010, 169, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Belshe, R.B. The origins of pandemic influenza--lessons from the 1918 virus. N. Engl. J. Med. 2005, 353, 2209–2211. [Google Scholar] [CrossRef] [PubMed]
- Burman, B.; Pesci, G.; Zamarin, D. Newcastle Disease Virus at the Forefront of Cancer Immunotherapy. Cancers 2020, 12, 3552. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Subbiah, M.; Samuel, A.S.; Collins, P.L.; Samal, S.K. Roles of the fusion and hemagglutinin-neuraminidase proteins in replication, tropism, and pathogenicity of avian paramyxoviruses. J. Virol. 2011, 85, 8582–8596. [Google Scholar] [CrossRef]
- Wu, Y.; He, J.; An, Y.; Wang, X.; Liu, Y.; Yan, S.; Ye, X.; Qi, J.; Zhu, S.; Yu, Q.; et al. Recombinant Newcastle disease virus (NDV/Anh-IL-2) expressing human IL-2 as a potential candidate for suppresses growth of hepatoma therapy. J. Pharmacol. Sci. 2016, 132, 24–30. [Google Scholar] [CrossRef]
- Mohamed Amin, Z.; Che Ani, M.A.; Tan, S.W.; Yeap, S.K.; Alitheen, N.B.; Syed Najmuddin, S.U.F.; Kalyanasundram, J.; Chan, S.C.; Veerakumarasivam, A.; Chia, S.L.; et al. Evaluation of a Recombinant Newcastle Disease Virus Expressing Human IL12 against Human Breast Cancer. Sci. Rep. 2019, 9, 13999. [Google Scholar] [CrossRef]
- Vigil, A.; Park, M.S.; Martinez, O.; Chua, M.A.; Xiao, S.; Cros, J.F.; Martinez-Sobrido, L.; Woo, S.L.; Garcia-Sastre, A. Use of reverse genetics to enhance the oncolytic properties of Newcastle disease virus. Cancer Res. 2007, 67, 8285–8292. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Mean Death Time |
|---|---|
| NDV-WT | >168 h |
| NDV-WT + IFN-γ | >168 h |
| rNDV-2HN | >168 h |
| rNDV-2HN + IFN-γ | >168 h |
| rNDV-2F/2HN | >168 h |
| rNDV-2F/2HN + IFN-γ | >168 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soliman, R.M.; Nishioka, K.; Daidoji, T.; Noyori, O.; Nakaya, T. Chimeric Newcastle Disease Virus Vectors Expressing Human IFN-γ Mediate Target Immune Responses and Enable Multifaceted Treatments. Biomedicines 2023, 11, 455. https://doi.org/10.3390/biomedicines11020455
Soliman RM, Nishioka K, Daidoji T, Noyori O, Nakaya T. Chimeric Newcastle Disease Virus Vectors Expressing Human IFN-γ Mediate Target Immune Responses and Enable Multifaceted Treatments. Biomedicines. 2023; 11(2):455. https://doi.org/10.3390/biomedicines11020455
Chicago/Turabian StyleSoliman, Rofaida Mostafa, Keisuke Nishioka, Tomo Daidoji, Osamu Noyori, and Takaaki Nakaya. 2023. "Chimeric Newcastle Disease Virus Vectors Expressing Human IFN-γ Mediate Target Immune Responses and Enable Multifaceted Treatments" Biomedicines 11, no. 2: 455. https://doi.org/10.3390/biomedicines11020455