
Distinctive Neuroanatomic Regions Involved in Cocaine-Induced Behavioral Sensitization in Mice
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Groups
2.3. Drugs
2.4. Open-Field Locomotor Activity
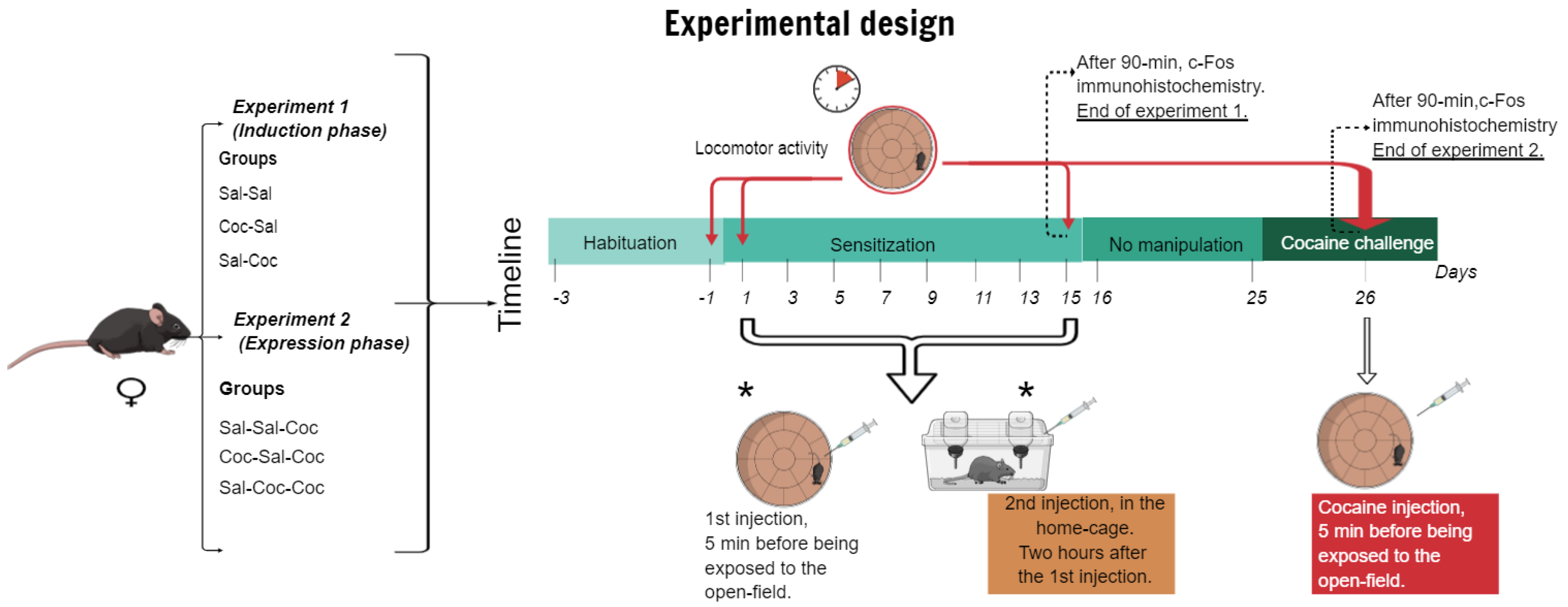
2.5. Experimental Procedures
2.5.1. Experiment 1: Characterization of the Induction Phase of Cocaine-Induced Hyperlocomotion
2.5.2. Experiment 2: Characterization of the Expression Phase of Cocaine-Induced Hyperlocomotion
2.6. Tissue Preparation
2.7. c-Fos Protein Expression
2.8. Stereological Analysis
2.9. Statistical Analysis
3. Results
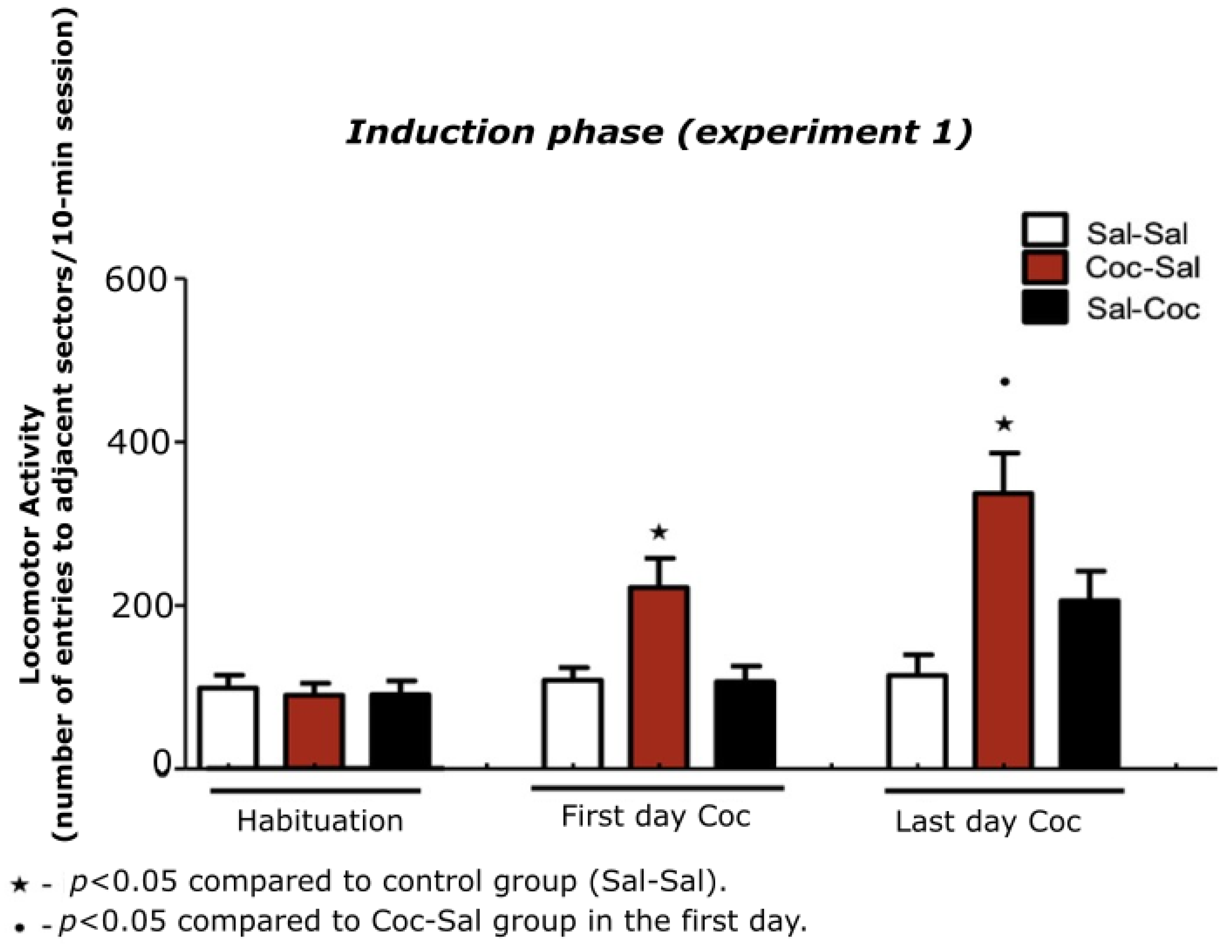
3.1. Experiment 1
3.1.1. Characterization of the Induction Phase of Cocaine-Induced Hyperlocomotion
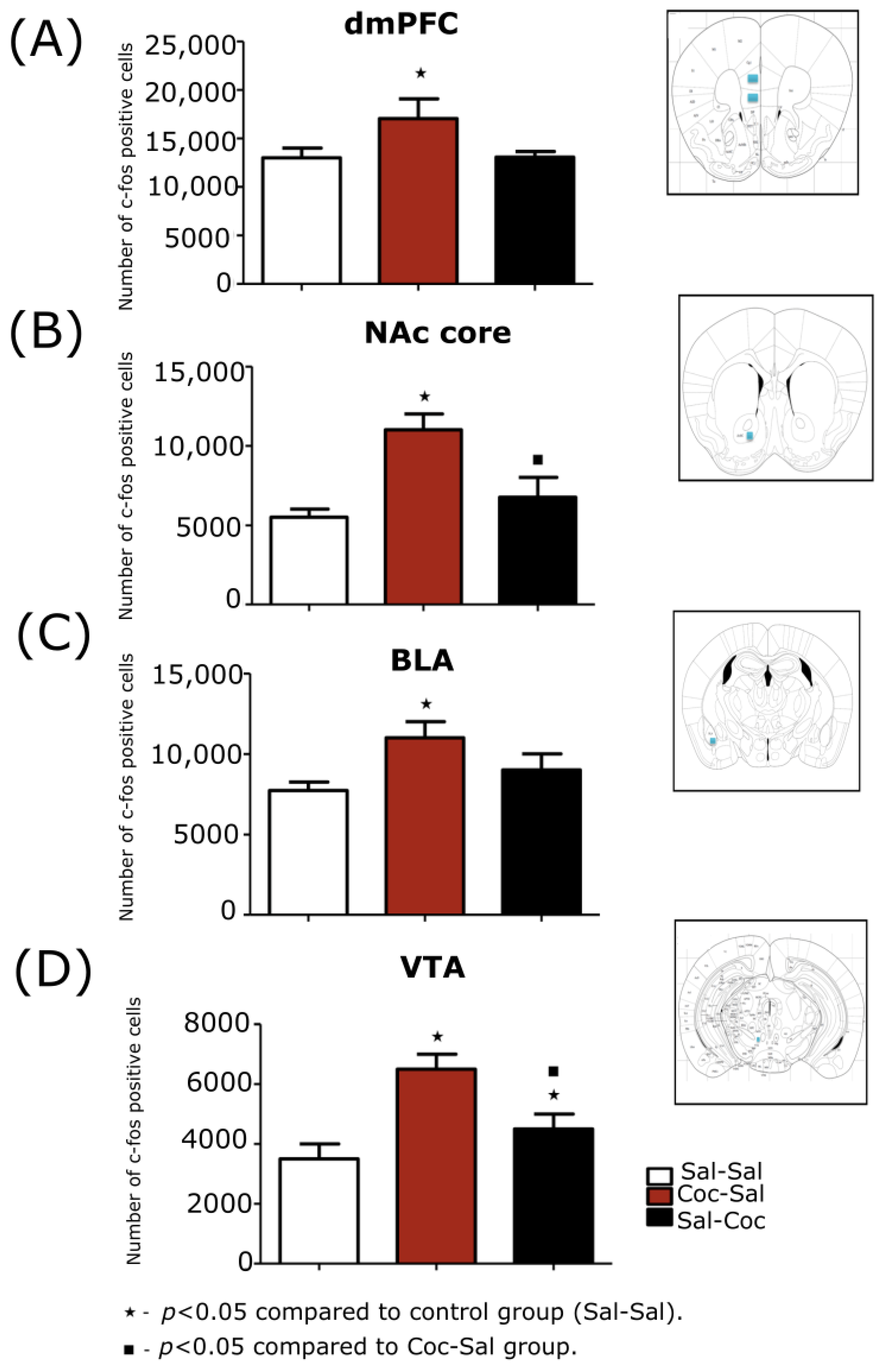
3.1.2. Stereological Evaluations of the Expression Phase of Cocaine-Induced Hyperlo-comotion
dmPFC
NAc Core
NAc Shell
BLA
VTA
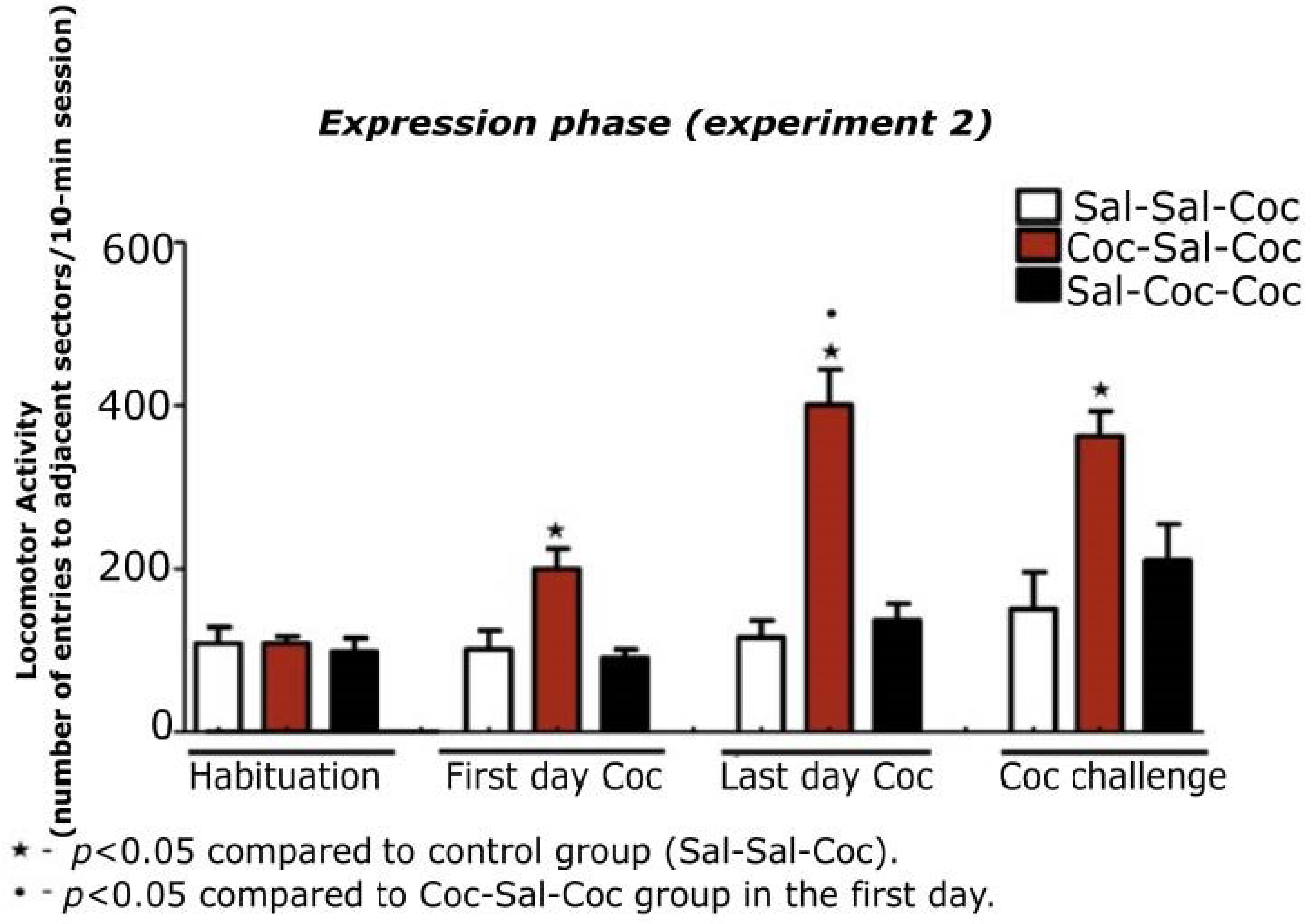
3.2. Experiment 2
3.2.1. Characterization of the Expression Phase of Cocaine-Induced Hyperlocomotion
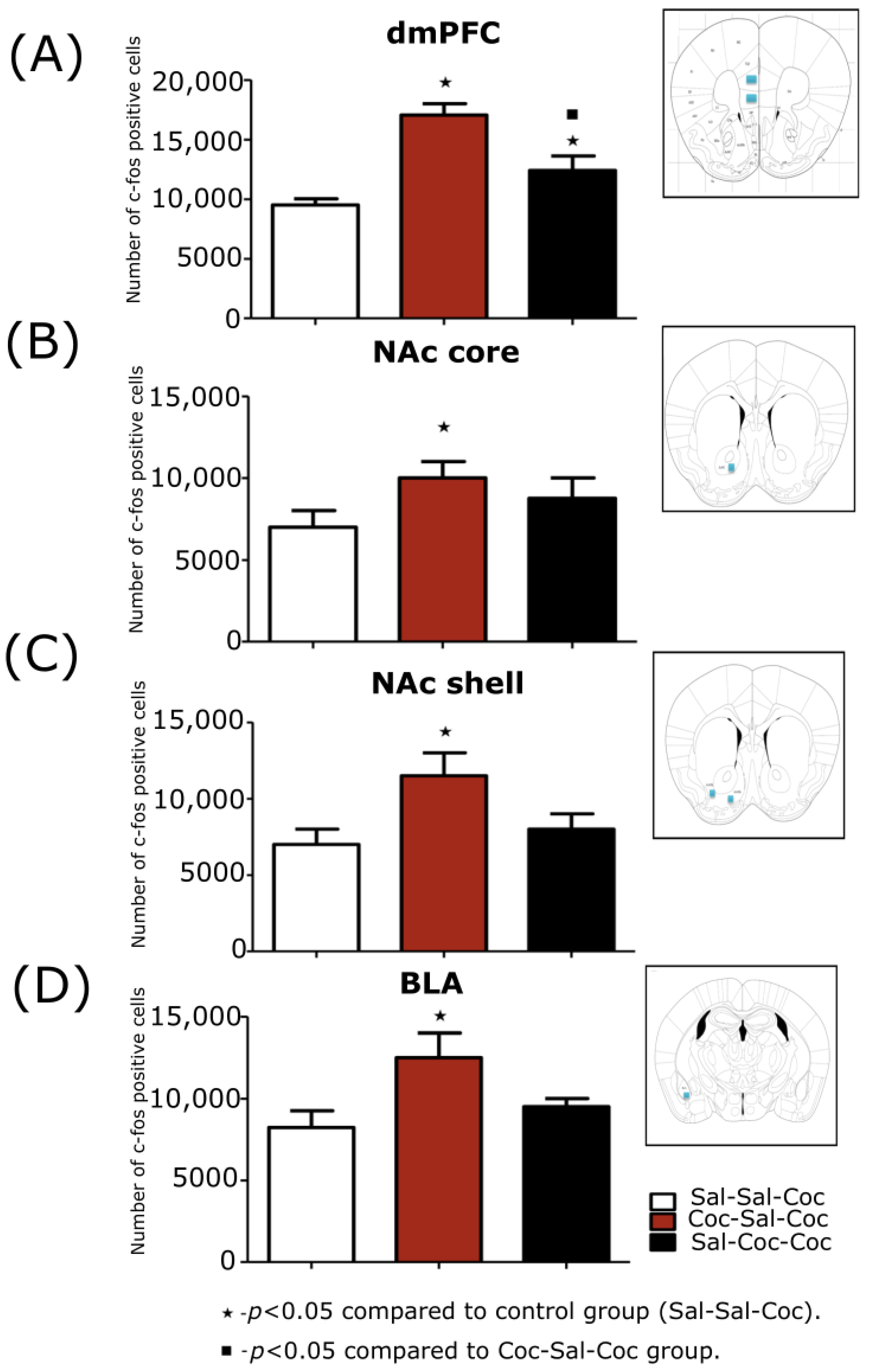
3.2.2. Stereological Evaluations of the Expression Phase of Cocaine-Induced Hyperlocomotion
dmPFC
NAc Core
NAc Shell
BLA
VTA
4. Discussion
5. Conclusions
6. Study limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalivas, P.W.; O’Brien, C. Drug Addiction as a Pathology of Staged Neuroplasticity. Neuropsychopharmacology 2008, 33, 166–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of Addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [Green Version]
- Feltenstein, M.W.; See, R.E.; Fuchs, R.A. Neural Substrates and Circuits of Drug Addiction. Cold Spring Harb. Perspect. Med. 2021, 11, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Volkow, N.D. Neurobiology of Addiction: A Neurocircuitry Analysis. Lancet Psychiatry 2016, 3, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, J.H.; Yun, K.; Kim, J.H. Alterations in Striatal Circuits Underlying Addiction-like Behaviors. Mol. Cells 2017, 40, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Alcaro, A.; Huber, R.; Panksepp, J. Behavioral Functions of the Mesolimbic Dopaminergic System: An Affective Neuroethological Perspective. Brain Res. Rev. 2007, 56, 283–321. [Google Scholar] [CrossRef] [Green Version]
- Solís, O.; García-Sanz, P.; Martín, A.B.; Granado, N.; Sanz-Magro, A.; Podlesniy, P.; Trullas, R.; Murer, M.G.; Maldonado, R.; Moratalla, R. Behavioral Sensitization and Cellular Responses to Psychostimulants Are Reduced in D2R Knockout Mice. Addict. Biol. 2021, 26, e12840. [Google Scholar] [CrossRef]
- Costa, F.G.; Frussa-Filho, R.; Canteras, N.S.; Valera, A.G.; Felicio, L.F. Blockade of Neurotensin Receptors during Amphetamine Discontinuation Indicates Individual Variability. Neuropeptides 2007, 41, 83–91. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. The Neural Basis of Drug Craving: An Incentive-Sensitization Theory of Addiction. Brain Res. Rev. 1993, 18, 247–291. [Google Scholar] [CrossRef]
- Vezina, P. Sensitization, Drug Addiction and Psychopathology in Animals and Humans. Prog. Neuropsychopharmacol. Biol. Psychiatry 2007, 31, 1553–1555. [Google Scholar] [CrossRef]
- Reeves, R.; Thiruchelvam, M.; Cory-Slechta, D.A. Development of Behavioral Sensitization to the Cocaine-like Fungicide Triadimefon Is Prevented by AMPA, NMDa, DA D1 but Not DA D2 Receptor Antagonists. Toxicol. Sci. 2004, 79, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Kalivas, P.; Sorg, B.; Hooks, M. The Pharmacology and Neural Circuitry of Sensitization to Psychostimulants. Behav. Pharmacol. 1993, 4, 315–334. [Google Scholar] [CrossRef] [PubMed]
- McDougall, S.A.; Rudberg, K.N.; Veliz, A.; Dhargalkar, J.M.; Garcia, A.S.; Romero, L.C.; Gonzalez, A.E.; Mohd-Yusof, A.; Crawford, C.A. Importance of D1 and D2 Receptor Stimulation for the Induction and Expression of Cocaine-Induced Behavioral Sensitization in Preweanling Rats. Behav. Brain Res. 2017, 326, 226–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinen, C.C.; Faria, R.R.; Frussa-Filho, R. Characterization of the Rapid-Onset Type of Behavioral Sensitization to Amphetamine in Mice: Role of Drug-Environment Conditioning. Neuropsychopharmacology 2006, 31, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Marinho, E.A.V.; Oliveira-Lima, A.J.; Santos, R.; Hollais, A.W.; Baldaia, M.A.; Wuo-Silva, R.; Yokoyama, T.S.; Takatsu-Coleman, A.L.; Patti, C.L.; Longo, B.M.; et al. Effects of Rimonabant on the Development of Single Dose-Induced Behavioral Sensitization to Ethanol, Morphine and Cocaine in Mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2015, 58, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Frussa-Filho, R.; Moraes Gonçalves, M.T.; Andersen, M.L.; De Araujo, N.P.; Chinen, C.C.; Tufik, S. Paradoxical Sleep Deprivation Potentiates Amphetamine-Induced Behavioural Sensitization by Increasing Its Conditioned Component. Brain Res. 2004, 1003, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Berro, L.F.; Hollais, A.W.; Patti, C.L.; Fukushiro, D.F.; Mári-Kawamoto, E.; Talhati, F.; Costa, J.M.; Zanin, K.A.; Lopes-Silva, L.B.; Ceccon, L.M.; et al. Sleep Deprivation Impairs the Extinction of Cocaine-Induced Environmental Conditioning in Mice. Pharmacol. Biochem. Behav. 2014, 124, 13–18. [Google Scholar] [CrossRef]
- Oliveira-Lima, A.J.; Santos, R.; Hollais, A.W.; Gerardi-Junior, C.A.; Baldaia, M.A.; Wuo-Silva, R.; Yokoyama, T.S.; Costa, J.L.; Malpezzi-Marinho, E.L.A.; Ribeiro-Barbosa, P.C.; et al. Effects of Ayahuasca on the Development of Ethanol-Induced Behavioral Sensitization and on a Post-Sensitization Treatment in Mice. Physiol. Behav. 2015, 142, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Childress, A.; Mozley, P.; McElgin, W.; Fitzgerald, J.; Reivich, M.; O’Brien, C. Limbic Activation During Cue-Induced Cocaine Craving. Am. J. Psychiatry 1999, 156, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Bradberry, C.W. Cocaine Sensitization and Dopamine Mediation of Cue Effects in Rodents, Monkeys, and Humans: Areas of Agreement, Disagreement, and Implications for Addiction. Psychopharmacology 2007, 191, 705–717. [Google Scholar] [CrossRef]
- Steketee, J.D.; Kalivas, P.W. Drug Wanting: Behavioral Sensitization and Relapse to Drug-Seeking Behavior. Pharmacol. Rev. 2011, 63, 348–365. [Google Scholar] [CrossRef] [PubMed]
- Berglind, W.J.; Whitfield, T.W.; Lalumiere, R.T.; Kalivas, P.W.; Mcginty, J.F. A Single Intra-PFC Infusion of BDNF Prevents Cocaineinduced Alterations in Extracellular Glutamate within the Nucleus Accumbens. J. Neurosci. 2009, 29, 3715–3719. [Google Scholar] [CrossRef] [Green Version]
- Andersen, S.L.; Teicher, M.H. Sex Differences in Dopamine Receptors and Their Relevance to ADHD. Neurosci. Biobehav. Rev. 2000, 24, 137–141. [Google Scholar] [CrossRef]
- Sneddon, E.A.; Ramsey, O.R.; Thomas, A.; Radke, A.K. Increased Responding for Alcohol and Resistance to Aversion in Female Mice. Alcohol. Clin. Exp. Res. 2020, 44, 1400–1409. [Google Scholar] [CrossRef]
- Reichel, C.M.; Chan, C.H.; Ghee, S.M.; See, R.E. Sex Differences in Escalation of Methamphetamine Self-Administration: Cognitive and Motivational Consequences in Rats. Psychopharmacology 2012, 223, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Monroe, S.; Radke, A.K. Aversion-Resistant Fentanyl Self-Administration in Mice. Psychopharmacology 2021, 238, 699–710. [Google Scholar] [CrossRef]
- Clayton, J.; Collins, F. NIH to Balance Sex in Cell and Animal Studies. Nature 2014, 509, 282–283. [Google Scholar] [CrossRef] [Green Version]
- Oliveira-Lima, A.J.; Marinho, E.A.V.; Santos-Baldaia, R.; Hollais, A.W.; Baldaia, M.A.; Talhati, F.; Ribeiro, L.T.; Wuo-Silva, R.; Berro, L.F.; Frussa-Filho, R. Context-Dependent Efficacy of a Counter-Conditioning Strategy with Atypical Neuroleptic Drugs in Mice Previously Sensitized to Cocaine. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 73, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Puntiverio, M.; Lerdrup, L.S.; Arvastson, L.; Aznar, S.; Andreasen, J.T. ADHD Medication and the Inverted U-Shaped Curve: A Pharmacological Study in Female Mice Performing the Rodent Continuous Performance Test (RCPT). Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 99, 109823. [Google Scholar] [CrossRef] [PubMed]
- Fukushiro, D.F.; Carvalho, R.d.C.; Ricardo, V.P.; Alvarez, J.d.N.; Ribeiro, L.T.C.; Frussa-Filho, R. Haloperidol (but Not Ziprasidone) Withdrawal Potentiates Sensitization to the Hyperlocomotor Effect of Cocaine in Mice. Brain Res. Bull. 2008, 77, 124–128. [Google Scholar] [CrossRef]
- Wuo-Silva, R.; Fukushiro-Lopes, D.F.; Fialho, B.P.; Hollais, A.W.; Santos-Baldaia, R.; Marinho, E.A.; Mári-Kawamoto, E.; Yokoyama, T.S.; Lopes-Silva, L.B.; Berro, L.F.; et al. Participation of Dopamine D1 and D2 receptors in the rapid-onset behavioral sensitization to modafinil. Front. Pharmacol. 2019, 10, 211. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Franklin, K. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Horger, B.A.; Iyasere, C.A.; Berhow, M.T.; Messer, C.J.; Nestler, E.J.; Taylor, J.R. Enhancement of Locomotor Activity and Conditioned Reward to Cocaine by Brain-Derived Neurotrophic Factor. J. Neurosci. 1999, 19, 4110–4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, P.J.; Meshul, C.K.; Phillips, T.J. Ethanol- and Cocaine-Induced Locomotion Are Genetically Related to Increases in Accumbal Dopamine. Genes Brain Behav. 2009, 8, 346–355. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, J.d.N.; Fukushiro, D.F.; Tatsu, J.A.O.; de Carvalho, E.P.; Gandolfi, A.C.d.C.; Tsuchiya, J.B.; Carrara-Nascimento, P.F.; Lima, M.L.; Bellot, R.G.; Frussa-Filho, R. Amphetamine-Induced Rapid-Onset Sensitization: Role of Novelty, Conditioning and Behavioral Parameters. Pharmacol. Biochem. Behav. 2006, 83, 500–507. [Google Scholar] [CrossRef]
- Fukushiro, D.F.; Josino, F.S.; Saito, L.P.; Berro, L.F.; Morgado, F.; Frussa-Filho, R. Acute and Chronic Ethanol Differentially Modify the Emotional Significance of a Novel Environment: Implications for Addiction. Int. J. Neuropsychopharmacol. 2012, 15, 1109–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koob, G.F.; Stinus, L.; Moal, M. Le Hyperactivity and Hypoactivity Produced by Lesions to the Mesolimbic Dopamine System. Behav. Brain Res. 1981, 3, 341–359. [Google Scholar] [CrossRef]
- Ford, C.P. The Role of D2-Autoreceptors in Regulating Dopamine Neuron Activity and Transmission. Neuroscience 2014, 282, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Kalivas, P.W.; Weber, B. Amphetamine Injection into the Ventral Mesencephalon Sensitizes Rats to Peripheral Amphetamine and Cocaine. J. Pharmacol. Exp. Ther. 1988, 245, 1095–1102. [Google Scholar]
- Thomas, M.J.; Kalivas, P.W.; Shaham, Y. Neuroplasticity in the Mesolimbic Dopamine System and Cocaine Addiction. Br. J. Pharmacol. 2008, 154, 327–342. [Google Scholar] [CrossRef] [Green Version]
- Mameli, M.; Balland, B.; Luján, R.; Lüscher, C. Rapid Synthesis and Synaptic Insertion of GluR2 for MGluR-LTD in the Ventral Tegmental Area. Science 2007, 317, 530–533. [Google Scholar] [CrossRef] [Green Version]
- Kalivas, P.W. The Glutamate Homeostasis Hypothesis of Addiction. Nat. Rev. Neurosci. 2009, 10, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Good, C.H.; Diaz-Ruiz, O.; Zhang, Y.; Hoffman, A.F.; Shan, L.; Kuang, S.Y.; Malik, N.; Chefer, V.I.; Tomac, A.C.; et al. NMDA Receptors on Non-Dopaminergic Neurons in the VTA Support Cocaine Sensitization. PLoS ONE 2010, 5, e12141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrario, C.R.; Li, X.; Wang, X.; Reimers, J.M.; Uejima, J.L.; Wolf, M.E. The Role of Glutamate Receptor Redistribution in Locomotor Sensitization to Cocaine. Neuropsychopharmacology 2010, 35, 818–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todtenkopf, M.S.; Stellar, J.R.; Melloni, R.H. Neither Ibotenic Acid nor Volkensin Lesions of the Nucleus Accumbens Shell Affect the Expression of Cocaine Sensitization. Eur. J. Neurosci. 2002, 16, 541–546. [Google Scholar] [CrossRef]
- Bossert, J.M.; Poles, G.C.; Wihbey, K.A.; Koya, E.; Shaham, Y. Differential Effects of Blockade of Dopamine D1-Family Receptors in Nucleus Accumbens Core or Shell on Reinstatement of Heroin Seeking Induced by Contextual and Discrete Cues. J. Neurosci. 2007, 27, 12655–12663. [Google Scholar] [CrossRef] [Green Version]
- Neisewander, J.L.; Baker, D.A.; Fuchs, R.A.; Tran-Nguyen, L.T.L.; Palmer, A.; Marshall, J.F. Fos Protein Expression and Cocaine-Seeking Behavior in Rats after Exposure to a Cocaine Self-Administration Environment. J. Neurosci. 2000, 20, 798–805. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, R.A.; Evans, K.A.; Ledford, C.C.; Parker, M.P.; Case, J.M.; Mehta, R.H.; See, R.E. The Role of the Dorsomedial Prefrontal Cortex, Basolateral Amygdala, and Dorsal Hippocampus in Contextual Reinstatement of Cocaine Seeking in Rats. Neuropsychopharmacology 2005, 30, 296–309. [Google Scholar] [CrossRef]
- Vanderschuren, L.J.M.J.; Kalivas, P.W. Alterations in Dopaminergic and Glutamatergic Transmission in the Induction and Expression of Behavioral Sensitization: A Critical Review of Preclinical Studies. Psychopharmacology 2000, 151, 99–120. [Google Scholar] [CrossRef]
- Pierce, R.C.; Kumaresan, V. The Mesolimbic Dopamine System: The Final Common Pathway for the Reinforcing Effect of Drugs of Abuse? Neurosci. Biobehav. Rev. 2006, 30, 215–238. [Google Scholar] [CrossRef]
- Kalivas, P.W. Neurocircuitry of Addiction. In Neuropsychopharmacology, The Fifth Generation of Progress; Davis, K.L., Charney, D., Coyle, J.T., Nemenorff, C., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2002; pp. 1357–1366. [Google Scholar]
- Ambroggi, F.; Ishikawa, A.; Fields, H.L.; Nicola, S.M. Basolateral Amygdala Neurons Facilitate Reward-Seeking Behavior by Exciting Nucleus Accumbens Neurons. Neuron 2008, 59, 648–661. [Google Scholar] [CrossRef] [Green Version]
- Gregoriou, G.C.; Kissiwaa, S.A.; Patel, S.D.; Bagley, E.E. Dopamine and Opioids Inhibit Synaptic Outputs of the Main Island of the Intercalated Neurons of the Amygdala. Eur. J. Neurosci. 2019, 50, 2065–2074. [Google Scholar] [CrossRef] [PubMed]
- Hotsenpiller, G.; Horak, B.T.; Wolf, M.E. Dissociation of Conditioned Locomotion and Fos Induction in Response to Stimuli Formerly Paired with Cocaine. Behav. Neurosci. 2002, 116, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, P.W.; Funk, D.; Juzytsch, W.; Li, Z.; Lê, A.D. Effects of Opioid Receptor Blockade on the Renewal of Alcohol Seeking Induced by Context: Relationship to c-Fos MRNA Expression. Eur. J. Neurosci. 2007, 26, 2815–2823. [Google Scholar] [CrossRef] [PubMed]
- Meil, W.M.; See, R.E. Lesions of the Basolateral Amygdala Abolish the Ability of Drug Associated Cues to Reinstate Responding during Withdrawal from Self- Administered Cocaine. Behav. Brain Res. 1997, 87, 139–148. [Google Scholar] [CrossRef]
- Volkow, N.D.; Fowler, J.S. Addiction, a Disease of Compulsion and Drive: Involvement of the Orbitofrontal Cortex. Cereb. Cortex 2000, 10, 318–325. [Google Scholar] [CrossRef]
- Jiang, Q.; Wang, C.M.; Fibuch, E.E.; Wang, J.Q.; Chu, X.P. Differential Regulation of Locomotor Activity to Acute and Chronic Cocaine Administration by Acid-Sensing Ion Channel 1a and 2 in Adult Mice. Neuroscience 2013, 246, 170–178. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment 1 (Induction Phase) | |||
|---|---|---|---|
| Groups | Injection in the open-field (days 1, 3, 5, 7, 9, 11, 13, 15) Behavioral test (days 1 and 15) | Injection in the home-cage (days 1, 3, 5, 7, 9, 11, 13, 15) Behavioral test (days 1 and 15) | |
| Sal-Sal | Saline | Saline | |
| Coc-Sal | Cocaine | Saline | |
| Sal-Coc | Saline | Cocaine | |
| Experiment 2 (Expression Phase) | |||
| Groups | Injection in the open-field (days 1, 3, 5, 7, 9, 11, 13, 15) Behavioral test (days 1 and 15) | Injection in the home-cage (days 1, 3, 5, 7, 9, 11, 13, 15) Behavioral test (days 1 and 15) | Cocaine challenge and behavioral test in the open-field (day 26) |
| Sal-Sal-Coc | Saline | Saline | Cocaine |
| Coc-Sal-Coc | Cocaine | Saline | Cocaine |
| Sal-Coc-Coc | Saline | Cocaine | Cocaine |
| Structure | Frame (µm × µm) | Grid (µm × µm) | Disector Height (µm) | Guard Zone (µm) |
|---|---|---|---|---|
| Dorsomedial prefrontal cortex | 50 × 50 | 200 × 200 | 12 | 3 |
| Nucleus accumbens core | 100 × 67 | 175 × 175 | 12 | 3 |
| Nucleus accumbens shell | 100 × 67 | 175 × 175 | 12 | 3 |
| Basolateral amygdala | 98 × 58 | 150 × 150 | 12 | 3 |
| Ventral tegmental area | 60 × 60 | 150 × 150 | 12 | 3 |
| Structures | Induction phase | Expression phase | ||
|---|---|---|---|---|
| Coc-Sal | Sal-Coc | Coc-Sal-Coc | Sal-Coc-Coc | |
| dmPFC | ↑ | = | ↑ | ↑↑ |
| NAc core | ↑ | = | ↑ | = |
| NAc shell | = | = | ↑ | = |
| BLA | ↑ | = | ↑ | = |
| VTA | ↑ | ↑↑ | = | = |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Baldaia, R.d.; Wuo-Silva, R.; Sanabria, V.; Baldaia, M.A.; Yokoyama, T.S.; Coppi, A.A.; Hollais, A.W.; Marinho, E.A.V.; Oliveira-Lima, A.J.; Longo, B.M. Distinctive Neuroanatomic Regions Involved in Cocaine-Induced Behavioral Sensitization in Mice. Biomedicines 2023, 11, 383. https://doi.org/10.3390/biomedicines11020383
Santos-Baldaia Rd, Wuo-Silva R, Sanabria V, Baldaia MA, Yokoyama TS, Coppi AA, Hollais AW, Marinho EAV, Oliveira-Lima AJ, Longo BM. Distinctive Neuroanatomic Regions Involved in Cocaine-Induced Behavioral Sensitization in Mice. Biomedicines. 2023; 11(2):383. https://doi.org/10.3390/biomedicines11020383
Chicago/Turabian StyleSantos-Baldaia, Renan dos, Raphael Wuo-Silva, Viviam Sanabria, Marilia A. Baldaia, Thais S. Yokoyama, Antonio Augusto Coppi, André W. Hollais, Eduardo A. V. Marinho, Alexandre J. Oliveira-Lima, and Beatriz M. Longo. 2023. "Distinctive Neuroanatomic Regions Involved in Cocaine-Induced Behavioral Sensitization in Mice" Biomedicines 11, no. 2: 383. https://doi.org/10.3390/biomedicines11020383