
Oxidative Stress in the Anterior Ocular Diseases: Diagnostic and Treatment
 ,
,  , , and
, , and
Abstract
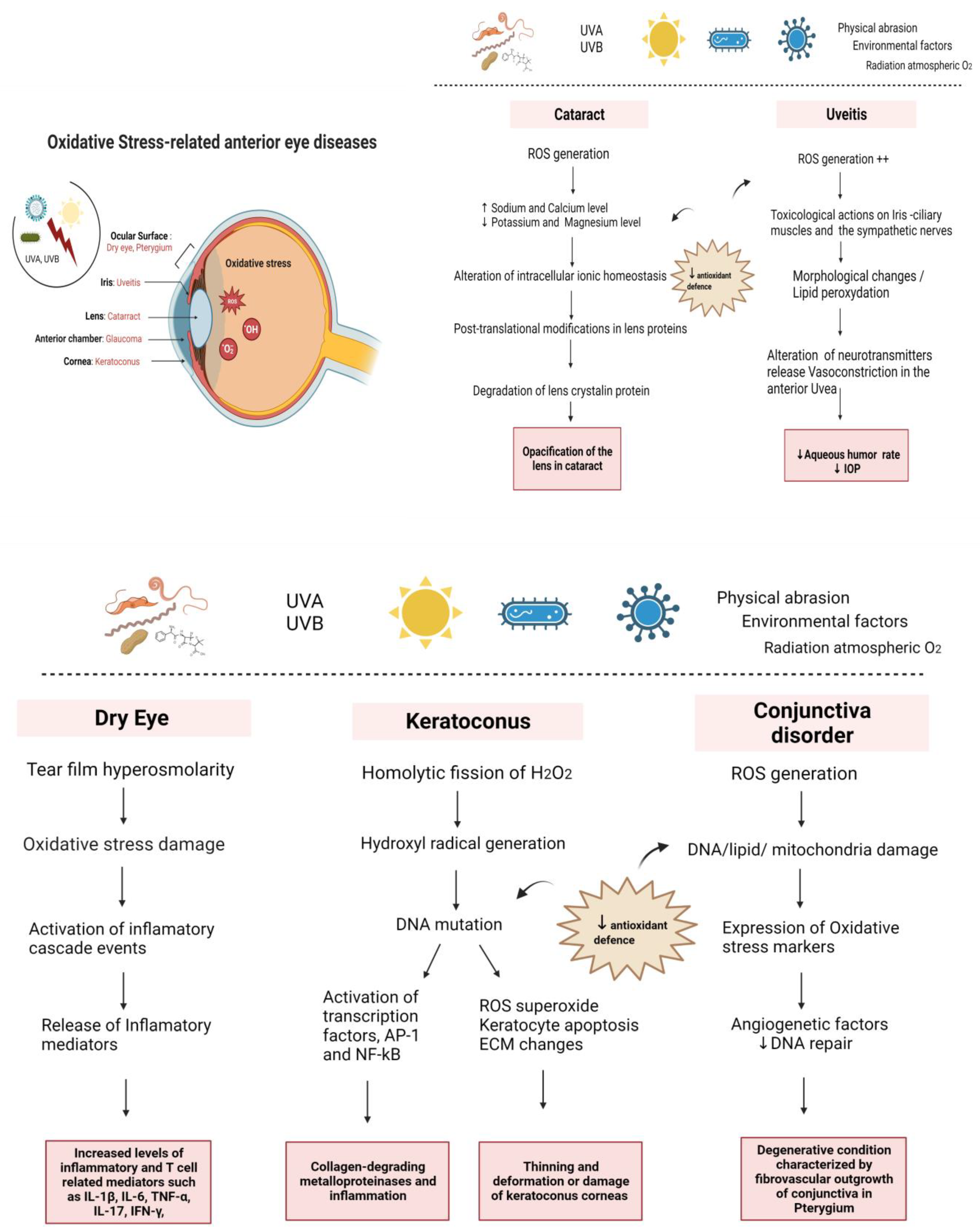
:1. Introduction
1.1. Oxidative Stress in the Anterior Segment of the Eye
1.2. Main Biomarkers of Oxidative Stress and Inflammation
2. Mechanism of Oxidative Stress in Anterior Eye Disorders
2.1. Oxidative Stress in the Ocular Surface
2.1.1. Dry Eye
2.1.2. Corneal Pathologies
2.1.3. Conjunctival Pathologies
2.2. Oxidative Stress in the Anterior Chamber
2.3. Oxidative Stress in the Iris
2.4. Oxidative Stress in the Lens
3. Diagnostic
3.1. Dry Eye
3.2. Keratoconus
3.3. Conjunctiva
3.4. Anterior Uveitis
3.5. Cataracts
4. Treatment
4.1. Dry Eye
4.2. Keratoconus
4.3. Conjunctiva
4.4. Anterior Uveitis
4.5. Cataracts
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perez-Garmendia, R.; Rodriguez, A.L.D.E.; Ramos-Martinez, I.; Zuñiga, N.M.; Gonzalez-Salinas, R.; Quiroz-Mercado, H.; Zenteno, E.; Hernández, E.R.; Hernández-Zimbrón, L.F. Interplay between oxidative stress, inflammation, and amyloidosis in the anterior segment of the eye; its pathological implications. Oxidative Med. Cell. Longev. 2020, 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Oduntan, O.; Masige, K.P. A review of the role of oxidative stress in the pathogenesis of eye diseases. Afr. Vis. Eye Health 2011, 70, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. IJBS 2008, 4, 89. [Google Scholar]
- Ahmad, A.; Ahsan, H. Biomarkers of inflammation and oxidative stress in ophthalmic disorders. J. Immunoass. Immunochem. 2020, 41, 257–271. [Google Scholar] [CrossRef]
- Shoham, A.; Hadziahmetovic, M.; Dunaief, J.L.; Mydlarski, M.B.; Schipper, H.M. Oxidative stress in diseases of the human cornea. Free Radic. Biol. Med. 2008, 45, 1047–1055. [Google Scholar] [CrossRef]
- Chen, Y.; Mehta, G.; Vasiliou, V. Antioxidant defenses in the ocular surface. Ocul. Surf. 2009, 7, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Sacca, S.C.; Roszkowska, A.M.; Izzotti, A. Environmental light and endogenous antioxidants as the main determinants of non-cancer ocular diseases. Mutat. Res./Rev. Mutat. Res. 2013, 752, 153–171. [Google Scholar] [CrossRef]
- Saccà, S.C.; Cutolo, C.A.; Ferrari, D.; Corazza, P.; Traverso, C.E. The eye, oxidative damage and polyunsaturated fatty acids. Nutrients 2018, 10, 668. [Google Scholar] [CrossRef] [Green Version]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2015, 15, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Čejková, J.; Vejražka, M.; Pláteník, J.; Štípek, S. Age-related changes in superoxide dismutase, glutathione peroxidase, catalase and xanthine oxidoreductase/xanthine oxidase activities in the rabbit cornea. Exp. Gerontol. 2004, 39, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K. Does the interdependence between oxidative stress and inflammation explain the antioxidant paradox? Oxidative Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative stress and inflammation: What polyphenols can do for us? Oxidative Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Perry, H.D. Dry eye disease: Pathophysiology, classification, and diagnosis. Am. J. Manag. Care 2008, 14 (Suppl. S3), S79–S87. [Google Scholar]
- Seen, S.; Tong, L. Dry eye disease and oxidative stress. Acta Ophthalmol. 2018, 96, e412–e420. [Google Scholar] [CrossRef] [Green Version]
- Clayton, J.A. Dry Eye. N. Engl. J. Med. 2018, 378, 2212–2223. [Google Scholar] [CrossRef]
- Alves, M.; Novaes, P.; Morraye, M.D.A.; Reinach, P.S.; Rocha, E.M. Is dry eye an environmental disease? Arq. Bras. Oftalmol. 2014, 77, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Jie, Y.; Xu, L.; Wu, Y.Y.; Jonas, J.B. Prevalence of dry eye among adult Chinese in the Beijing Eye Study. Eye 2009, 23, 688–693. [Google Scholar] [CrossRef] [Green Version]
- Junqueira, V.B.; Barrros, S.B.M.; Cham, S.S.; Rodrigues, L.; Giavarotti, L.; Abud, R.L.; Deucher, G.P. Aging and oxidative stress. Mol. Asp. Med. 2004, 25, 5–16. [Google Scholar] [CrossRef]
- Kruk, J.; Kubasik-Kladna, K.; Aboul-Enein, H.Y. The Role Oxidative Stress in the Pathogenesis of Eye Diseases: Current Status and a Dual Role of Physical Activity. Mini Rev. Med. Chem. 2015, 16, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Cheeseman, K.H.; Slater, T.F. An introduction to free radical biochemistry. Br. Med. Bull. 1993, 49, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar] [CrossRef]
- Deng, R.; Hua, X.; Li, J.; Chi, W.; Zhang, Z.; Lu, F.; Zhang, L.; Pflugfelder, S.C.; Li, D.-Q. Oxidative stress markers induced by hyperosmolarity in primary human corneal epithelial cells. PLoS ONE 2015, 10, e0126561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The definition and classification of dry eye disease: Report of the Definition and Classification Subcommittee of the International Dry Eye WorkShop (2007). Ocul. Surf. 2007, 5, 75–92. [CrossRef]
- Nakamura, S.; Shibuya, M.; Nakashima, H.; Hisamura, R.; Masuda, N.; Imagawa, T.; Uehara, M.; Tsubota, K. Involvement of oxidative stress on corneal epithelial alterations in a blink-suppressed dry eye. Investig. Opthalmol. Vis. Sci. 2007, 48, 1552–1558. [Google Scholar] [CrossRef]
- Enríquez-de-Salamanca, A.; Castellanos, E.; Stern, M.E.; Fernández, I.; Carreño, E.; García-Vázquez, C.; Herreras, J.M.; Calonge, M. Tear cytokine and chemokine analysis and clinical correlations in evaporative-type dry eye disease. Mol. Vis. 2010, 16, 862–873. [Google Scholar]
- Pinazo-Durán, M.D.; Galbis-Estrada, C.; Pons-Vázquez, S.; Cantú-Dibildox, J.; Marco-Ramírez, C.; Benítez-del-Castillo, J. Effects of a nutraceutical formulation based on the combination of antioxidants and ω-3 essential fatty acids in the expression of inflammation and immune response mediators in tears from patients with dry eye disorders. Clin. Interv. Aging 2013, 8, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Galbis-Estrada, C.; Cantu-Dibildox, J.; Galbis-Estrada, C.; Marco-Ramirez, C.; Diaz-Llopis, M.; Benitez-Del-Castillo, J. Patients undergoing long-term treatment with antihypertensive eye drops responded positively with respect to their ocular surface disorder to oral supplementation with antioxidants and essential fatty acids. Clin. Interv. Aging 2013, 8, 711–719. [Google Scholar]
- Kojima, T.; Wakamatsu, T.H.; Dogru, M.; Ogawa, Y.; Igarashi, A.; Ibrahim, O.M.A.; Inaba, T.; Shimizu, T.; Noda, S.; Obata, H.; et al. Age-related dysfunction of the lacrimal gland and oxidative stress: Evidence from the Cu,Zn-superoxide dismutase-1 (Sod1) knockout mice. Am. J. Pathol. 2012, 180, 1879–1896. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, Y.; Dogru, M.; Tsubota, K. Laboratory findings in tear fluid analysis. Clin. Chim. Acta 2006, 369, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Uchino, Y.; Kawakita, T.; Miyazawa, M.; Ishii, T.; Onouchi, H.; Yasuda, K.; Ogawa, Y.; Shimmura, S.; Ishii, N.; Tsubota, K. Oxidative stress induced inflammation initiates functional decline of tear production. PLoS ONE 2012, 7, e45805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, T.M.; Tomiyoshi, L.M.; Dias, A.C.; Roma, L.P.; Módulo, C.M.; Malki, L.T.; Filho, E.B.M.; Deminice, R.; Jordão, A.A.; Cunha, D.A.; et al. Age-dependent changes in rat lacrimal gland anti-oxidant and vesicular related protein expression profiles. Mol. Vis. 2012, 18, 194–202. [Google Scholar] [PubMed]
- Dogru, M.; Kojima, T.; Simsek, C.; Tsubota, K. Potential Role of Oxidative Stress in Ocular Surface Inflammation and Dry Eye Disease. Investig. Ophthalmol. Vis. Sci. 2018, 59, DES163–DES168. [Google Scholar] [CrossRef] [Green Version]
- Tangvarasittichai, O.; Tangvarasittichai, S. Oxidative Stress, Ocular Disease and Diabetes Retinopathy. Curr. Pharm. Des. 2018, 24, 4726–4741. [Google Scholar] [CrossRef]
- Leung, E.W.; Rife, L.; Smith, R.E.; Kay, E.P. Extracellular matrix components in retrocorneal fibrous membrane in comparison to corneal endothelium and Descemet’s membrane. Mol. Vis. 2000, 6, 15–23. [Google Scholar]
- Wenk, J.; Brenneisen, P.; Meewes, C.; Wlaschek, M.; Peters, T.; Blaudschun, R.; Ma, W.; Kuhr, L.; Schneider, L.; Scharffet-ter-Kochanek, K. UV-induced oxidative stress and photoaging. Curr. Probl. Dermatol. 2001, 29, 83–94. [Google Scholar]
- Choi, S.I.; Dadakhujaev, S.; Ryu, H.; Kim, T.I.; Kim, E.K. Melatonin protects against oxidative stress in granular corneal dystrophy type 2 corneal fibroblasts by mechanisms that involve membrane melatonin receptors. J. Pineal. Res. 2011, 51, 94–103. [Google Scholar] [CrossRef]
- Peng, C.Y.; Lan, C.-H.; Juang, Y.-J.; Tsao, T.-H.; Dai, Y.-T.; Liu, H.-H.; Chen, C.-J. Exposure assessment of aluminum arc welding radiation. Health Phys. 2007, 93, 298–306. [Google Scholar] [CrossRef] [Green Version]
- Sherwin, T.; Brookes, N.H. Morphological changes in keratoconus: Pathology or pathogenesis. Clin. Exp. Ophthalmol. 2004, 32, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, Y.S. Keratoconus. Surv. Ophthalmol. 1998, 42, 297–319. [Google Scholar] [CrossRef] [PubMed]
- Gulpamuk, B.; Koç, M.; Karatepe, M.S.; Yildiz, A.; Erel, O.; Neselioglu, S.; Yilmazbas, P. Novel Assay Assessment of Oxidative Stress Biomarkers in Patients with Keratoconus: Thiol-Disulfide Homeostasis. Curr. Eye Res. 2017, 42, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Toprak, I.; Kucukatay, V.; Yildirim, C.; Kilic-Toprak, E.; Kilic-Erkek, O. Increased systemic oxidative stress in patients with keratoconus. Eye 2014, 28, 285–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chwa, M.; Atilano, S.; Reddy, V.; Jordan, N.; Kim, D.W.; Kenney, M.C. Increased stress-induced generation of reactive oxygen species and apoptosis in human keratoconus fibroblasts. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1902–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chwa, M.; Atilano, S.R.; Hertzog, D.; Zheng, H.; Langberg, J.; Kim, D.W.; Kenney, M.C. Hypersensitive response to oxidative stress in keratoconus corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4361–4369. [Google Scholar] [CrossRef]
- Määttä, M.; Heljasvaara, R.; Sormunen, R.; Pihlajaniemi, T.; Autio-Harmainen, H.; Tervo, T. Differential expression of collagen types XVIII/endostatin and XV in normal, keratoconus, and scarred human corneas. Cornea 2006, 25, 341–349. [Google Scholar] [CrossRef]
- Brown, D.J.; Lin, B.; Chwa, M.; Atilano, S.R.; Kim, D.W.; Kenney, M.C. Elements of the nitric oxide pathway can degrade TIMP-1 and increase gelatinase activity. Mol. Vis. 2004, 10, 281–288. [Google Scholar] [PubMed]
- Nita, M.; Nita, M.; Strzałka-Mrozik, B.; Grzybowski, A.; Mazurek, U. Age-related macular degeneration and changes in the extracellular matrix. Med. Sci. Monit. 2014, 20, 1003–1016. [Google Scholar]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Smith, V.A.; Hoh, H.B.; Littleton, M.; Easty, D.L. Over-expression of a gelatinase A activity in keratoconus. Eye 1995, 9 Pt 4, 429–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surazynski, A.; Liu, Y.; Miltyk, W.; Phang, J.M. Nitric oxide regulates prolidase activity by serine/threonine phosphorylation. J. Cell. Biochem. 2005, 96, 1086–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palka, J.A.; Phang, J.M. Prolidase activity in fibroblasts is regulated by interaction of extracellular matrix with cell surface integrin receptors. J. Cell. Biochem. 1997, 67, 166–175. [Google Scholar] [CrossRef]
- Göncü, T.; Akal, A.; Adibelli, F.M.; Çakmak, S.; Sezen, H.; Ylmaz, F. Tear Film and Serum Prolidase Activity and Oxidative Stress in Patients with Keratoconus. Cornea 2015, 34, 1019–1023. [Google Scholar] [CrossRef]
- Balasubramanian, S.A.; Mohan, S.; Pye, D.C.; Willcox, M.D.P. Proteases, proteolysis and inflammatory molecules in the tears of people with keratoconus. Acta Ophthalmol. 2012, 90, e303–e309. [Google Scholar] [CrossRef]
- Kao, W.W.; Vergnes, J.-P.; Ebert, J.; Sundar-Raj, C.; Brown, S.I. Increased collagenase and gelatinase activities in keratoconus. Biochem. Biophys. Res. Commun. 1982, 107, 929–936. [Google Scholar] [CrossRef]
- Nichols, B.A. Conjunctiva. Microsc. Res. Tech. 1996, 33, 296–319. [Google Scholar] [CrossRef]
- Yokoi, N.; Inatomi, T.; Kinoshita, S. Surgery of the conjunctiva. Dev. Ophthalmol. 2008, 41, 138–158. [Google Scholar]
- de Paiva, C.S. Effects of Aging in Dry Eye. Int. Ophthalmol. Clin. 2017, 57, 47–64. [Google Scholar] [CrossRef] [Green Version]
- García-Posadas, L.; Hodges, R.R.; Li, D.; Shatos, M.A.; Storr-Paulsen, T.; Diebold, Y.; Dartt, D.A. Interaction of IFN-γ with cholinergic agonists to modulate rat and human goblet cell function. Mucosal. Immunol. 2016, 9, 206–217. [Google Scholar] [CrossRef] [Green Version]
- De Paiva, C.S.; Raince, J.K.; McClellan, A.J.; Shanmugam, K.P.; Pangelinan, S.B.; A Volpe, E.; Corrales, R.M.; Farley, W.J.; Corry, D.B.; Li, D.-Q.; et al. Homeostatic control of conjunctival mucosal goblet cells by NKT-derived IL-13. Mucosal Immunol. 2011, 4, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakamatsu, T.H.; Dogru, M.; Matsumoto, Y.; Kojima, T.; Kaido, M.; Ibrahim, O.M.A.; Sato, E.A.; Igarashi, A.; Ichihashi, Y.; Satake, Y.; et al. Evaluation of lipid oxidative stress status in Sjögren syndrome patients. Investig. Ophthalmol. Vis. Sci. 2013, 54, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cejková, J.; Ardan, T.; Simonova, Z.; Cejka, C.; Malec, J.; Dotrelová, D.; Brunová, B. Decreased expression of antioxidant enzymes in the conjunctival epithelium of dry eye (Sjögren’s syndrome) and its possible contribution to the development of ocular surface oxidative injuries. Histol. Histopathol. 2008, 23, 1477–1483. [Google Scholar] [PubMed]
- Malozhen, S.A.; Trufanov, S.V.; Krakhmaleva, D.A. Pterygium: Etiology, pathogenesis, treatment. Vestn. Oftalmol. 2017, 133, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Hovanesian, J.A.; Starr, C.E.; Vroman, D.T.; Mah, F.S.; Gomes, J.A.; Farid, M.; Shamie, N.; Davidson, R.S.; John, T.; Holland, E.J.; et al. Surgical techniques and adjuvants for the management of primary and recurrent pterygia. J. Cataract. Refract. Surg. 2017, 43, 405–419. [Google Scholar] [CrossRef]
- Mudhar, H.S. Update on conjunctival pathology. Indian J. Ophthalmol. 2017, 65, 797–807. [Google Scholar] [CrossRef]
- Austin, P.; Jakobiec, F.A.; Iwamoto, T. Elastodysplasia and elastodystrophy as the pathologic bases of ocular pterygia and pinguecula. Ophthalmology 1983, 90, 96–109. [Google Scholar] [CrossRef]
- Liu, T.; Liu, Y.; Xie, L.; He, X.; Bai, J. Progress in the pathogenesis of pterygium. Curr. Eye Res. 2013, 38, 1191–1197. [Google Scholar] [CrossRef]
- Beatty, S.; Koh, H.-H.; Phil, M.; Henson, D.; Boulton, M. The Role of Oxidative Stress in the Pathogenesis of Age-Related Macular Degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Kormanovski, A.; Parra, F.; Jarillo-Luna, A.; Lara-Padilla, E.; Pacheco-Yépez, J.; Campos-Rodriguez, R. Oxidant/antioxidant state in tissue of prymary and recurrent pterygium. BMC Ophthalmol. 2014, 14, 149. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.C.; Tsai, Y.-Y.; Bau, D.-T.; Cheng, Y.-W.; Tseng, S.-H.; Wang, R.-F.; Tsai, F.-J. Pterygium and genetic polymorphisms of the DNA repair enzymes XRCC1, XPA, and XPD. Mol. Vis. 2010, 16, 698–704. [Google Scholar]
- Bradley, J.C.; Yang, W.; Bradley, R.H.; Reid, T.W.; Schwab, I.R. The science of pterygia. Br. J. Ophthalmol. 2010, 94, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Anguria, P.; Kitinya, J.; Ntuli, S.; Carmichael, T. The role of heredity in pterygium development. Int. J. Ophthalmol. 2014, 7, 563–573. [Google Scholar] [PubMed]
- Crowston, J.G.; Weinreb, R.N. Glaucoma medication and aqueous humor dynamics. Curr. Opin. Ophthalmol. 2005, 16, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Namekata, K.; Guo, X.; Noro, T.; Harada, C.; Harada, T. Targeting Oxidative Stress for Treatment of Glaucoma and Optic Neuritis. Oxidative Med. Cell. Longev. 2017, 2017, 2817252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zheng, Y. Oxidative stress and antioxidants in the trabecular meshwork. PeerJ 2019, 7, e8121. [Google Scholar] [CrossRef]
- Babizhayev, M.A. Current ocular drug delivery challenges for N-acetylcarnosine: Novel patented routes and modes of delivery, design for enhancement of therapeutic activity and drug delivery relationships. Recent Pat. Drug Deliv. Formul. 2009, 3, 229–265. [Google Scholar] [CrossRef]
- Bill, A. Some aspects of aqueous humour drainage. Eye 1993, 7 Pt 1, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Ammar, D.A.; Hamweyah, K.M.; Kahook, M.Y. Antioxidants Protect Trabecular Meshwork Cells from Hydrogen Peroxide-Induced Cell Death. Transl. Vis. Sci. Technol. 2012, 1, 4. [Google Scholar] [CrossRef]
- Saccà, S.C.; Izzotti, A.; Rossi, P.; Traverso, C. Glaucomatous outflow pathway and oxidative stress. Exp. Eye Res. 2007, 84, 389–399. [Google Scholar] [CrossRef]
- Tian, B.; Geiger, B.; Epstein, D.L.; Kaufman, P.L. Cytoskeletal involvement in the regulation of aqueous humor outflow. Investig. Ophthalmol. Vis. Sci. 2000, 41, 619–623. [Google Scholar]
- Izzotti, A.; Sacca’, S.; Longobardi, M.; Cartiglia, C. Sensitivity of ocular anterior chamber tissues to oxidative damage and its relevance to the pathogenesis of glaucoma. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5251–5258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, S.M.; Lerner, S.; Brunzini, R.; Evelson, P.A.; Llesuy, S.F. Oxidative stress markers in aqueous humor of glaucoma patients. Am. J. Ophthalmol. 2004, 137, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Acott, T.S.; Kelley, M.J. Extracellular matrix in the trabecular meshwork. Exp. Eye Res. 2008, 86, 543–561. [Google Scholar] [CrossRef] [Green Version]
- Lütjen-Drecoll, E. Functional morphology of the trabecular meshwork in primate eyes. Prog. Retin. Eye Res. 1999, 18, 91–119. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Brodskaya, M.W. Fibronectin detection in drainage outflow system of human eyes in ageing and progression of open-angle glaucoma. Mech. Ageing Dev. 1989, 47, 145–157. [Google Scholar] [CrossRef]
- Tamm, E.R.; Fuchshofer, R. What increases outflow resistance in primary open-angle glaucoma? Surv. Ophthalmol. 2007, 52 (Suppl. S2), S101–S104. [Google Scholar] [CrossRef]
- Scott, P.A.; Lu, Z.; Liu, Y.; Gong, H. Relationships between increased aqueous outflow facility during washout with the changes in hydrodynamic pattern and morphology in bovine aqueous outflow pathways. Exp. Eye Res. 2009, 89, 942–949. [Google Scholar] [CrossRef]
- Tamm, E.R.; Russell, P.; Johnson, D.H.; Piatigorsky, J. Human and monkey trabecular meshwork accumulate alpha B-crystallin in response to heat shock and oxidative stress. Investig. Ophthalmol. Vis. Sci. 1996, 37, 2402–2413. [Google Scholar]
- Tektas, O.Y.; Lütjen-Drecoll, E. Structural changes of the trabecular meshwork in different kinds of glaucoma. Exp. Eye Res. 2009, 88, 769–775. [Google Scholar] [CrossRef]
- Kahn, M.G.; Giblin, F.J.; Epstein, D.L. Glutathione in calf trabecular meshwork and its relation to aqueous humor outflow facility. Investig. Ophthalmol. Vis. Sci 1983, 24, 1283–1287. [Google Scholar]
- Zhao, J.; Giblin, F.J.; Epstein, D.L. Oxidative stress in the trabecular meshwork (Review). Int. J. Mol. Med. 2016, 38, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Leung, K.W.; Zhang, Y.-H.; Duan, S.; Zhong, X.-F.; Jiang, R.-Z.; Peng, Z.; Tombran-Tink, J.; Ge, J. Mitochondrial complex I defect induces ROS release and degeneration in trabecular meshwork cells of POAG patients: Protection by antioxidants. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1447–1458. [Google Scholar] [CrossRef] [Green Version]
- Pokrovskaya, O.; O’Brien, C. What’s in a Gene? Pseudoexfoliation Syndrome and Pigment Dispersion Syndrome in the Same Patient. Case Rep. Ophthalmol. 2016, 7, 54–60. [Google Scholar] [CrossRef]
- Mastronikolis, S.; Pagkalou, M.; Plotas, P.; Kagkelaris, K.; Georgakopoulos, C.D. Emerging roles of oxidative stress in the pathogenesis of pseudoexfoliation syndrome (Review). Exp. Ther. Med. 2022, 24, 602. [Google Scholar] [CrossRef]
- Benoist d’Azy, C.; Pereira, B.; Chiambaretta, F.; Dutheil, F. Oxidative and Anti-Oxidative Stress Markers in Chronic Glaucoma: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0166915. [Google Scholar] [CrossRef] [PubMed]
- Shirakami, T.; Yamanaka, M.; Fujihara, J.; Matsuoka, Y.; Gohto, Y.; Obana, A.; Tanito, M. Advanced Glycation End Product Accumulation in Subjects with Open-Angle Glaucoma with and without Exfoliation. Antioxidants 2020, 9, 755. [Google Scholar] [CrossRef] [PubMed]
- Zeppieri, M. Pigment dispersion syndrome: A brief overview. J. Clin. Transl. Res. 2022, 8, 344–350. [Google Scholar]
- Mermoud, A. Physiopathology of uveitic glaucoma. Klin. Monbl. Augenheilkd. 1997, 210, 269–273. [Google Scholar] [CrossRef]
- Moorthy, R.S.; Mermoud, A.; Baerveldt, G.; Minckler, D.S.; Lee, P.P.; Rao, N.A. Glaucoma associated with uveitis. Surv. Ophthalmol. 1997, 41, 361–394. [Google Scholar] [CrossRef]
- Dumbrăveanu, L.; Cușnir, V.; Bobescu, D. A review of neovascular glaucoma. Etiopathogenesis and treatment. Rom. J. Ophthalmol. 2021, 65, 315–329. [Google Scholar] [PubMed]
- Barac, I.R.; Pop, M.D.; Gheorghe, A.I.; Taban, C. Neovascular secondary glaucoma, etiology and pathogenesis. Rom. J. Ophthalmol. 2015, 59, 24–28. [Google Scholar]
- Sun, C.; Zhang, H.; Tang, Y.; Chen, Y.; Li, Y.; Nie, C.; Gu, J.; Luo, L.; Wang, Z. Aqueous Inflammation and Ischemia-Related Biomarkers in Neovascular Glaucoma with Stable Iris Neovascularization. Curr. Eye Res. 2020, 45, 1504–1513. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G. Oxidative stress in glaucomatous neurodegeneration: Mechanisms and consequences. Prog. Retin. Eye Res. 2006, 25, 490–513. [Google Scholar] [CrossRef]
- Matthews, A.G. The lens and cataracts. Vet. Clin. N. Am. Equine. Pract. 2004, 20, 393–415. [Google Scholar] [CrossRef]
- Lee, J.; Giordano, S.; Zhang, J. Autophagy, mitochondria and oxidative stress: Cross-talk and redox signalling. Biochem. J 2012, 441, 523–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.C.; Richer, S.; Bode, A.M. Ocular oxidants and antioxidant protection. Proc. Soc. Exp. Biol. Med. 1998, 217, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Ohia, S.E.; Opere, C.A.; Leday, A.M. Pharmacological consequences of oxidative stress in ocular tissues. Mutat. Res. 2005, 579, 22–36. [Google Scholar] [CrossRef]
- Opere, C.; Tang, L.; Imler, M.; Kim, J.; Okoye, M.; Ohia, S. Regulation of uveal sympathetic neurotransmission by peroxides. Investig. Ophthalmol. Vis. Sci. 1997, 38, 842–847. [Google Scholar]
- Gilgun-Sherki, Y.; Rosenbaum, Z.; Melamed, E.; Offen, D. Antioxidant therapy in acute central nervous system injury: Current state. Pharmacol. Rev. 2002, 54, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Csukas, S.C.; Green, K. Effects of intracameral hydrogen peroxide in the rabbit anterior chamber. Investig. Ophthalmol. Vis. Sci. 1988, 29, 335–339. [Google Scholar]
- Artola, A.; Alio, J.L.; Bellot, J.L.; Ruiz, J.M. Lipid peroxidation in the iris and its protection by means of viscoelastic substances (sodium hyaluronate and hydroxypropylmethylcellulose). Ophthalmic. Res. 1993, 25, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Van der Vliet, A.; Bast, A. Effect of oxidative stress on receptors and signal transmission. Chem. Biol. Interact. 1992, 85, 95–116. [Google Scholar] [CrossRef] [PubMed]
- Flammer, J.; Haefliger, I.O.; Orgül, S.; Resink, T. Vascular dysregulation: A principal risk factor for glaucomatous damage? J. Glaucoma 1999, 8, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhu, X.; Lu, Y. The Proteome of Cataract Markers: Focus on Crystallins. Adv Clin. Chem. 2018, 86, 179–210. [Google Scholar] [PubMed]
- Delbarre, M.; Froussart-Maille, F. Signs, symptoms, and clinical forms of cataract in adults. J. Fr. Ophtalmol. 2020, 43, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, C.; Sardinha, L.R.; Rizzo, L.V. Cytokine and Chemokine Concentration in the Tear of Patients with Age-Related Cataract. Curr Eye Res. 2020, 45, 1101–1106. [Google Scholar] [CrossRef]
- Waris, G.; Ahsan, H. Reactive oxygen species: Role in the development of cancer and various chronic conditions. J. Carcinog. 2006, 5, 14. [Google Scholar] [CrossRef]
- Lü, J.M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell. Mol. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef]
- Ung, L.; Pattamatta, U.; Carnt, N.; Wilkinson-Berka, J.L.; Liew, G.; White, A.J. Oxidative stress and reactive oxygen species: A review of their role in ocular disease. Clin. Sci. 2017, 131, 2865–2883. [Google Scholar] [CrossRef]
- Kukreja, R.C.; Hess, M.L. The oxygen free radical system: From equations through membrane-protein interactions to cardiovascular injury and protection. Cardiovasc. Res. 1992, 26, 641–655. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell. Biol 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Sawada, H.; Fukuchi, T.; Abe, H. Oxidative stress markers in aqueous humor of patients with senile cataracts. Curr. Eye Res. 2009, 34, 36–41. [Google Scholar] [CrossRef]
- Kannabiran, C.; Balasubramanian, D. Molecular genetics of cataract. Indian J. Ophthalmol. 2000, 48, 5–13. [Google Scholar] [PubMed]
- Oh, M.S.; Carroll, H.J. The anion gap. Engl. J. Med. 1977, 297, 814–817. [Google Scholar] [CrossRef] [PubMed]
- Delaye, M.; Tardieu, A. Short-range order of crystallin proteins accounts for eye lens transparency. Nature 1983, 302, 415–417. [Google Scholar] [CrossRef]
- Makley, L.N.; McMenimen, K.A.; DeVree, B.T.; Goldman, J.W.; McGlasson, B.N.; Rajagopal, P.; Dunyak, B.M.; McQuade, T.J.; Thompson, A.D.; Sunahara, R.; et al. Pharmacological chaperone for α-crystallin partially restores transparency in cataract models. Science 2015, 350, 674–677. [Google Scholar] [CrossRef] [Green Version]
- Lampi, K.J.; Lampi, K.J.; Ma, Z.; Shih, M.; Shearer, T.R.; Smith, J.B.; Smith, D.L.; David, L.L. Sequence analysis of betaA3, betaB3, and betaA4 crystallins completes the identification of the major proteins in young human lens. J. Biol. Chem. 1997, 272, 2268–2275. [Google Scholar] [CrossRef] [Green Version]
- Robinson, N.E.; Lampi, K.J.; Speir, J.P.; Kruppa, G.; Easterling, M.; Robinson, A.B. Quantitative measurement of young human eye lens crystallins by direct injection Fourier transform ion cyclotron resonance mass spectrometry. Mol. Vis. 2006, 12, 704–711. [Google Scholar]
- Lampi, K.J.; Ma, Z.; Hanson, S.R.; Azuma, M.; Shih, M.; Shearer, T.R.; Smith, D.L.; Smith, J.B.; David, L.L. Age-related changes in human lens crystallins identified by two-dimensional electrophoresis and mass spectrometry. Exp. Eye Res. 1998, 67, 31–43. [Google Scholar] [CrossRef]
- Zhu, X.J.; Zhu, X.J.; Zhang, K.K.; He, W.W.; Du, Y.; Hooi, M.; Lu, Y. Racemization at the Asp 58 residue in αA-crystallin from the lens of high myopic cataract patients. J. Cell. Mol. Med. 2018, 22, 1118–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Sakai, M.; Sadakane, Y.; Haga, T.; Goto, Y.; Kinouchi, T.; Saito, T.; Fujii, N. Differential rate constants of racemization of aspartyl and asparaginyl residues in human alpha A-crystallin mutants. Biochim. Biophys. Acta 2008, 1784, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Simeonova, G.P.; Krastev, S.Z.; Simeonov, R.S. Immunological and pathological investigations in equine experimental uveitis. Vet. Res. Commun. 2016, 40, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Braakhuis, A.J.; Donaldson, C.I.; Lim, J.C.; Donaldson, P.J. Nutritional Strategies to Prevent Lens Cataract: Current Status and Future Strategies. Nutrients 2019, 11, 1186. [Google Scholar] [CrossRef] [Green Version]
- Tarwadi, K.V.; Chiplonkar, S.A.; Agte, V. Dietary and nutritional biomarkers of lens degeneration, oxidative stress and micronutrient inadequacies in Indian cataract patients. Clin. Nutr. 2008, 27, 464–472. [Google Scholar] [CrossRef]
- Balne, P.K.; Au, V.B.; Tong, L.; Ghosh, A.; Agrawal, M.; Connolly, J.; Agrawal, R. Bead Based Multiplex Assay for Analysis of Tear Cytokine Profiles. J. Vis. Exp. 2017, 128, e55993. [Google Scholar] [CrossRef]
- Willcox, M.D.; Argüeso, P.; Georgiev, G.A.; Holopainen, J.M.; Laurie, G.W.; Millar, T.J.; Papas, E.B.; Rolland, J.P.; Schmidt, T.A.; Stahl, U.; et al. TFOS DEWS II Tear Film Report. Ocul. Surf. 2017, 15, 366–403. [Google Scholar] [CrossRef]
- Chiva, A. Electrophoresis of tear proteins as a new diagnostic tool for two high risk groups for dry eye: Computer users and contact lens wearers. J. Med. Life 2011, 4, 228–233. [Google Scholar]
- Hagan, S. Biomarkers of ocular surface disease using impression cytology. Biomark. Med. 2017, 11, 1135–1147. [Google Scholar] [CrossRef] [Green Version]
- Villani, E.; Bonsignore, F.; Cantalamessa, E.; Serafino, M.; Nucci, P. Imaging Biomarkers for Dry Eye Disease. Eye Contact Lens 2020, 46 (Suppl. S2), S141–S145. [Google Scholar] [CrossRef]
- Baudouin, C.; Irkeç, M.; Messmer, E.M.; Benítez-Del-Castillo, J.M.; Bonini, S.; Figueiredo, F.C.; Geerling, G.; Labetoulle, M.; Lemp, M.; Rolando, M.; et al. Clinical impact of inflammation in dry eye disease: Proceedings of the ODISSEY group meeting. Acta Ophthalmol. 2018, 96, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, N.S.; Wei, Y.; Kuklinski, E.; Asbell, P.A. The Growing Need for Validated Biomarkers and Endpoints for Dry Eye Clinical Research. Investig. Ophthalmol. Vis. Sci. 2017, 58, BIO1–BIO19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, B.D.; Crews, L.; Messmer, E.M.; Foulks, G.N.; Nichols, K.K.; Baenninger, P.; Geerling, G.; Figueiredo, F.; Lemp, M.A. Correlations between commonly used objective signs and symptoms for the diagnosis of dry eye disease: Clinical implications. Acta Ophthalmol. 2014, 92, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Careba, I.; Chiva, A.; Totir, M.; Ungureanu, E.; Gradinaru, S. Tear lipocalin, lysozyme and lactoferrin concentrations in postmenopausal women. J. Med. Life 2015, 8, 94–98. [Google Scholar] [PubMed]
- Versura, P.; Nanni, P.; Bavelloni, A.; Blalock, W.L.; Piazzi, M.; Roda, A.; Campos, E.C. Tear proteomics in evaporative dry eye disease. Eye 2010, 24, 1396–1402. [Google Scholar] [CrossRef] [Green Version]
- Tamhane, M.; Cabrera-Ghayouri, S.; Abelian, G.; Viswanath, V. Review of Biomarkers in Ocular Matrices: Challenges and Opportunities. Pharm. Res. 2019, 36, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Hagan, S.; Martin, E.; Enríquez-de-Salamanca, A. Tear fluid biomarkers in ocular and systemic disease: Potential use for predictive, preventive and personalised medicine. EPMA J. 2016, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Beuerman, R.W.; Chan, C.M.; Zhao, S.Z.; Li, X.R.; Yang, H.; Tong, L.; Liu, S.; Stern, M.E.; Tan, D. Identification of tear fluid biomarkers in dry eye syndrome using iTRAQ quantitative proteomics. J. Proteome. Res. 2009, 8, 4889–4905. [Google Scholar] [CrossRef]
- Choi, W.; Lian, C.; Ying, L.; Kim, G.E.; You, I.C.; Park, S.H.; Yoon, K.C. Expression of Lipid Peroxidation Markers in the Tear Film and Ocular Surface of Patients with Non-Sjogren Syndrome: Potential Biomarkers for Dry Eye Disease. Curr. Eye Res. 2016, 41, 1143–1149. [Google Scholar] [CrossRef]
- Labbé, A.; Brignole-Baudouin, F.; Baudouin, C. Ocular surface investigations in dry eye. J. Fr. Ophtalmol. 2007, 30, 76–97. [Google Scholar] [CrossRef]
- Nichols, J.J.; Willcox, M.D.P.; Bron, A.J.; Belmonte, C.; Ciolino, J.B.; Craig, J.P.; Dogru, M.; Foulks, G.N.; Jones, L.; Nelson, J.D.; et al. The TFOS International Workshop on Contact Lens Discomfort: Executive summary. Investig. Ophthalmol Vis. Sci. 2013, 54, TFOS7–TFOS13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, S.; Tong, L. Practical issues concerning tear protein assays in dry eye. Eye Vis. 2014, 1, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramann, C.; Boehm, N.; Lorenz, K.; Wehrwein, N.; Stoffelns, B.M.; Pfeiffer, N.; Grus, F.H. Effect of contact lenses on the protein composition in tear film: A ProteinChip study. Graefes Arch. Clin. Exp. Ophthalmol. 2011, 249, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Belin, M.W.; Jang, H.S.; Borgstrom, M. Keratoconus: Diagnosis and Staging. Cornea 2022, 41, 1–11. [Google Scholar] [CrossRef]
- Katipoğlu, Z.; Mirza, E.; Oltulu, R.; Katipoglu, B. May Monocyte/HDL Cholesterol Ratio (MHR) and Neutrophil/Lymphocyte Ratio (NLR) Be an Indicator of Inflammation and Oxidative Stress in Patients with Keratoconus? Ocul. Immunol. Inflamm. 2020, 28, 632–636. [Google Scholar] [CrossRef]
- McKay, T.B.; Hjortdal, J.; Priyadarsini, S.; Karamichos, D. Acute hypoxia influences collagen and matrix metalloproteinase expression by human keratoconus cells in vitro. PLoS ONE 2017, 12, e0176017. [Google Scholar] [CrossRef] [Green Version]
- Lackner, E.M.; Matthaei, M.; Meng, H.; Ardjomand, N.; Eberhart, C.G.; Jun, A.S. Design and analysis of keratoconus tissue microarrays. Cornea 2014, 33, 49–55. [Google Scholar] [CrossRef]
- Balmus, I.M.; Alexa, A.I.; Ciuntu, R.-E.; Danielescu, C.; Stoica, B.; Cojocaru, S.I.; Ciobica, A.; Cantemir, A. Oxidative stress markers dynamics in keratoconus patients’ tears before and after corneal collagen crosslinking procedure. Exp. Eye Res. 2020, 190, 107897. [Google Scholar] [CrossRef]
- Kılıç, R.; Ayraktar, A.C.; Bayraktar, S.; Kurt, A.; Kavutçu, M. Evaluation of Serum Superoxide Dismutase Activity, Malondialdehyde, and Zinc and Copper Levels in Patients With Keratoconus. Cornea 2016, 35, 1512–1515. [Google Scholar] [CrossRef]
- Abdul-Maksoud, R.S.; Fouad, R.A.; Elsayed, T.G.; Ibrahem, R.A.; Badawi, A.E. The impact of catalase and glutathione peroxidase-1 genetic polymorphisms on their enzyme activities among Egyptian patients with keratoconus. J. Gene Med. 2020, 22, e3192. [Google Scholar] [CrossRef]
- Saijyothi, A.V.; Fowjana, J.; Madhumathi, S.; Rajeshwari, M.; Thennarasu, M.; Prema, P.; Angayarkanni, N. Tear fluid small molecular antioxidants profiling shows lowered glutathione in keratoconus. Exp. Eye Res. 2012, 103, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Arnal, E.; Peris-Martínez, C.; Menezo, J.L.; Johnsen-Soriano, S.; Romero, F.J. Oxidative stress in keratoconus? Investig. Ophthalmol. Vis. Sci. 2011, 52, 8592–8597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekin, S.; Seven, E. Assessment of serum catalase, reduced glutathione, and superoxide dismutase activities and malondialdehyde levels in keratoconus patients. Eye 2022, 36, 2062–2066. [Google Scholar] [CrossRef] [PubMed]
- Horwath-Winter, J.; Kirchengast, S.; Meinitzer, A.; Wachswender, C.; Faschinger, C.; Schmut, O. Determination of uric acid concentrations in human tear fluid, aqueous humour and serum. Acta Ophthalmol. 2009, 87, 188–192. [Google Scholar] [CrossRef]
- Bamdad, S.; Owji, N.; Bolkheir, A. Association Between Advanced Keratoconus and Serum Levels of Zinc, Calcium, Magnesium, Iron, Copper, and Selenium. Cornea 2018, 37, 1306–1310. [Google Scholar] [CrossRef]
- Zarei-Ghanavati, S.; Ahaghi, B.; Hassanzadeh, S.; Ghayour-Mobarhan, M.; Hakimi, H.R.; Eghbali, P. Serum 25-Hydroxyvitamin D, Selenium, Zinc and Copper in Patients with Keratoconus. J. Curr. Ophthalmol. 2020, 32, 26–31. [Google Scholar] [CrossRef]
- Balasubramanian, S.A.; Pye, D.C.; Willcox, M.D. Levels of lactoferrin, secretory IgA and serum albumin in the tear film of people with keratoconus. Exp. Eye Res. 2012, 96, 132–137. [Google Scholar] [CrossRef]
- Gupta, P.K.; Berdahl, J.P.; Chan, C.C.M.F.; Rocha, K.M.; Yeu, E.; Ayres, B.; Farid, M.; Lee, W.B.; Beckman, K.A.; Kim, T.; et al. The corneal endothelium: Clinical review of endothelial cell health and function. J. Cataract. Refract. Surg. 2021, 47, 1218–1226. [Google Scholar] [CrossRef]
- Epstein, S.P.; Gadaria-Rathod, N.; Wei, Y.; Maguire, M.G.; Asbell, P.A. HLA-DR expression as a biomarker of inflammation for multicenter clinical trials of ocular surface disease. Exp. Eye Res. 2013, 111, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Roebuck, K.A.; Finnegan, A. Regulation of intercellular adhesion molecule-1 (CD54) gene expression. J. Leukoc. Biol. 1999, 66, 876–888. [Google Scholar] [CrossRef]
- Jones, D.T.; Jones, D.T.; Monroy, D.; Ji, Z.; Atherton, S.S.; Pflugfelder, S.C. Sjögren’s syndrome: Cytokine and Epstein-Barr viral gene expression within the conjunctival epithelium. Investig. Ophthalmol. Vis. Sci. 1994, 35, 3493–3504. [Google Scholar]
- Tsubota, K.; Fujihara, T.; Saito, K.; Takeuchi, T. Conjunctival epithelium expression of HLA-DR in dry eye patients. Ophthalmologica 1999, 213, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Schindowski, K.; Schindowski, K.; Leutner, S.; Müller, W.E.; Eckert, A. Age-related changes of apoptotic cell death in human lymphocytes. Neurobiol. Aging 2000, 21, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Lee, T.L.; Chen, Y.C.; Liang, C.J.; Wang, S.H.; Lue, J.H.; Tsai, J.S.; Lee, S.W.; Chen, S.H.; Yang, Y.F.; et al. PM(2.5)-induced oxidative stress increases intercellular adhesion molecule-1 expression in lung epithelial cells through the IL-6/AKT/STAT3/NF-κB-dependent pathway. Part. Fibre Toxicol. 2018, 15, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, F.L.; Xiao, Y.; Bian, F.; Coursey, T.G.; Ko, B.Y.; Clevers, H.; De Paiva, C.S.; Pflugfelder, S.C. Goblet Cells Contribute to Ocular Surface Immune Tolerance-Implications for Dry Eye Disease. Int. J. Mol. Sci. 2017, 18, 978. [Google Scholar] [CrossRef] [Green Version]
- McCauley, H.A.; Liu, C.-Y.; Attia, A.C.; Wikenheiser-Brokamp, K.A.; Zhang, Y.; Whitsett, J.A.; Guasch, G. TGFβ signaling inhibits goblet cell differentiation via SPDEF in conjunctival epithelium. Development 2014, 141, 4628–4639. [Google Scholar] [CrossRef] [Green Version]
- Marko, C.K.; Menon, B.B.; Chen, G.; Whitsett, J.A.; Clevers, H.; Gipson, I.K. Spdef null mice lack conjunctival goblet cells and provide a model of dry eye. Am. J. Pathol. 2013, 183, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Nelson, J.D.; Wright, J.C. Conjunctival goblet cell densities in ocular surface disease. Arch. Ophthalmol. 1984, 102, 1049–1051. [Google Scholar] [CrossRef]
- Sommer, A. Effects of vitamin A deficiency on the ocular surface. Ophthalmology 1983, 90, 592–600. [Google Scholar] [CrossRef]
- Palace, V.P.; Khaper, N.; Qin, Q.; Singal, P.K. Antioxidant potentials of vitamin A and carotenoids and their relevance to heart disease. Free Radic. Biol. Med. 1999, 26, 746–761. [Google Scholar] [CrossRef]
- Dogru, M.; Matsumoto, Y.; Okada, N.; Igarashi, A.; Fukagawa, K.; Shimazaki, J.; Tsubota, K.; Fujishima, H. Alterations of the ocular surface epithelial MUC16 and goblet cell MUC5AC in patients with atopic keratoconjunctivitis. Allergy 2008, 63, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Takeyama, K.; Dabbagh, K.; Shim, J.J.; Dao-Pick, T.; Ueki, I.F.; Nadel, J.A. Oxidative stress causes mucin synthesis via transactivation of epidermal growth factor receptor: Role of neutrophils. J. Immunol. 2000, 164, 1546–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyfuss, J.L.; Regatieri, C.V.; Coelho, B.; Barbosa, J.B.; De Freitas, D.; Nader, H.; Martins, J.R. Altered hyaluronic acid content in tear fluid of patients with adenoviral conjunctivitis. An. Acad. Bras. Cienc. 2015, 87, 455–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grishko, V.; Xu, M.; Ho, R.; Mates, A.; Watson, S.; Kim, J.T.; Wilson, G.L.; Pearsall, A.W. Effects of hyaluronic acid on mitochondrial function and mitochondria-driven apoptosis following oxidative stress in human chondrocytes. J. Biol. Chem. 2009, 284, 9132–9139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, J. Ocular allergy test and biomarkers on the ocular surface: Clinical test for evaluating the ocular surface condition in allergic conjunctival diseases. Allergol. Int. 2020, 69, 496–504. [Google Scholar] [CrossRef]
- Kurtul, B.E.; Kabatas, E.U.; Boybeyi, S.D.; Caglar, A.A.; Ozer, P.A. Increased red cell distribution width levels in children with seasonal allergic conjunctivitis. Int. Ophthalmol. 2018, 38, 1079–1084. [Google Scholar] [CrossRef]
- Gueudry, J.; Muraine, M. Anterior uveitis. J. Fr. Ophtalmol. 2018, 41, e11–e21. [Google Scholar] [CrossRef]
- Bloch-Michel, E.; Nussenblatt, R.B. International Uveitis Study Group recommendations for the evaluation of intraocular inflammatory disease. Am. J. Ophthalmol. 1987, 103, 234–235. [Google Scholar] [CrossRef]
- Hunter, R.S.; Lobo, A.M. Current diagnostic approaches to infectious anterior uveitis. Int. Ophthalmol. Clin. 2011, 51, 145–156. [Google Scholar] [CrossRef]
- Nguyen, A.M.; Sève, P.; Le Scanff, J.; Gambrelle, J.; Fleury, J.; Broussolle, C.; Grange, J.D.; Kodjikian, L. Clinical and etiological aspects of uveitis: A retrospective study of 121 patients referred to a tertiary centre of ophthalmology. Rev. Med. Interne 2011, 32, 9–16. [Google Scholar] [CrossRef]
- McKay, K.M.; Jacobs, D.S. In Vivo Confocal Microscopy of Keratic Precipitates in Uveitis. Int. Ophthalmol. Clin. 2019, 59, 95–103. [Google Scholar] [CrossRef]
- Wertheim, M.S.; Mathers, W.D.; Planck, S.J.; Martin, T.M.; Suhler, E.B.; Smith, J.R.; Rosenbaum, J.T. In vivo confocal microscopy of keratic precipitates. Arch. Ophthalmol. 2004, 122, 1773–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesh, S.K.; Roopleen; Biswas, J.; Veena, N. Role of high-resolution computerized tomography (HRCT) of the chest in granulomatous uveitis: A tertiary uveitis clinic experience from India. Ocul. Immunol. Inflamm. 2011, 19, 51–57. [Google Scholar] [CrossRef]
- Doycheva, D.; Pfannenberg, C.; Hetzel, J.; Deuter, C.M.E.; Pavesio, C.; Kempf, V.A.J.; Schuelen, E.; Aschoff, P.; Rao, N.; Zierhut, M. Presumed tuberculosis-induced retinal vasculitis, diagnosed with positron emission tomography (18F-FDG-PET/CT), aspiration biopsy, and culture. Ocul. Immunol. Inflamm. 2010, 18, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Doycheva, D.; Deuter, C.; Hetzel, J.; Frick, J.-S.; Aschoff, P.; Schuelen, E.; Zierhut, M.; Pfannenberg, C. The use of positron emission tomography/CT in the diagnosis of tuberculosis-associated uveitis. Br. J. Ophthalmol. 2011, 95, 1290–1294. [Google Scholar] [CrossRef] [PubMed]
- Lalvani, A. Diagnosing tuberculosis infection in the 21st century: New tools to tackle an old enemy. Chest 2007, 131, 1898–1906. [Google Scholar] [CrossRef]
- Kato, A.; Ishihara, M.; Mizuki, N. Interferon-induced sarcoidosis with uveitis as the initial symptom: A case report and review of the literature. J. Med. Case. Rep. 2021, 15, 568. [Google Scholar] [CrossRef]
- Harper, T.W.; Miller, D.; Schiffman, J.C.; Davis, J.L. Polymerase chain reaction analysis of aqueous and vitreous specimens in the diagnosis of posterior segment infectious uveitis. Am J. Ophthalmol. 2009, 147, 140–147.e2. [Google Scholar] [CrossRef] [Green Version]
- Rothova, A.; de Boer, J.H.; Loon, N.H.T.D.-V.; Postma, G.; de Visser, L.; Zuurveen, S.J.; Schuller, M.; Weersink, A.J.; van Loon, A.M.; de Groot-Mijnes, J.D. Usefulness of aqueous humor analysis for the diagnosis of posterior uveitis. Ophthalmology 2008, 115, 306–311. [Google Scholar] [CrossRef]
- Choi, W.; Kang, H.G.; Choi, E.Y.; Kim, S.S.; Kim, C.Y.; Koh, H.J.; Lee, S.C.; Kim, M. Clinical utility of aqueous humor polymerase chain reaction and serologic testing for suspected infectious uveitis: A single-center retrospective study in South Korea. BMC Ophthalmol. 2020, 20, 242. [Google Scholar] [CrossRef]
- Sugita, S.; Shimizu, N.; Watanabe, K.; Mizukami, M.; Morio, T.; Sugamoto, Y.; Mochizuki, M. Use of multiplex PCR and real-time PCR to detect human herpes virus genome in ocular fluids of patients with uveitis. Br. J. Ophthalmol. 2008, 92, 928–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamid, S.; Gul, A.; Hamid, Q. Relationship of cytokines and AGE products in diabetic and non-diabetic patients with cataract. Int. J. Health. Sci. 2016, 10, 507–515. [Google Scholar] [CrossRef]
- Dong, Y.; Mu, G.-Y.; Chen, F.; Zhao, R.-L.; Wang, M.; Wang, B. Correlation between MMP-2 gene polymorphism and cataract susceptibility. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3167–3172. [Google Scholar] [PubMed]
- Klein, B.E.; Klein, B.E.; Klein, R.; Lee, K.E.; Knudtson, M.D.; Tsai, M.Y. Markers of inflammation, vascular endothelial dysfunction, and age-related cataract. Am. J. Ophthalmol. 2006, 141, 116–122. [Google Scholar] [CrossRef]
- Zheng, Y.; Rao, Y.-Q.; Li, J.-K.; Huang, Y.; Zhao, P.; Li, J. Age-related pro-inflammatory and pro-angiogenic changes in human aqueous humor. Int. J. Ophthalmol. 2018, 11, 196–200. [Google Scholar]
- Mitrović, S.; Kelava, T.; Šućur, A.; Grčević, D. Levels of Selected Aqueous Humor Mediators (IL-10, IL-17, CCL2, VEGF, FasL) in Diabetic Cataract. Ocul. Immunol. Inflamm. 2016, 24, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Wishart, T.F.L.; Flokis, M.; Shu, D.Y.; Das, S.J.; Lovicu, F.J. Hallmarks of lens aging and cataractogenesis. Exp. Eye. Res. 2021, 210, 108709. [Google Scholar] [CrossRef] [PubMed]
- Singh, S. Role of Malondialdehyde (MDA) in senile cataract. J. Med. Res. 2016, 2, 44–46. [Google Scholar]
- Bhatia, R.P.; Rai, R.; Rao, G.R. Role of malondialdehyde and superoxide dismutase in cataractogenesis. Ann. Ophthalmol. 2006, 38, 103–106. [Google Scholar] [CrossRef]
- Kaur, J.; Kukreja, S.; Kaur, A.; Malhotra, N.; Kaur, R. The oxidative stress in cataract patients. J. Clin. Diagn. Res. 2012, 6, 1629–1632. [Google Scholar] [CrossRef]
- Kisić, B.; Kisić, B.; Žorić, L.; Dolićanin, Z.; Mitić, R.; Mirić, M. The Impact of Senile Cataract Maturity on Blood Oxidative Stress Markers and Glutathione-Dependent Antioxidants: Relations with Lens Variables. J. Med. Biochem. 2012, 31, 184–192. [Google Scholar]
- Yildirim, Z.; Yildirim, F.; Uçgun, N.I.; Kiliç, N.; Yıldırım, F. The evaluation of the oxidative stress parameters in nondiabetic and diabetic senile cataract patients. Biol. Trace. Elem. Res. 2009, 128, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarczyk-Sedlak, I.; Folwarczna, J.; Sedlak, L.; Zych, M.; Wojnar, W.; Szumińska, I.; Wyględowska-Promieńska, D.; Mrukwa-Kominek, E.; Szumińska, I. Effect of caffeine on biomarkers of oxidative stress in lenses of rats with streptozotocin-induced diabetes. Arch. Med. Sci. 2019, 15, 1073–1080. [Google Scholar] [CrossRef]
- Alapure, B.; Praveen, M.R.; Gajjar, D.; Vasavada, A.R.; Rajkumar, S.; Johar, K. Matrix metalloproteinase-9 activity in human lens epithelial cells of cortical, posterior subcapsular, and nuclear cataracts. J. Cataract. Refract. Surg. 2009, 34, 2063–2067. [Google Scholar] [CrossRef] [PubMed]
- Augustin, A.J.; Spitznas, M.; Kaviani, N.; Meller, D.; Koch, F.H.; Grus, F.; Göbbels, M.J. Oxidative reactions in the tear fluid of patients suffering from dry eyes. Graefes Arch. Clin. Exp. Ophthalmol. 1995, 233, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, T.H.; Dogru, M.; Tsubota, K. Tearful relations: Oxidative stress, inflammation and eye diseases. Arq. Bras. Oftalmol. 2008, 71 (Suppl. S6), 72–79. [Google Scholar]
- Dogru, M.; Matsumoto, Y.; Yamamoto, Y.; Goto, E.; Saiki, M.; Shimazaki, J.; Takebayashi, T.; Tsubota, K. Lactoferrin in Sjögren’s syndrome. Ophthalmology 2007, 114, 2366–2367. [Google Scholar] [CrossRef] [PubMed]
- Sethu, S.; Shetty, R.; Deshpande, K.; Pahuja, N.; Chinnappaiah, N.; Agarwal, A.; Sharma, A.; Ghosh, A. Correlation between tear fluid and serum vitamin D levels. Eye Vis. 2016, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Peponis, V.; Papathanasiou, M.; Kapranou, A.; Magkou, C.; Tyligada, A.; Melidonis, A.; Drosos, T.; Sitaras, N.M. Protective role of oral antioxidant supplementation in ocular surface of diabetic patients. Br. J. Ophthalmol. 2002, 86, 1369–1373. [Google Scholar] [CrossRef] [Green Version]
- Jin, K.W.; Ro, J.W.; Shin, Y.J.; Hyon, J.Y.; Wee, W.R.; Park, S.G. Correlation of vitamin D levels with tear film stability and secretion in patients with dry eye syndrome. Acta Ophthalmol. 2017, 95, e230–e235. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Xiang, J.; Zhu, W.; Wei, A.; Le, Q.; Xu, J.; Zhou, X. Vitamin A Palmitate and Carbomer Gel Protects the Conjunctiva of Patients With Long-term Prostaglandin Analogs Application. J. Glaucoma 2016, 25, 487–492. [Google Scholar] [CrossRef]
- Huang, J.Y.; Yeh, P.T.; Hou, Y.C. A randomized, double-blind, placebo-controlled study of oral antioxidant supplement therapy in patients with dry eye syndrome. Clin. Ophthalmol. 2016, 10, 813–820. [Google Scholar] [PubMed] [Green Version]
- Dobrzynska, M.; Napierala, M.; Florek, E. Flavonoid Nanoparticles: A Promising Approach for Cancer Therapy. Biomolecules 2020, 10, 1268. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Huang, J.; Zhang, H.; Li, B.; Xu, M.; Zhang, Y.; Xie, M.; Li, X. Micronized curcumin fabricated by supercritical CO(2) to improve antibacterial activity against Pseudomonas aeruginosa. Artif. Cells Nanomed. Biotechnol. 2020, 48, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Favero, G.; Moretti, E.; Krajčíková, K.; Tomečková, V.; Rezzani, R. Evidence of Polyphenols Efficacy against Dry Eye Disease. Antioxidants 2021, 10, 190. [Google Scholar] [CrossRef]
- Lin, M.; Sun, X.; Ye, S.; Chen, Y.; Gao, J.; Yuan, F.; Lin, N.; Lawson, T.; Liu, Y.; Deng, R. A new antioxidant made from a pterostilbene functionalized graphene nanocomposite as an efficient treatment for dry eye disease. Front. Chem. 2022, 10, 942578. [Google Scholar] [CrossRef]
- van ‘t Erve, T.J. Strategies to decrease oxidative stress biomarker levels in human medical conditions: A meta-analysis on 8-iso-prostaglandin F(2α). Redox. Biol. 2018, 17, 284–296. [Google Scholar] [CrossRef]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Gomes, J.A.; Rapuano, C.J.; Belin, M.W.; Ambrósio, R. Global Consensus on Keratoconus Diagnosis. Cornea 2015, 34, e38–e39. [Google Scholar] [CrossRef]
- Cheung, I.M.; McGhee, C.N.; Sherwin, T. Beneficial effect of the antioxidant riboflavin on gene expression of extracellular matrix elements, antioxidants and oxidases in keratoconic stromal cells. Clin. Exp. Optom. 2014, 97, 349–355. [Google Scholar] [CrossRef]
- Pastori, V.; Tavazzi, S.; Lecchi, M. Lactoferrin-loaded contact lenses: Eye protection against oxidative stress. Cornea 2015, 34, 693–697. [Google Scholar] [CrossRef]
- Reiss, G.R.; Werness, P.G.; Zollman, P.E.; Brubaker, R.F. Ascorbic acid levels in the aqueous humor of nocturnal and diurnal mammals. Arch. Ophthalmol. 1986, 104, 753–755. [Google Scholar] [CrossRef] [PubMed]
- Gross, R.L. The effect of ascorbate on wound healing. Int. Ophthalmol. Clin. 2000, 40, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Gujral, G.S.; Gujral, G.S.; Askari, S.; Ahmad, S.; Zakir, S.M. Topical vitamin C, vitamin E, and acetylcysteine as corneal wound healing agents: A comparative study. Indian J. Ophthalmol. 2020, 68, 2935–2939. [Google Scholar]
- Azari, A.A.; Arabi, A. Conjunctivitis: A Systematic Review. J. Ophthalmic. Vis. Res. 2020, 15, 372–395. [Google Scholar] [CrossRef]
- Tredici, C.; Fasciani, R.; Villano, A.; Gambini, G.; Caporossi, A. Efficacy of eye drops containing crosslinked hyaluronic acid and CoQ10 in restoring ocular health exposed to chlorinated water. Eur. J. Ophthalmol. 2020, 30, 430–438. [Google Scholar] [CrossRef] [PubMed]
- López-de la Rosa, A.; Pinto-Fraga, J.; Arauzo, F.B.; Rodríguez, R.U.; González-García, M.J. Safety and Efficacy of an Artificial Tear Containing 0.3% Hyaluronic Acid in the Management of Moderate-to-Severe Dry Eye Disease. Eye Contact Lens 2017, 43, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Fraga, J.; de la Rosa, A.L.; Arauzo, F.B.; Rodríguez, R.U.; González-García, M.J. Efficacy and Safety of 0.2% Hyaluronic Acid in the Management of Dry Eye Disease. Eye Contact Lens 2017, 43, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Demir, U.; Demir, T.; Ilhan, N. The protective effect of alpha-lipoic acid against oxidative damage in rabbit conjunctiva and cornea exposed to ultraviolet radiation. Ophthalmologica 2005, 219, 49–53. [Google Scholar] [CrossRef]
- Chen, B.Y.; Lin, A.P.-C.; Chang, L.-S.; Huang, T.-P.; Liu, H.-J.; Luk, C.-P.; Lo, Y.-L.; Chang, H.-H. Dietary α-lipoic acid prevents UVB-induced corneal and conjunctival degeneration through multiple effects. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6757–6766. [Google Scholar] [CrossRef] [Green Version]
- Destefanis, S.; Giretto, D.; Muscolo, M.C.; Di Cerbo, A.; Guidetti, G.; Canello, S.; Giovazzino, A.; Centenaro, S.; Terrazzano, G. Clinical evaluation of a nutraceutical diet as an adjuvant to pharmacological treatment in dogs affected by Keratoconjunctivitis sicca. BMC Vet. Res. 2016, 12, 214. [Google Scholar]
- Unsal, A.I.A.; Kocaturk, T.; Gunel, C.; Meteoglu, I.; Omurlu, I.K.; Cakmak, H.; Demirci, B. Effect of Pycnogenol® on an experimental rat model of allergic conjunctivitis. Graefes Arch. Clin. Exp. Ophthalmol. 2018, 256, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Katsinas, N.; Rodríguez-Rojo, S.; Enríquez-de-Salamanca, A. Olive Pomace Phenolic Compounds and Extracts Can Inhibit Inflammatory- and Oxidative-Related Diseases of Human Ocular Surface Epithelium. Antioxidants 2021, 10, 1150. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kang, S.-S.; Kim, J.Y.; Tchah, H. The Antioxidant N-Acetylcysteine Inhibits Inflammatory and Apoptotic Processes in Human Conjunctival Epithelial Cells in a High-Glucose Environment. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5614–5621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balci, Y.I.; Acer, S.; Yagci, R.; Kucukatay, V.; Sarbay, H.; Bozkurt, K.; Polat, A. N-acetylcysteine supplementation reduces oxidative stress for cytosine arabinoside in rat model. Int. Ophthalmol. 2017, 37, 209–214. [Google Scholar] [CrossRef]
- Chen, Y.; Hong, X. Effects of carvedilol reduce conjunctivitis through changes in inflammation, NGF and VEGF levels in a rat model. Exp. Ther. Med. 2016, 11, 1987–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Kim, B.-M.; Shin, S.; Kim, T.-Y.; Chung, S.-H. Superoxide dismutase 3 attenuates experimental Th2-driven allergic conjunctivitis. Clin. Immunol. 2017, 176, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Troger, A.; Spahiu, V.; Perekhvatova, N.; Skulachev, M.; Petrov, A.; Chernyak, B.; Asbell, P. The Role of SKQ1 (Visomitin) in Inflammation and Wound Healing of the Ocular Surface. Ophthalmol. Ther. 2019, 8, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Elgouhary, S.M.; Elmazar, H.F.; Naguib, M.I.; Bayomy, N.R. Role of oxidative stress and vascular endothelial growth factor expression in pterygium pathogenesis and prevention of pterygium recurrence after surgical excision. Int. Ophthalmol. 2020, 40, 2593–2606. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Tsai, C.-F.; Tsai, M.-C.; Hsu, Y.-W.; Lu, F.-J. Inhibitory effects of rosmarinic acid on pterygium epithelial cells through redox imbalance and induction of extrinsic and intrinsic apoptosis. Exp. Eye Res. 2017, 160, 96–105. [Google Scholar] [CrossRef]
- López-Montemayor, P.; Zavala, J.; Montalvo-Parra, M.D.; Guerrero-Ramírez, G.I.; Mayolo-Deloisa, K.; Enriquez-Ochoa, D.; Martínez-García, B.; Loya-García, D.; Guerrero-Martínez, A.M.; Valdez-García, J.E. Phytochemical Profile and Antioxidant and Antiproliferative Activity of Sedum dendroideum on Pterygium Fibroblasts. Evid. Based. Complement. Altern. Med. 2021, 2021, 5814221. [Google Scholar] [CrossRef]
- Lingham, G.; Kugelman, J.; Charng, J.; Lee, S.S.; Yazar, S.; McKnight, C.M.; Coroneo, M.T.; Lucas, R.M.; Brown, H.; Stevenson, L.J.; et al. Conjunctival ultraviolet autofluorescence area decreases with age and sunglasses use. Br. J. Ophthalmol. 2021, 2021, 320284. [Google Scholar] [CrossRef] [PubMed]
- Shiratori, K.; Ohgami, K.; Ilieva, I.; Jin, X.-H.; Koyama, Y.; Miyashita, K.; Yoshida, K.; Kase, S.; Ohno, S. Effects of fucoxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Exp. Eye. Res. 2005, 81, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.H.; Ohgami, K.; Shiratori, K.; Suzuki, Y.; Hirano, T.; Koyama, Y.; Yoshida, K.; Ilieva, I.; Iseki, K.; Ohno, S. Inhibitory effects of lutein on endotoxin-induced uveitis in Lewis rats. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2562–2568. [Google Scholar] [CrossRef] [PubMed]
- Ohgami, K.; Ilieva, I.; Shiratori, K.; Koyama, Y.; Yoshida, K.; Kase, S.; Suzuki, Y.; Ohno, S.; Jin, X.-H.; Kitaichi, N.; et al. Anti-inflammatory effects of aronia extract on rat endotoxin-induced uveitis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Ohgami, K.; Shiratori, K.; Kotake, S.; Nishida, T.; Mizuki, N.; Yazawa, K.; Ohno, S. Effects of astaxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2694–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rooij, J.; thoe Schwartzenberg, S.G.; Mulder, P.G.; Baarsma, S.G. Oral vitamins C and E as additional treatment in patients with acute anterior uveitis: A randomised double masked study in 145 patients. Br. J. Ophthalmol. 1999, 83, 1277–1282. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.J.; Chu, K.O.; Yip, Y.W.Y.; Li, W.Y.; Yang, Y.P.; Chan, K.P.; Ren, J.L.; Chan, S.O.; Pang, C.P. Green tea extract treatment alleviates ocular inflammation in a rat model of endotoxin-induced uveitis. PLoS ONE 2014, 9, e103995. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Kanai, K.; Ozaki, M.; Kagawa, T.; Kita, M.; Yamashita, Y.; Nagai, N.; Tajima, K. Preventive effects of tyrosol, a natural phenolic compound, on anterior uveitis induced by anterior chamber paracentesis in healthy beagle dogs. J. Vet. Med. Sci. 2019, 81, 573–576. [Google Scholar] [CrossRef] [Green Version]
- Lal, B.; Kapoor, A.K.; Asthana, O.P.; Agrawal, P.K.; Prasad, R.; Kumar, P.; Srimal, R.C. Efficacy of curcumin in the management of chronic anterior uveitis. Phytother. Res. 1999, 13, 318–322. [Google Scholar] [CrossRef]
- Allegri, P.; Mastromarino, A.; Neri, P. Management of chronic anterior uveitis relapses: Efficacy of oral phospholipidic curcumin treatment. Long-term follow-up. Clin. Ophthalmol. 2010, 4, 1201–1206. [Google Scholar]
- Yang, C.H.; Yang, C.H.; Fang, I.M.; Lin, C.P.; Yang, C.M.; Chen, M.S. Effects of the NF-kappaB inhibitor pyrrolidine dithiocarbamate on experimentally induced autoimmune anterior uveitis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1339–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, I.M.; Yang, C.-H.; Lin, C.-P.; Yang, C.-M.; Chen, M.-S. Effects of pyrrolidine dithiocarbamate, an NF-kappaB inhibitor, on cytokine expression and ocular inflammation in experimental autoimmune anterior uveitis. J. Ocul. Pharmacol. Ther. 2005, 21, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Bora, N.S.; Sohn, J.-H.; Bora, P.S.; Kaplan, H.J.; Kulkarni, P. Anti-inflammatory effects of specific cyclooxygenase 2,5-lipoxygenase, and inducible nitric oxide synthase inhibitors on experimental autoimmune anterior uveitis (EAAU). Ocul. Immunol. Inflamm. 2005, 13, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnokova, N.B.; Neroev, V.V.; Beznos, O.V.; Beyshenova, G.A.; Panova, I.G.; Tatikolov, A.S. Effects of dexamethasone and superoxide dismutase instillations on clinical course of uveitis and local biochemical processes (experimental study). Vestn. Oftalmol. 2015, 131, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Beyer-Mears, A.; Farnsworth, P.N. Diminished sugar cataractogenesis by quercetin. Exp. Eye Res. 1979, 28, 709–716. [Google Scholar] [CrossRef]
- Ramana, B.V.; Raju, T.N.; Kumar, V.V.; Reddy, P.U.M. Defensive role of quercetin against imbalances of calcium, sodium, and potassium in galactosemic cataract. Biol. Trace Elem. Res. 2007, 119, 35–41. [Google Scholar] [CrossRef]
- Alexiou, P.; Pegklidou, K.; Chatzopoulou, M.; Nicolaou, I.; Demopoulos, V.J. Aldose reductase enzyme and its implication to major health problems of the 21(st) century. Curr. Med. Chem. 2009, 16, 734–752. [Google Scholar] [CrossRef]
- Singh, A.; Bodakhe, S.H. Resveratrol delay the cataract formation against naphthalene-induced experimental cataract in the albino rats. J. Biochem. Mol. Toxicol. 2020, 34, e22420. [Google Scholar] [CrossRef]
- Doganay, S.; Borazan, M.; Iraz, M.; Çiğremiş, Y. The effect of resveratrol in experimental cataract model formed by sodium selenite. Curr. Eye Res. 2006, 31, 147–153. [Google Scholar] [CrossRef]
- Higashi, Y.; Higashi, K.; Mori, A.; Sakamoto, K.; Ishii, K.; Nakahara, T. Anti-cataract Effect of Resveratrol in High-Glucose-Treated Streptozotocin-Induced Diabetic Rats. Biol. Pharm. Bull. 2018, 41, 1586–1592. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Rivera, F.; Concheiro, A.; Alvarez-Lorenzo, C. Epalrestat-loaded silicone hydrogels as contact lenses to address diabetic-eye complications. Eur. J. Pharm. Biopharm. 2018, 122, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chai, F.-Y.; Yan, H.; Guo, Y.; Harding, J. Effects of N-acetylcysteine and glutathione ethyl ester drops on streptozotocin-induced diabetic cataract in rats. Mol. Vis. 2008, 14, 862–870. [Google Scholar] [PubMed]
- Carey, J.W.; Pinarci, E.Y.; Penugonda, S.; Karacal, H.; Ercal, N. In vivo inhibition of l-buthionine-(S,R)-sulfoximine-induced cataracts by a novel antioxidant, N-acetylcysteine amide. Free Radic. Biol. Med. 2011, 50, 722–729. [Google Scholar] [CrossRef]
- Yigit, E.A.; Ercal, N. Release of N-acetylcysteine and N-acetylcysteine amide from contact lenses. Eye Contact Lens 2013, 39, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Babizhayev, M.A.; Guiotto, A.; Kasus-Jacobi, A. N-Acetylcarnosine and histidyl-hydrazide are potent agents for multitargeted ophthalmic therapy of senile cataracts and diabetic ocular complications. J. Drug Target. 2009, 17, 36–63. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Deyev, A.I.; Yermakova, V.N.; Semiletov, Y.A.; Davydova, N.G.; Kurysheva, N.I.; Zhukotskii, A.V.; Goldman, I.M. N-Acetylcarnosine, a natural histidine-containing dipeptide, as a potent ophthalmic drug in treatment of human cataracts. Peptides 2001, 22, 979–994. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Deyev, A.I.; Yermakova, V.N.; Semiletov, Y.A.; Davydova, N.G.; Doroshenko, V.S.; Zhukotskii, A.V.; Goldman, I.M. Efficacy of N-acetylcarnosine in the treatment of cataracts. Drugs R & D 2002, 3, 87–103. [Google Scholar]
- Babizhayev, M.; Deyev, A.; Yermakova, V.; Remenshchikov, V.; Bours, J. Revival of the Lens Transparency with N-Acetylcarnosine. Curr. Drug Ther. 2006, 100, 91–116. [Google Scholar] [CrossRef]
- Babizhayev, M.A.; Burke, L.; Micans, P.; Richer, S.P. N-Acetylcarnosine sustained drug delivery eye drops to control the signs of ageless vision: Glare sensitivity, cataract amelioration and quality of vision currently available treatment for the challenging 50,000-patient population. Clin. Interv. Aging 2009, 4, 31–50. [Google Scholar] [CrossRef] [Green Version]
- Klepac, N.; Rudeš, Z.; Klepac, R. Effects of melatonin on plasma oxidative stress in rats with streptozotocin induced diabetes. Biomed. Pharmacother. 2006, 60, 32–35. [Google Scholar] [CrossRef]
- Aksoy, N.; Sabuncu, T.; Aksoy, S. Effects of melatonin on oxidative-antioxidative status of tissues in streptozotocin-induced diabetic rats. Cell Biochem. Funct. 2003, 21, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Khorsand, M.; Akmali, M.; Sharzad, S.; Beheshtitabar, M. Melatonin Reduces Cataract Formation and Aldose Reductase Activity in Lenses of Streptozotocin-induced Diabetic Rat. Iran. J. Med. Sci. 2016, 41, 305–313. [Google Scholar]
- Taysi, S.; Memisogullari, R.; Koc, M.; Yazici, A.T.; Aslankurt, M.; Gumustekin, K.; Al, B.; Ozabacigil, F.; Yilmaz, A.; Ozder, H.T. Melatonin reduces oxidative stress in the rat lens due to radiation-induced oxidative injury. Int. J. Radiat. Biol. 2008, 84, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Anwar, M.M.; Moustafa, M.A. The effect of melatonin on eye lens of rats exposed to ultraviolet radiation. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2001, 129, 57–63. [Google Scholar] [CrossRef]
- Karslioglu, I.; Ertekin, M.V.; Taysi, S.; Koçer, I.; Sezen, O.; Gepdiremen, A.; Koç, M.; Bakan, N. Radioprotective effects of melatonin on radiation-induced cataract. J. Radiat. Res. 2005, 46, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Hegde, K.; Varma, S. Protective effect of ascorbate against oxidative stress in the mouse lens. Biochim. Biophys. Acta 2004, 1670, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.; Lu, M.; Dudek, E.; Reddan, J.; Taylor, A. Vitamin C and vitamin E restore the resistance of GSH-depleted lens cells to H2O2. Free Radic. Biol. Med. 2003, 34, 521–530. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Hashizume, K.; Kishimoto, S.; Tezuka, Y.; Nishigori, H.; Yamamoto, N.; Kondo, Y.; Maruyama, N.; Ishigami, A.; Kurosaka, D. Effect of vitamin C depletion on UVR-B induced cataract in SMP30/GNL knockout mice. Exp. Eye Res. 2012, 94, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.N.; Giblin, F.J.; Lin, L.R.; Chakrapani, B. The effect of aqueous humor ascorbate on ultraviolet-B-induced DNA damage in lens epithelium. Investig. Ophthalmol. Vis. Sci. 1998, 39, 344–350. [Google Scholar]
- Özkaya, D.; Naziroğlu, M.; Armağan, A.; Demirel, A.; Köroglu, B.K.; Çolakoğlu, N.; Kükner, A.; Sönmez, T.T. Dietary vitamin C and E modulates oxidative stress induced-kidney and lens injury in diabetic aged male rats through modulating glucose homeostasis and antioxidant systems. Cell Biochem. Funct. 2011, 29, 287–293. [Google Scholar] [CrossRef]
- Ravindran, R.D.; Vashist, P.; Gupta, S.K.; Young, I.S.; Maraini, G.; Camparini, M.; Jayanthi, R.; John, N.; Fitzpatrick, K.E.; Chakravarthy, U.; et al. Inverse association of vitamin C with cataract in older people in India. Ophthalmology 2011, 118, 1958–1965.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mares, J.A.; Voland, R.; Adler, R.; Tinker, L.; Millen, A.E.; Moeller, S.M.; Chappell, R.J.; Neuhouser, M.L.; Sarto, G.E.; CAREDS Group. Healthy diets and the subsequent prevalence of nuclear cataract in women. Arch. Ophthalmol. 2010, 128, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor-Valero, M. Fruit and vegetable intake and vitamins C and E are associated with a reduced prevalence of cataract in a Spanish Mediterranean population. BMC Ophthalmol. 2013, 13, 52. [Google Scholar] [CrossRef] [Green Version]
- Theodoropoulou, S.; Samoli, E.; Theodossiadis, P.G.; Papathanassiou, M.; Lagiou, A.; Lagiou, P.; Tzonou, A. Diet and cataract: A case-control study. Int. Ophthalmol. 2014, 34, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Christen, W.G.; Glynn, R.J.; Sesso, H.D.; Kurth, T.; MacFadyen, J.; Bubes, V.; Buring, J.E.; Manson, J.E.; Gaziano, J.M. Age-related cataract in a randomized trial of vitamins E and C in men. Arch. Ophthalmol 2010, 128, 1397–1405. [Google Scholar] [CrossRef] [Green Version]
- Christen, W.G.; Glynn, R.J.; Chew, E.Y.; Buring, J.E. Vitamin E and age-related cataract in a randomized trial of women. Ophthalmology 2008, 115, 822–829.e1. [Google Scholar] [CrossRef]
- Gritz, D.C.; Srinivasan, M.; Smith, S.D.; Kim, U.; Lietman, T.M.; Wilkins, J.H.; Priyadharshini, B.; John, R.K.; Aravind, S.; Prajna, N.V.; et al. The Antioxidants in Prevention of Cataracts Study: Effects of antioxidant supplements on cataract progression in South India. Br. J. Ophthalmol. 2006, 90, 847–851. [Google Scholar] [CrossRef]
- Christen, W.G.; Glynn, R.J.; Gaziano, J.M.; Darke, A.K.; Crowley, J.J.; Goodman, P.J.; Lippman, S.M.; Lad, T.E.; Bearden, J.D.; Goodman, G.E.; et al. Age-related cataract in men in the selenium and vitamin e cancer prevention trial eye endpoints study: A randomized clinical trial. JAMA Ophthalmol. 2015, 133, 17–24. [Google Scholar] [CrossRef]
- McNeil, J.J.; Robman, L.; Tikellis, G.; Sinclair, M.I.; McCarty, C.A.; Taylor, H.R. Vitamin E supplementation and cataract: Randomized controlled trial. Ophthalmology 2004, 111, 75–84. [Google Scholar] [CrossRef]
- Rautiainen, S.; Rautiainen, S.; Lindblad, B.E.; Morgenstern, R.; Wolk, A. Vitamin C supplements and the risk of age-related cataract: A population-based prospective cohort study in women. Am. J. Clin. Nutr. 2010, 91, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Zheng Selin, J.; Selin, J.Z.; Rautiainen, S.; Lindblad, B.E.; Morgenstern, R.; Wolk, A. High-dose supplements of vitamins C and E, low-dose multivitamins, and the risk of age-related cataract: A population-based prospective cohort study of men. Am. J. Epidemiol. 2013, 177, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Age-Related Eye Disease Study Research Group. A randomized, placebo-controlled, clinical trial of high-dose supplementation with vitamins C and E, beta carotene, and zinc for age-related macular degeneration and vision loss: AREDS report no. 8. Arch. Ophthalmol. 2001, 119, 1417–1436. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Disease | Biomarker | References |
|---|---|---|
| Dry Eye | TGF-β2, matrix protease inhibitors | Baudouin, C., et al. [141] |
| TNF-α, IL-17A, IL-6, IL-8, MMP-9 | Roy, N.S., et al. [142,143] | |
| Secretory phospholipase A2, prostaglandin E2, arachidonic acid, docosahexaenoic acid, eicosapentaenoic acid, leukotriene B4 | Roy, N.S., et al. [142] | |
| Lysozyme-C, lipocalin 1, lactoferrin, lysozyme proline-rich protein 4, 3 | Tamhane, M., et al. [146] | |
| Annexin 5, alpha 2-glycoprotein 1, lacritin, caspase 14, proline-rich protein 3 and 4, cystatin S, cathepsin B, secretoglobin 1D1 Prolactin inducible protein Mucin 5AC | Careba, I., et al. [144,147,148] | |
| Annexin 2, Enolase 1α, Albumin, Nerve growth factor Clusterin, β2 microglobulin, Calgranulin A (S100 A8), B (S100 A9) Cystatin SN, Cathepsin S, Defensins α and β, Glycoprotein 340, Secretoglobin 2A | Careba, I., et al. [144,147,148] | |
| lactoferrin of <18%, lysozyme of <35% albumin of >15% | Careba, I., et al. [144] | |
| 4-hydroxy-2-nonenal and malondialdehyde | Choi, W., et al. [149] | |
| S100A Superoxide dismutase, Peroxidase, catalase, | Labbé, A., F [150] | |
| Secretoglobin 1D1, β2 microglobulin, proline rich protein 4, lacritin | D’Souza, S. and L. Tong, [152,153] | |
| Protein S100A8 secretoglobin 1 A2, albumin, nerve growth factor, prolactin inducible protein | Kramann, C., et al. [142,143,147,148,152,153] | |
| Keratoconus | Monocyte/High-density lipoprotein cholesterol ratio (MHR) Neutrophil/lymphocyte ratio (NLR) | Katipoğlu, Z., et al. [155] |
| 8-OHdG | McKay, T.B., et al. [156,157] | |
| GPx, MDA | Balmus, I.M., et al. [158] | |
| CAT | Abdul-Maksoud, R.S., et al. [160] | |
| Native and total thiol-disulfide | Gulpamuk, B., et al. [43] | |
| Glutathione | Saijyothi, A.V., et al. [43,161,162,163]. | |
| L-tyrosine | Saijyothi, A.V., et al. [161] | |
| Uric acid | Horwath-Winter, J., et al. [164] | |
| Decreased Cu, Zn and Se | Bamdad, S., N. Owji, and A. Bolkheir. [165,166] | |
| Lactoferrin | Balasubramanian, S.A., D.C. Pye, and M.D. Willcox [167] | |
| Keratoconjunctivitis | TGF beta | McCauley, H.A., et al. [176] |
| 5AC (MUC5AD) | Dogru, M., et al. [181] | |
| Hyaluronic acid (HA) | Dreyfuss, J.L., et al. [183] | |
| Eotaxin, Tumor necrosis factor-alpha soluble IL-6 receptor | Shoji, J., et al. [185] | |
| Osteopontin | Shoji, J., et al. [185] | |
| Red cell distribution width (RDW) | Kurtul, B.E., et al. [186] | |
| Uveitis | Interferon-Ƴ | Kato, A., M. Ishihara, and N. Mizuki [197] |
| Cataract | IL-6, IL-8 and TNF- α | Hamid et al. [202] |
| IL-6, IL-1β, CRP, and TNF-1α | [203] | |
| IL-6 and s-ICAM-1 | E K Klein et al. [204] | |
| IFN-γ, IL-6, IL-13, IL-12, IL-10, IFN-α2, CCL2, CCL3, CCL4, CXCL8, CXCL9, CXCL10 | Zheng, Y., et al. [205] | |
| VEGF IL-10 | Mitrovic et al. [206] | |
| MDA | Singh, S. [208,209,210,211] | |
| TGFβ and MMP-9 | Alapure et al. [214] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dammak, A.; Pastrana, C.; Martin-Gil, A.; Carpena-Torres, C.; Peral Cerda, A.; Simovart, M.; Alarma, P.; Huete-Toral, F.; Carracedo, G. Oxidative Stress in the Anterior Ocular Diseases: Diagnostic and Treatment. Biomedicines 2023, 11, 292. https://doi.org/10.3390/biomedicines11020292
Dammak A, Pastrana C, Martin-Gil A, Carpena-Torres C, Peral Cerda A, Simovart M, Alarma P, Huete-Toral F, Carracedo G. Oxidative Stress in the Anterior Ocular Diseases: Diagnostic and Treatment. Biomedicines. 2023; 11(2):292. https://doi.org/10.3390/biomedicines11020292
Chicago/Turabian StyleDammak, Azza, Cristina Pastrana, Alba Martin-Gil, Carlos Carpena-Torres, Assumpta Peral Cerda, Mirjam Simovart, Pilar Alarma, Fernando Huete-Toral, and Gonzalo Carracedo. 2023. "Oxidative Stress in the Anterior Ocular Diseases: Diagnostic and Treatment" Biomedicines 11, no. 2: 292. https://doi.org/10.3390/biomedicines11020292