


Thioacetamide-Induced Acute Liver Injury Increases Metformin Plasma Exposure by Downregulating Renal OCT2 and MATE1 Expression and Function


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Development of ALI Rats
2.4. Pharmacokinetics of Metformin after Oral or Intravenous Administration to Rats
2.5. Intestinal Absorption of Metformin in Rat Duodenum
2.6. Urinary Secretion of Metformin
2.7. Western Blot
2.8. Real-Time RT-PCR Analysis mRNA Levels of OCT2 and MATE1 in the Kidney of Rats
2.9. Cell Culture of HK-2 and Drug Treatment
2.10. Statistical Analysis
3. Results
3.1. Biochemical and Physiological Parameters of ALI Rats
3.2. Plasma Concentration of Metformin in Rats after Metformin Gavage
3.3. Alternations in Intestinal Absorption of Metformin in ALI Rats
3.4. Plasma Concentration of Metformin in Rats after Intravenous Injection
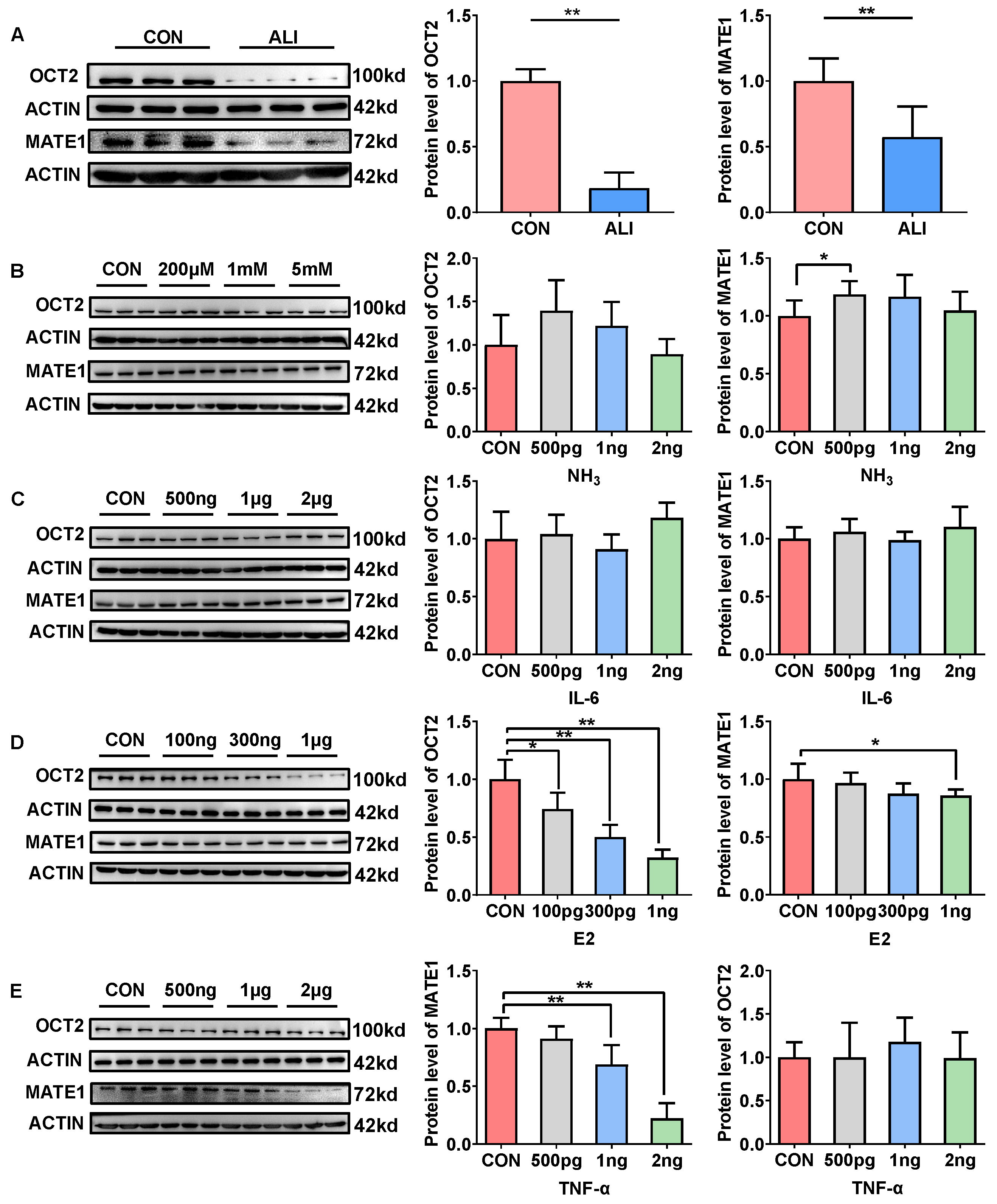
3.5. ALI Downregulates Renal OCT2 and MATE1 Expression to Decrease Metformin Excretion
3.6. Mechanisms of ALI Downregulation of Renal OCT2 and MATE1 Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stravitz, R.T.; Lee, W.M. Acute liver failure. Lancet 2019, 394, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Singanayagam, A.; Triantafyllou, E. Macrophages in Chronic Liver Failure: Diversity, Plasticity and Therapeutic Targeting. Front. Immunol. 2021, 12, 661182. [Google Scholar] [CrossRef]
- Arroyo, V.; Angeli, P.; Moreau, R.; Jalan, R.; Claria, J.; Trebicka, J.; Fernandez, J.; Gustot, T.; Caraceni, P.; Bernardi, M.; et al. The systemic inflammation hypothesis: Towards a new paradigm of acute decompensation and multiorgan failure in cirrhosis. J. Hepatol. 2021, 74, 670–685. [Google Scholar] [CrossRef] [PubMed]
- Simonetto, D.A.; Gines, P.; Kamath, P.S. Hepatorenal syndrome: Pathophysiology, diagnosis, and management. BMJ 2020, 370, m2687. [Google Scholar] [CrossRef] [PubMed]
- Gines, P.; Guevara, M.; Arroyo, V.; Rodes, J. Hepatorenal syndrome. Lancet 2003, 362, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Frassetto, L.; Benet, L.Z. Effects of renal failure on drug transport and metabolism. Pharmacol. Ther. 2006, 109, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Miao, M.X.; Sun, B.B.; Wang, Z.J.; Tang, X.G.; Chen, Y.; Zhao, K.J.; Liu, X.D.; Liu, L. Acute liver failure enhances oral plasma exposure of zidovudine in rats by downregulation of hepatic UGT2B7 and intestinal P-gp. Acta Pharmacol. Sin. 2017, 38, 1554–1565. [Google Scholar] [CrossRef] [PubMed]
- Crowley, M.J.; Diamantidis, C.J.; McDuffie, J.R.; Cameron, C.B.; Stanifer, J.W.; Mock, C.K.; Wang, X.; Tang, S.; Nagi, A.; Kosinski, A.S.; et al. Clinical Outcomes of Metformin Use in Populations With Chronic Kidney Disease, Congestive Heart Failure, or Chronic Liver Disease: A Systematic Review. Ann. Intern. Med. 2017, 166, 191–200. [Google Scholar] [CrossRef]
- Hong, S.; Li, S.; Meng, X.; Li, P.; Wang, X.; Su, M.; Liu, X.; Liu, L. Bile duct ligation differently regulates protein expressions of organic cation transporters in intestine, liver and kidney of rats through activation of farnesoid X receptor by cholate and bilirubin. Acta Pharm. Sin. B 2023, 13, 227–245. [Google Scholar] [CrossRef]
- Türk, D.; Scherer, N.; Selzer, D.; Dings, C.; Hanke, N.; Dallmann, R.; Schwab, M.; Timmins, P.; Nock, V.; Lehr, T. Significant impact of time-of-day variation on metformin pharmacokinetics. Diabetologia 2023, 66, 1024–1034. [Google Scholar] [CrossRef]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef] [PubMed]
- Ciarimboli, G. Regulation Mechanisms of Expression and Function of Organic Cation Transporter 1. Front. Pharmacol. 2020, 11, 607613. [Google Scholar] [CrossRef] [PubMed]
- Lozano, E.; Herraez, E.; Briz, O.; Robledo, V.S.; Hernandez-Iglesias, J.; Gonzalez-Hernandez, A.; Marin, J.J. Role of the plasma membrane transporter of organic cations OCT1 and its genetic variants in modern liver pharmacology. Biomed. Res. Int. 2013, 2013, 692071. [Google Scholar] [CrossRef] [PubMed]
- Ciarimboli, G.; Lancaster, C.S.; Schlatter, E.; Franke, R.M.; Sprowl, J.A.; Pavenstädt, H.; Massmann, V.; Guckel, D.; Mathijssen, R.H.; Yang, W.; et al. Proximal tubular secretion of creatinine by organic cation transporter OCT2 in cancer patients. Clin. Cancer Res. 2012, 18, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.; Lips, K.; Volk, C. Polyspecific organic cation transporters: Structure, function, physiological roles, and biopharmaceutical implications. Pharm. Res. 2007, 24, 1227–1251. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, H.; Inui, K. Organic cation transporter OCTs (SLC22) and MATEs (SLC47) in the human kidney. AAPS J. 2013, 15, 581–588. [Google Scholar] [CrossRef]
- Kimura, N.; Masuda, S.; Tanihara, Y.; Ueo, H.; Okuda, M.; Katsura, T.; Inui, K. Metformin is a superior substrate for renal organic cation transporter OCT2 rather than hepatic OCT1. Drug Metab. Pharmacokinet. 2005, 20, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Giacomini, K.M. Transporters Involved in Metformin Pharmacokinetics and Treatment Response. J. Pharm. Sci. 2017, 106, 2245–2250. [Google Scholar] [CrossRef]
- Toyama, K.; Yonezawa, A.; Masuda, S.; Osawa, R.; Hosokawa, M.; Fujimoto, S.; Inagaki, N.; Inui, K.; Katsura, T. Loss of multidrug and toxin extrusion 1 (MATE1) is associated with metformin-induced lactic acidosis. Br. J. Pharmacol. 2012, 166, 1183–1191. [Google Scholar] [CrossRef]
- Konig, J.; Zolk, O.; Singer, K.; Hoffmann, C.; Fromm, M.F. Double-transfected MDCK cells expressing human OCT1/MATE1 or OCT2/MATE1: Determinants of uptake and transcellular translocation of organic cations. Br. J. Pharmacol. 2011, 163, 546–555. [Google Scholar] [CrossRef]
- Muller, F.; Weitz, D.; Mertsch, K.; Konig, J.; Fromm, M.F. Importance of OCT2 and MATE1 for the Cimetidine-Metformin Interaction: Insights from Investigations of Polarized Transport in Single- And Double-Transfected MDCK Cells with a Focus on Perpetrator Disposition. Mol. Pharm. 2018, 15, 3425–3433. [Google Scholar] [CrossRef] [PubMed]
- Meyer zu Schwabedissen, H.E.; Verstuyft, C.; Kroemer, H.K.; Becquemont, L.; Kim, R.B. Human multidrug and toxin extrusion 1 (MATE1/SLC47A1) transporter: Functional characterization, interaction with OCT2 (SLC22A2), and single nucleotide polymorphisms. Am. J. Physiol. Ren. Physiol. 2010, 298, F997–F1005. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Terada, T.; Mizuno, T.; Katsura, T.; Shimakura, J.; Inui, K. Targeted disruption of the multidrug and toxin extrusion 1 (mate1) gene in mice reduces renal secretion of metformin. Mol. Pharmacol. 2009, 75, 1280–1286. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.W.; Bedwell, D.W.; Zamek-Gliszczynski, M.J. Ablation of both organic cation transporter (OCT)1 and OCT2 alters metformin pharmacokinetics but has no effect on tissue drug exposure and pharmacodynamics. Drug Metab. Dispos. 2012, 40, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Karimian Pour, N.; McColl, E.R.; Piquette-Miller, M. Impact of Viral Inflammation on the Expression of Renal Drug Transporters in Pregnant Rats. Pharmaceutics 2019, 11, 624. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Lauber, C.; Harrach, S.; Pap, T.; Fischer, M.; Victor, M.; Heitzmann, M.; Hansen, U.; Fobker, M.; Brand, S.M.; Sindic, A.; et al. Transport mechanisms and their pathology-induced regulation govern tyrosine kinase inhibitor delivery in rheumatoid arthritis. PLoS ONE 2012, 7, e52247. [Google Scholar] [CrossRef]
- Karimian Pour, N.; Piquette-Miller, M. Endotoxin Modulates the Expression of Renal Drug Transporters in HIV-1 Transgenic Rats. J. Pharm. Pharm. Sci. 2018, 21, 117s–129s. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, K.; Yoda, N.; Morokado, F.; Komori, H.; Nakanishi, T.; Tamai, I. Changes of drug pharmacokinetics mediated by downregulation of kidney organic cation transporters Mate1 and Oct2 in a rat model of hyperuricemia. PLoS ONE 2019, 14, e0214862. [Google Scholar] [CrossRef]
- He, R.; Ai, L.; Zhang, D.; Wan, L.; Zheng, T.; Yin, J.; Lu, H.; Lu, J.; Lu, F.; Liu, F.; et al. Different effect of testosterone and oestrogen on urinary excretion of metformin via regulating OCTs and MATEs expression in the kidney of mice. J. Cell. Mol. Med. 2016, 20, 2309–2317. [Google Scholar] [CrossRef]
- Jin, S.; Wang, X.T.; Liu, L.; Yao, D.; Liu, C.; Zhang, M.; Guo, H.F.; Liu, X.D. P-glycoprotein and multidrug resistance-associated protein 2 are oppositely altered in brain of rats with thioacetamide-induced acute liver failure. Liver Int. 2013, 33, 274–282. [Google Scholar] [CrossRef]
- Marciniak, S.; Wnorowski, A.; Smolińska, K.; Walczyna, B.; Turski, W.; Kocki, T.; Paluszkiewicz, P.; Parada-Turska, J. Kynurenic Acid Protects against Thioacetamide-Induced Liver Injury in Rats. Anal. Cell. Pathol. 2018, 2018, 1270483. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, R.E.; Salama, A.A.A.; Abdel-Rahman, R.F.; Ogaly, H.A. Hepato- and neuro-protective influences of biopropolis on thioacetamide-induced acute hepatic encephalopathy in rats. Can. J. Physiol. Pharmacol. 2017, 95, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Dai, Y.; Liu, Z.; Wang, C.; Meng, Q.; Huo, X.; Sun, H.; Ma, X.; Peng, J.; Liu, K. Involvement of organic cation transporter 2 in the metformin-associated increased lactate levels caused by contrast-induced nephropathy. Biomed. Pharmacother. 2018, 106, 1760–1766. [Google Scholar] [CrossRef]
- Jayakumar, A.R.; Valdes, V.; Norenberg, M.D. The Na-K-Cl cotransporter in the brain edema of acute liver failure. J. Hepatol. 2011, 54, 272–278. [Google Scholar] [CrossRef]
- Bruck, R.; Aeed, H.; Avni, Y.; Shirin, H.; Matas, Z.; Shahmurov, M.; Avinoach, I.; Zozulya, G.; Weizman, N.; Hochman, A. Melatonin inhibits nuclear factor kappa B activation and oxidative stress and protects against thioacetamide induced liver damage in rats. J. Hepatol. 2004, 40, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Xu, P.; Sun, B.; Zhong, Z.; Liu, C.; Ling, Z.; Chen, Y.; Shu, N.; Zhao, K.; et al. Acute liver failure impairs function and expression of breast cancer-resistant protein (BCRP) at rat blood-brain barrier partly via ammonia-ROS-ERK1/2 activation. J. Neurochem. 2016, 138, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Komazawa, H.; Yamaguchi, H.; Hidaka, K.; Ogura, J.; Kobayashi, M.; Iseki, K. Renal uptake of substrates for organic anion transporters Oat1 and Oat3 and organic cation transporters Oct1 and Oct2 is altered in rats with adenine-induced chronic renal failure. J. Pharm. Sci. 2013, 102, 1086–1094. [Google Scholar] [CrossRef]
- Porta, V.; Schramm, S.G.; Kano, E.K.; Koono, E.E.; Armando, Y.P.; Fukuda, K.; Serra, C.H. HPLC-UV determination of metformin in human plasma for application in pharmacokinetics and bioequivalence studies. J. Pharm. Biomed. Anal. 2008, 46, 143–147. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, H.; Xu, J.; Zhao, K.; Chen, Y.; Liang, L.; Li, P.; Chen, N.; Geng, D.; Zhang, X.; et al. Prediction of Atorvastatin Pharmacokinetics in High-Fat Diet and Low-Dose Streptozotocin-Induced Diabetic Rats Using a Semiphysiologically Based Pharmacokinetic Model Involving Both Enzymes and Transporters. Drug Metab. Dispos. 2019, 47, 1066–1079. [Google Scholar] [CrossRef]
- Yang, H.; Su, M.; Liu, M.; Sheng, Y.; Zhu, L.; Yang, L.; Mu, R.; Zou, J.; Liu, X.; Liu, L. Hepatic retinaldehyde deficiency is involved in diabetes deterioration by enhancing PCK1- and G6PC-mediated gluconeogenesis. Acta Pharm. Sin. B 2023, 13, 3728–3743. [Google Scholar] [CrossRef]
- Kong, W.; Sun, X.; Yu, S.; Liu, P.; Zheng, X.; Zhang, J.; Zhu, L.; Jiang, T.; Jin, M.; Gao, J.; et al. Bile duct ligation increased dopamine levels in the cerebral cortex of rats partly due to induction of tyrosine hydroxylase. Br. J. Pharmacol. 2023, 180, 1690–1709. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.C.; Stocker, S.L.; Danta, M.; Carland, J.E.; Kumar, S.S.; Liu, Z.; Greenfield, J.R.; Braithwaite, H.E.; Cheng, T.S.; Graham, G.G.; et al. The safety and pharmacokinetics of metformin in patients with chronic liver disease. Aliment. Pharmacol. Ther. 2020, 51, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Brockmöller, J.; Seitz, T.; König, J.; Chen, X.; Tzvetkov, M.V. Tropane alkaloids as substrates and inhibitors of human organic cation transporters of the SLC22 (OCT) and the SLC47 (MATE) families. Biol. Chem. 2017, 398, 237–249. [Google Scholar] [CrossRef]
- Westphal, J.F.; Brogard, J.M. Drug administration in chronic liver disease. Drug Saf. 1997, 17, 47–73. [Google Scholar] [CrossRef] [PubMed]
- González, G.; Arancibia, A.; Rivas, M.I.; Caro, P.; Antezana, C. Pharmacokinetics of furosemide in patients with hepatic cirrhosis. Eur. J. Clin. Pharmacol. 1982, 22, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Marcantonio, L.A.; Auld, W.H.; Murdoch, W.R.; Purohit, R.; Skellern, G.G.; Howes, C.A. The pharmacokinetics and pharmacodynamics of the diuretic bumetanide in hepatic and renal disease. Br. J. Clin. Pharmacol. 1983, 15, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Cello, J.P.; Oie, S. Cimetidine disposition in patients with Laennec’s cirrhosis during multiple dosing therapy. Eur. J. Clin. Pharmacol. 1983, 25, 223–229. [Google Scholar] [CrossRef]
- Wang, M.X.; Liu, Y.L.; Yang, Y.; Zhang, D.M.; Kong, L.D. Nuciferine restores potassium oxonate-induced hyperuricemia and kidney inflammation in mice. Eur. J. Pharmacol. 2015, 747, 59–70. [Google Scholar] [CrossRef]
- Ryan, M.J.; Johnson, G.; Kirk, J.; Fuerstenberg, S.M.; Zager, R.A.; Torok-Storb, B. HK-2: An immortalized proximal tubule epithelial cell line from normal adult human kidney. Kidney Int. 1994, 45, 48–57. [Google Scholar] [CrossRef]
- Chen, S.Q.; Hu, B.F.; Yang, Y.R.; He, Y.; Yue, L.; Guo, D.; Wu, T.N.; Feng, X.W.; Li, Q.; Zhang, W.; et al. The protective effect of rabeprazole on cisplatin-induced apoptosis and necroptosis of renal proximal tubular cells. Biochem. Biophys. Res. Commun. 2022, 612, 91–98. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, S.; Hu, T.; Qu, X.; Zhai, J.; Zhang, Y.; Tao, L.; Yin, J.; Song, Y. Omeprazole protects against cisplatin-induced nephrotoxicity by alleviating oxidative stress, inflammation, and transporter-mediated cisplatin accumulation in rats and HK-2 cells. Chem. Biol. Interact. 2019, 297, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Abo El-Magd, N.F.; El-Kashef, D.H.; El-Sherbiny, M.; Eraky, S.M. Hepatoprotective and cognitive-enhancing effects of hesperidin against thioacetamide-induced hepatic encephalopathy in rats. Life Sci. 2023, 313, 121280. [Google Scholar] [CrossRef]
- Kobayashi, H.; Yoshida, S.; Sun, Y.J.; Shirasawa, N.; Naito, A. Gastric estradiol-17beta (E2) and liver ERalpha correlate with serum E2 in the cholestatic male rat. J. Endocrinol. 2013, 219, 39–49. [Google Scholar] [CrossRef]
- Wang, H.; Che, J.; Cui, K.; Zhuang, W.; Li, H.; Sun, J.; Chen, J.; Wang, C. Schisantherin A ameliorates liver fibrosis through TGF-β1mediated activation of TAK1/MAPK and NF-κB pathways in vitro and in vivo. Phytomedicine Int. J. Phytother. Phytopharm. 2021, 88, 153609. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, A.M.H.; Younis, N.S.; Metwaly, H.A. Vanillin augments liver regeneration effectively in Thioacetamide induced liver fibrosis rat model. Life Sci. 2021, 286, 120036. [Google Scholar] [CrossRef] [PubMed]
- El-Marasy, S.A.; El Awdan, S.A.; Abd-Elsalam, R.M. Protective role of chrysin on thioacetamide-induced hepatic encephalopathy in rats. Chem. Biol. Interact. 2019, 299, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Belenguer, G.; Duart-Abadia, P.; Jordán-Pla, A.; Domingo-Muelas, A.; Blasco-Chamarro, L.; Ferrón, S.R.; Morante-Redolat, J.M.; Fariñas, I. Adult Neural Stem Cells Are Alerted by Systemic Inflammation through TNF-α Receptor Signaling. Cell Stem Cell 2021, 28, 285–299.e9. [Google Scholar] [CrossRef]
- Chakraborty, B.; Byemerwa, J.; Krebs, T.; Lim, F.; Chang, C.Y.; McDonnell, D.P. Estrogen Receptor Signaling in the Immune System. Endocr. Rev. 2023, 44, 117–141. [Google Scholar] [CrossRef]
- Yang, D.; Xie, P.; Guo, S.; Li, H. Induction of MAPK phosphatase-1 by hypothermia inhibits TNF-alpha-induced endothelial barrier dysfunction and apoptosis. Cardiovasc. Res. 2010, 85, 520–529. [Google Scholar] [CrossRef]
- Yamashita, M.; Passegué, E. TNF-α Coordinates Hematopoietic Stem Cell Survival and Myeloid Regeneration. Cell Stem Cell 2019, 25, 357–372.e7. [Google Scholar] [CrossRef]
- Das, M.; Sabio, G.; Jiang, F.; Rincón, M.; Flavell, R.A.; Davis, R.J. Induction of hepatitis by JNK-mediated expression of TNF-alpha. Cell 2009, 136, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Qin, Z.; Wan, J.J.; Wang, P.Y.; Yang, Y.L.; Yu, J.G.; Hu, B.H.; Su, D.F.; Luo, Z.M.; Liu, X. Estrogen weakens muscle endurance via estrogen receptor-p38 MAPK-mediated orosomucoid (ORM) suppression. Exp. Mol. Med. 2018, 50, e463. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | CON | ALI |
|---|---|---|
| Body weight (BW) (g) | 236.71 ± 15.85 | 216.40 ± 6.29 ** |
| Liver weight (% BW) | 2.75 ± 0.17 | 4.18 ± 0.24 ** |
| Spleen weight (% BW) | 0.25 ± 0.04 | 0.22 ± 0.03 |
| ALT (IU/L) | 20.35 ± 13.51 | 88.08 ± 38.28 ** |
| AST (IU/L) | 10.21 ± 6.90 | 39.72 ± 2.65 ** |
| TBL (μmol/L) | ND | 3.61 ± 1.19 ** |
| Serum ammonia (μmol/L) | 182.32 ± 13.26 | 322.75 ± 46.26 ** |
| Estrogen (pg/mL) | 27.06 ± 2.93 | 86.42 ± 38.42 ** |
| TNF-α (pg/mL) | 5.82 ± 2.17 | 165.23 ± 32.00 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhi, H.; Dai, Y.; Su, L.; Yang, L.; Wu, W.; Wang, Z.; Zhu, X.; Liu, L.; Aa, J.; Yang, H. Thioacetamide-Induced Acute Liver Injury Increases Metformin Plasma Exposure by Downregulating Renal OCT2 and MATE1 Expression and Function. Biomedicines 2023, 11, 3314. https://doi.org/10.3390/biomedicines11123314
Zhi H, Dai Y, Su L, Yang L, Wu W, Wang Z, Zhu X, Liu L, Aa J, Yang H. Thioacetamide-Induced Acute Liver Injury Increases Metformin Plasma Exposure by Downregulating Renal OCT2 and MATE1 Expression and Function. Biomedicines. 2023; 11(12):3314. https://doi.org/10.3390/biomedicines11123314
Chicago/Turabian StyleZhi, Hao, Yidong Dai, Lin Su, Lu Yang, Wenhan Wu, Zehua Wang, Xinyue Zhu, Li Liu, Jiye Aa, and Hanyu Yang. 2023. "Thioacetamide-Induced Acute Liver Injury Increases Metformin Plasma Exposure by Downregulating Renal OCT2 and MATE1 Expression and Function" Biomedicines 11, no. 12: 3314. https://doi.org/10.3390/biomedicines11123314