

Rapid and Accurate Discrimination of Mycobacterium abscessus Subspecies Based on Matrix-Assisted Laser Desorption Ionization-Time of Flight Spectrum and Machine Learning Algorithms

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Protocol
2.2. Sample Preparation
2.3. Measurement and Preprocessing of MALDI–TOF Spectra
2.4. Determination of MABC Subspecies
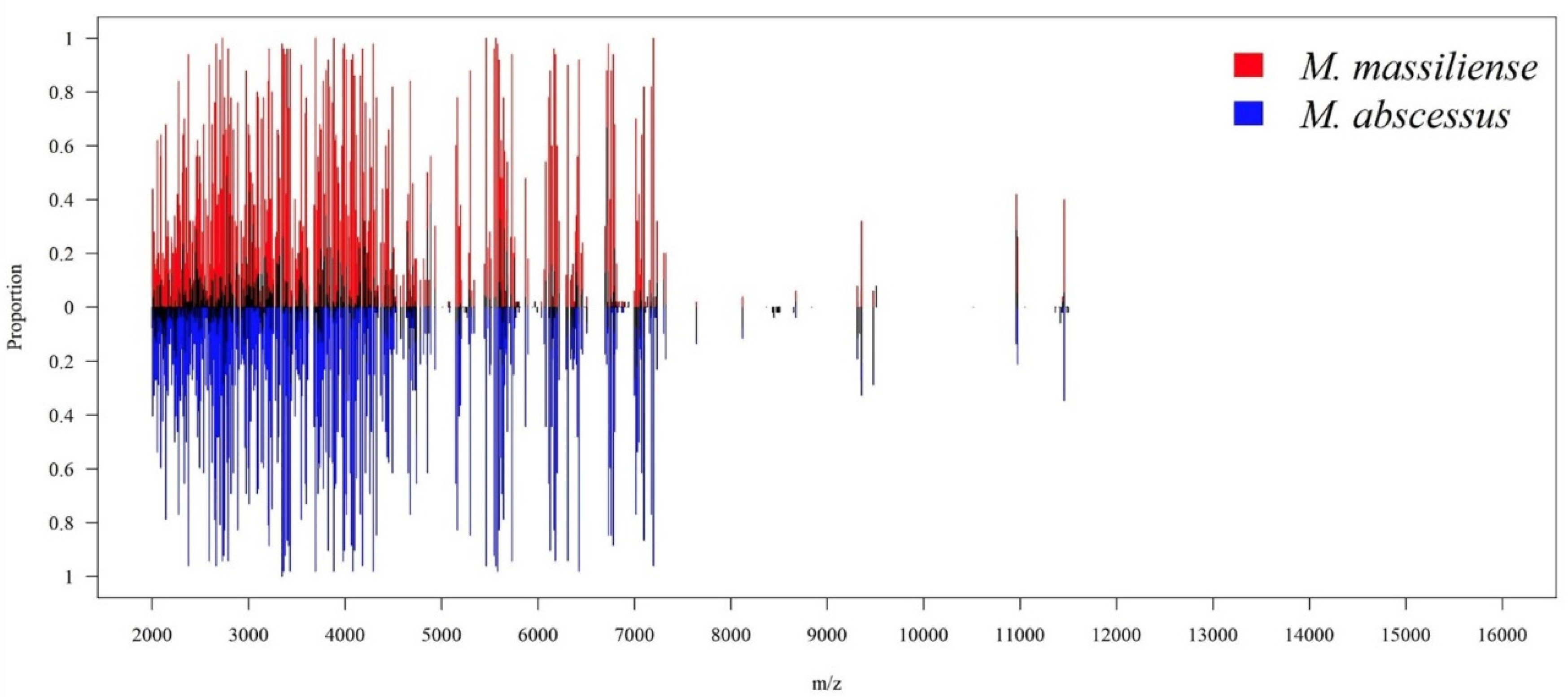
2.5. Visual Illustration of MS Spectrum Data onto a Two-Dimensional Plot
2.6. Feature Selection and Predictive Model Construction by ML Method
2.7. Model Performance Evaluation
3. Results
3.1. Bacterial Isolates and MALDI–TOF MS Spectra
3.2. Performance of Constructed Models
3.3. Discriminative Peaks
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jankovic, M.; Sabol, I.; Zmak, L.; Sola, A.-M.; Bulat-Kardum, L.; Popovic Grle, S.; Ticac, B.; Jakopovic, M.; Vukic Dugac, A.; Hecimovic, A.; et al. 5-year survival of patients with nontuberculous mycobacteria pulmonary disease (NTM-PD)–data from Croatian national registry. Eur. Respir. J. 2017, 50, PA2733. [Google Scholar] [CrossRef] [Green Version]
- Park, S.C.; Kang, M.J.; Han, C.H.; Lee, S.M.; Kim, C.J.; Lee, J.M.; Kang, Y.A. Prevalence, incidence, and mortality of nontuberculous mycobacterial infection in Korea: A nationwide population-based study. BMC Pulm. Med. 2019, 19, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.-H.; Pan, S.-W.; Wang, S.-M.; Shu, C.-C.; Chang, C.-H. The Impact of Nontuberculous Mycobacteria Species on Mortality in Patients with Nontuberculous Mycobacterial Lung Disease. Front. Microbiol. 2022, 13, 9274. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Cheng, M.H.; Lu, P.L.; Shu, C.C.; Wang, J.Y.; Wang, J.T.; Chong, I.W.; Lee, L.N. Epidemiology and Predictors of NTM Pulmonary Infection in Taiwan—A Retrospective, Five-Year Multicenter Study. Sci. Rep. 2017, 7, 16300. [Google Scholar] [CrossRef] [Green Version]
- Koh, W.-J.; Jeong, B.-H.; Jeon, K.; Kim, S.-Y.; Park, K.U.; Park, H.Y.; Huh, H.J.; Ki, C.-S.; Lee, N.Y.; Lee, S.-H.; et al. Oral Macrolide Therapy Following Short-term Combination Antibiotic Treatment of Mycobacterium massiliense Lung Disease. Chest 2016, 150, 1211–1221. [Google Scholar] [CrossRef]
- Chien, J.Y.; Lai, C.C.; Sheng, W.H.; Yu, C.J.; Hsueh, P.R. Pulmonary infection and colonization with nontuberculous mycobacteria, Taiwan, 2000-2012. Emerg. Infect. Dis. 2014, 20, 1382–1385. [Google Scholar] [CrossRef] [Green Version]
- Zweijpfenning, S.M.H.; Ingen, J.V.; Hoefsloot, W. Geographic Distribution of Nontuberculous Mycobacteria Isolated from Clinical Specimens: A Systematic Review. Semin. Respir. Crit. Care Med. 2018, 39, 336–342. [Google Scholar] [CrossRef]
- Degiacomi, G.; Sammartino, J.C.; Chiarelli, L.R.; Riabova, O.; Makarov, V.; Pasca, M.R. Mycobacterium abscessus, an Emerging and Worrisome Pathogen among Cystic Fibrosis Patients. Int. J. Mol. Sci. 2019, 20, 5868. [Google Scholar] [CrossRef] [Green Version]
- Haworth, C.S.; Banks, J.; Capstick, T.; Fisher, A.J.; Gorsuch, T.; Laurenson, I.F.; Leitch, A.; Loebinger, M.R.; Milburn, H.J.; Nightingale, M.; et al. British Thoracic Society guidelines for the management of non-tuberculous mycobacterial pulmonary disease (NTM-PD). Thorax 2017, 72, ii1–ii64. [Google Scholar] [CrossRef] [Green Version]
- Choi, G.E.; Shin, S.J.; Won, C.J.; Min, K.N.; Oh, T.; Hahn, M.Y.; Lee, K.; Lee, S.H.; Daley, C.L.; Kim, S.; et al. Macrolide treatment for Mycobacterium abscessus and Mycobacterium massiliense infection and inducible resistance. Am. J. Respir. Crit. Care Med. 2012, 186, 917–925. [Google Scholar] [CrossRef]
- Koh, W.J.; Jeon, K.; Lee, N.Y.; Kim, B.J.; Kook, Y.H.; Lee, S.H.; Park, Y.K.; Kim, C.K.; Shin, S.J.; Huitt, G.A.; et al. Clinical significance of differentiation of Mycobacterium massiliense from Mycobacterium abscessus. Am. J. Respir. Crit. Care Med. 2011, 183, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Shallom, S.J.; Gardina, P.J.; Myers, T.G.; Sebastian, Y.; Conville, P.; Calhoun, L.B.; Tettelin, H.; Olivier, K.N.; Uzel, G.; Sampaio, E.P.; et al. New rapid scheme for distinguishing the subspecies of the Mycobacterium abscessus group and identifying Mycobacterium massiliense isolates with inducible clarithromycin resistance. J. Clin. Microbiol. 2013, 51, 2943–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassi, M.; Drancourt, M. Genome analysis reveals three genomospecies in Mycobacterium abscessus. BMC Genom. 2014, 15, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.C.; Sun, P.L.; Wu, T.L.; Wang, L.H.; Yang, C.H.; Chung, W.H.; Kuo, A.J.; Liu, T.P.; Lu, J.J.; Chiu, C.H.; et al. Antimicrobial resistance in Mycobacterium abscessus complex isolated from patients with skin and soft tissue infections at a tertiary teaching hospital in Taiwan. J. Antimicrob. Chemother. 2017, 72, 2782–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benwill, J.L.; Wallace, R.J., Jr. Mycobacterium abscessus: Challenges in diagnosis and treatment. Curr. Opin. Infect. Dis. 2014, 27, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Weis, C.V.; Jutzeler, C.R.; Borgwardt, K. Machine learning for microbial identification and antimicrobial susceptibility testing on MALDI–TOF mass spectra: A systematic review. Clin. Microbiol. Infect. 2020, 26, 1310–1317. [Google Scholar] [CrossRef]
- Teng, S.H.; Chen, C.M.; Lee, M.R.; Lee, T.F.; Chien, K.Y.; Teng, L.J.; Hsueh, P.R. Matrix-assisted laser desorption ionization-time of flight mass spectrometry can accurately differentiate between Mycobacterium masilliense (M. abscessus subspecies bolletti) and M. abscessus (sensu stricto). J. Clin. Microbiol. 2013, 51, 3113–3116. [Google Scholar] [CrossRef] [Green Version]
- Mediavilla-Gradolph, M.C.; De Toro-Peinado, I.; Bermúdez-Ruiz, M.P.; García-Martínez Mde, L.; Ortega-Torres, M.; Montiel Quezel-Guerraz, N.; Palop-Borrás, B. Use of MALDI–TOF MS for Identification of Nontuberculous Mycobacterium Species Isolated from Clinical Specimens. Biomed Res. Int. 2015, 2015, 854078. [Google Scholar] [CrossRef] [Green Version]
- Mather, C.A.; Werth, B.J.; Sivagnanam, S.; SenGupta, D.J.; Butler-Wu, S.M. Rapid Detection of Vancomycin-Intermediate Staphylococcus aureus by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2016, 54, 883–890. [Google Scholar] [CrossRef] [Green Version]
- Wolters, M.; Rohde, H.; Maier, T.; Belmar-Campos, C.; Franke, G.; Scherpe, S.; Aepfelbacher, M.; Christner, M. MALDI–TOF MS fingerprinting allows for discrimination of major methicillin-resistant Staphylococcus aureus lineages. Int. J. Med. Microbiol. 2011, 301, 64–68. [Google Scholar] [CrossRef]
- van Oosten, L.N.; Klein, C.D. Machine Learning in Mass Spectrometry: A MALDI–TOF MS Approach to Phenotypic Antibacterial Screening. J. Med. Chem. 2020, 63, 8849–8856. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-Y.; Chung, C.-R.; Tseng, Y.-J.; Huang, Y.-C.; Yu, J.-R.; Chen, C.-J.; Lin, T.-W.; Huang, W.-T.; Lee, T.-Y.; Horng, J.T.; et al. Effect of an Artificial Intelligence-assisted Antibiotic Susceptibility Test on Reducing the Mortality Rate of and Antibiotics Use in Patients with Bacteremia: A Prospective Observational Cohort Study. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Zelazny, A.M.; Root, J.M.; Shea, Y.R.; Colombo, R.E.; Shamputa, I.C.; Stock, F.; Conlan, S.; McNulty, S.; Brown-Elliott, B.A.; Wallace, R.J., Jr.; et al. Cohort study of molecular identification and typing of Mycobacterium abscessus, Mycobacterium massiliense, and Mycobacterium bolletii. J. Clin. Microbiol. 2009, 47, 1985–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuck, M.; Grélard, F.; Blanc, L.; Desbenoit, N. MALDI-MSI Towards Multimodal Imaging: Challenges and Perspectives. Front. Chem. 2022, 10, 904688. [Google Scholar] [CrossRef]
- Verbeeck, N.; Caprioli, R.M.; Van de Plas, R. Unsupervised machine learning for exploratory data analysis in imaging mass spectrometry. Mass Spectrom. Rev. 2020, 39, 245–291. [Google Scholar] [CrossRef] [Green Version]
- Rossel, S.; Martínez Arbizu, P. Effects of Sample Fixation on Specimen Identification in Biodiversity Assemblies Based on Proteomic Data (MALDI–TOF). Front. Mar. Sci. 2018, 5, 149. [Google Scholar] [CrossRef] [Green Version]
- Bradley, A.P. The use of the area under the ROC curve in the evaluation of machine learning algorithms. Pattern Recognit. 1997, 30, 1145–1159. [Google Scholar] [CrossRef] [Green Version]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Mase, A.; Yamaguchi, F.; Funaki, T.; Yamazaki, Y.; Shikama, Y.; Fukuchi, K. PCR amplification of the erm(41) gene can be used to predict the sensitivity of Mycobacterium abscessus complex strains to clarithromycin. Exp. Ther. Med. 2020, 19, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Nie, W.; Duan, H.; Huang, H.; Lu, Y.; Bi, D.; Chu, N. Species identification of Mycobacterium abscessus subsp. abscessus and Mycobacterium abscessus subsp. bolletii using rpoB and hsp65, and susceptibility testing to eight antibiotics. Int. J. Infect. Dis. 2014, 25, 170–174. [Google Scholar] [CrossRef]
- Cheng, A.; Sun, H.Y.; Tsai, Y.T.; Chang, S.Y.; Wu, U.I.; Hsueh, P.R.; Sheng, W.H.; Chen, Y.C.; Chang, S.C. Comparing the Utilities of Different Multilocus Sequence Typing Schemes for Identifying Outbreak Strains of Mycobacterium abscessus subsp. massiliense. J. Clin. Microbiol. 2019, 58, e01304-19. [Google Scholar] [CrossRef]
- Marras, S.A.E.; Chen, L.; Shashkina, E.; Davidson, R.M.; Strong, M.; Daley, C.L.; Kreiswirth, B.N. A Molecular-Beacon-Based Multiplex Real-Time PCR Assay to Distinguish Mycobacterium abscessus Subspecies and Determine Macrolide Susceptibility. J. Clin. Microbiol. 2021, 59, e0045521. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, Z.; Wang, H.Y.; Chung, C.R.; Horng, J.T.; Lu, J.J.; Lee, T.Y. Large-Scale Samples Based Rapid Detection of Ciprofloxacin Resistance in Klebsiella pneumoniae Using Machine Learning Methods. Front. Microbiol. 2022, 13, 827451. [Google Scholar] [CrossRef] [PubMed]
- Breiman, L. Bagging predictors. Mach. Learn. 1996, 24, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Dematheis, F.; Walter, M.C.; Lang, D.; Antwerpen, M.; Scholz, H.C.; Pfalzgraf, M.-T.; Mantel, E.; Hinz, C.; Wölfel, R.; Zange, S. Machine Learning Algorithms for Classification of MALDI–TOF MS Spectra from Phylogenetically Closely Related Species Brucella melitensis, Brucella abortus and Brucella suis. Microorganisms 2022, 10, 1658. [Google Scholar] [CrossRef]
- Panagea, T.; Pincus, D.H.; Grogono, D.; Jones, M.; Bryant, J.; Parkhill, J.; Floto, R.A.; Gilligan, P. Mycobacterium abscessus Complex Identification with Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2015, 53, 2355–2358. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Y.; Li, W.C.; Huang, K.Y.; Chung, C.R.; Horng, J.T.; Hsu, J.F.; Lu, J.J.; Lee, T.Y. Rapid classification of group B Streptococcus serotypes based on matrix-assisted laser desorption ionization-time of flight mass spectrometry and machine learning techniques. BMC Bioinform. 2019, 20, 703. [Google Scholar] [CrossRef] [Green Version]
- Weis, C.; Cuénod, A.; Rieck, B.; Dubuis, O.; Graf, S.; Lang, C.; Oberle, M.; Brackmann, M.; Søgaard, K.K.; Osthoff, M.; et al. Direct antimicrobial resistance prediction from clinical MALDI–TOF mass spectra using machine learning. Nat. Med. 2022, 28, 164–174. [Google Scholar] [CrossRef]
- Weis, C.; Horn, M.; Rieck, B.; Cuénod, A.; Egli, A.; Borgwardt, K. Topological and kernel-based microbial phenotype prediction from MALDI–TOF mass spectra. Bioinformatics 2020, 36, i30–i38. [Google Scholar] [CrossRef]
- Kehrmann, J.; Kurt, N.; Rueger, K.; Bange, F.C.; Buer, J. GenoType NTM-DR for Identifying Mycobacterium abscessus Subspecies and Determining Molecular Resistance. J. Clin. Microbiol. 2016, 54, 1653–1655. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LR | DT | RF | KNN | SVM | |
|---|---|---|---|---|---|
| SEN | 0.5037 (0.4896, 0.5179) | 0.8399 (0.8313, 0.8484) | 0.8655 (0.8582, 0.8727) | 0.6315 (0.6222, 0.6408) | 0.8015 (0.7929, 0.8101) |
| SPE | 0.4867 (0.4731, 0.5003) | 0.8627 (0.8537, 0.8718) | 0.8672 (0.86, 0.8744) | 0.6348 (0.6254, 0.6442) | 0.8048 (0.7965, 0.8132) |
| ACC | 0.4930 (0.4825, 0.5035) | 0.8513 (0.8438, 0.8589) | 0.8666 (0.8594, 0.8737) | 0.6329 (0.6237, 0.6421) | 0.8031 (0.7948, 0.8115) |
| AUROC | 0.5812 (0.5732, 0.5893) | 0.8909 (0.8845, 0.8974) | 0.9134 (0.9072, 0.9196) | 0.6873 (0.6781, 0.6965) | 0.8709 (0.8634, 0.8785) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-Y.; Kuo, C.-H.; Chung, C.-R.; Lin, W.-Y.; Wang, Y.-C.; Lin, T.-W.; Yu, J.-R.; Lu, J.-J.; Wu, T.-S. Rapid and Accurate Discrimination of Mycobacterium abscessus Subspecies Based on Matrix-Assisted Laser Desorption Ionization-Time of Flight Spectrum and Machine Learning Algorithms. Biomedicines 2023, 11, 45. https://doi.org/10.3390/biomedicines11010045
Wang H-Y, Kuo C-H, Chung C-R, Lin W-Y, Wang Y-C, Lin T-W, Yu J-R, Lu J-J, Wu T-S. Rapid and Accurate Discrimination of Mycobacterium abscessus Subspecies Based on Matrix-Assisted Laser Desorption Ionization-Time of Flight Spectrum and Machine Learning Algorithms. Biomedicines. 2023; 11(1):45. https://doi.org/10.3390/biomedicines11010045
Chicago/Turabian StyleWang, Hsin-Yao, Chi-Heng Kuo, Chia-Ru Chung, Wan-Ying Lin, Yu-Chiang Wang, Ting-Wei Lin, Jia-Ruei Yu, Jang-Jih Lu, and Ting-Shu Wu. 2023. "Rapid and Accurate Discrimination of Mycobacterium abscessus Subspecies Based on Matrix-Assisted Laser Desorption Ionization-Time of Flight Spectrum and Machine Learning Algorithms" Biomedicines 11, no. 1: 45. https://doi.org/10.3390/biomedicines11010045