
Improved Protocol to Study Osteoblast and Adipocyte Differentiation Balance
 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Literature Revision
2.3. Cell Culture and Differentiation
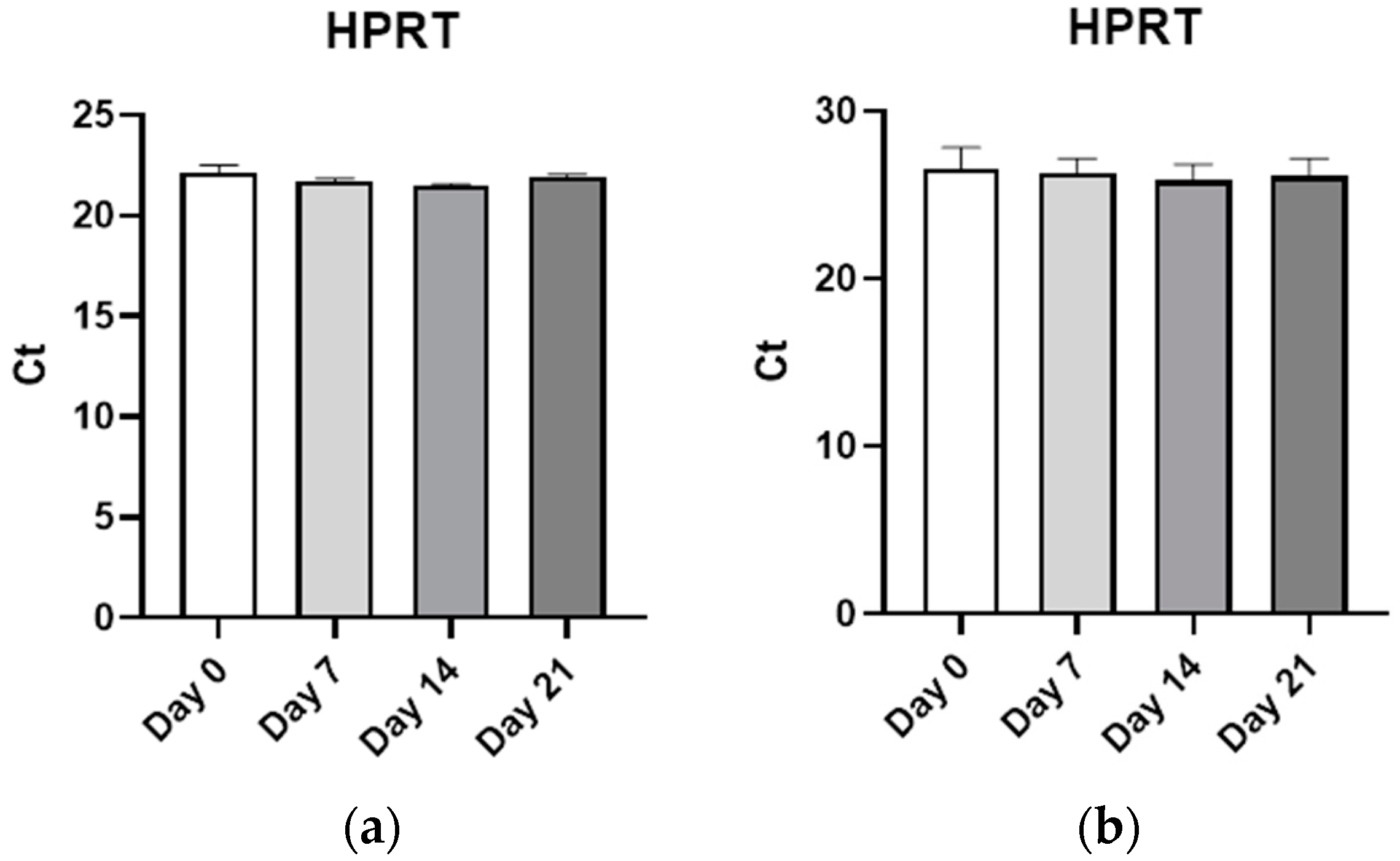
2.4. Gene Expression Analysis
2.5. Cellular Stainings
2.6. Statistical Analysis
3. Results
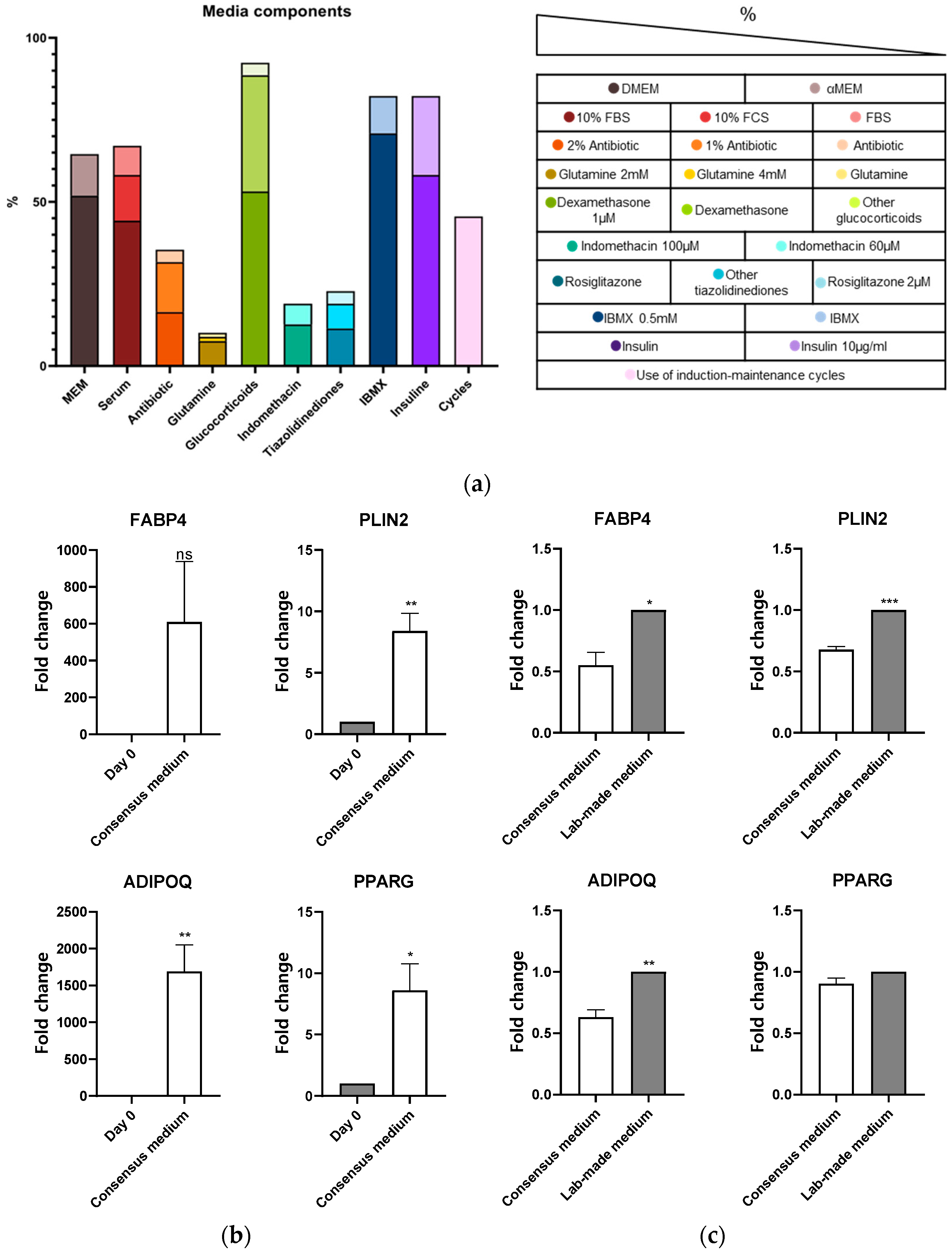
3.1. Adipogenic Consensus Medium
3.2. Osteoblastic Consensus Medium
3.3. Effect of Lab-Made Media vs. Commercial Media on Primary hMSCs
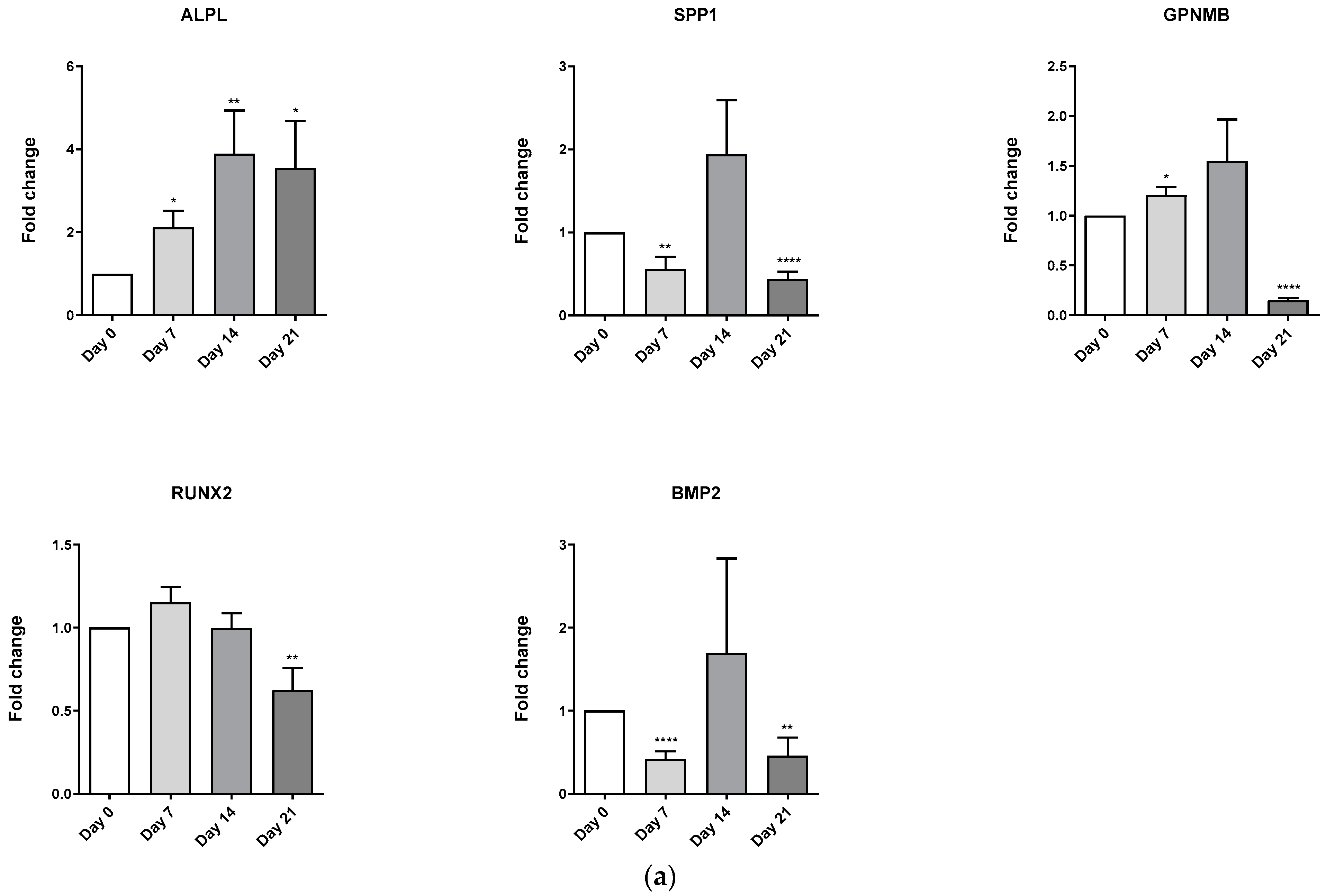
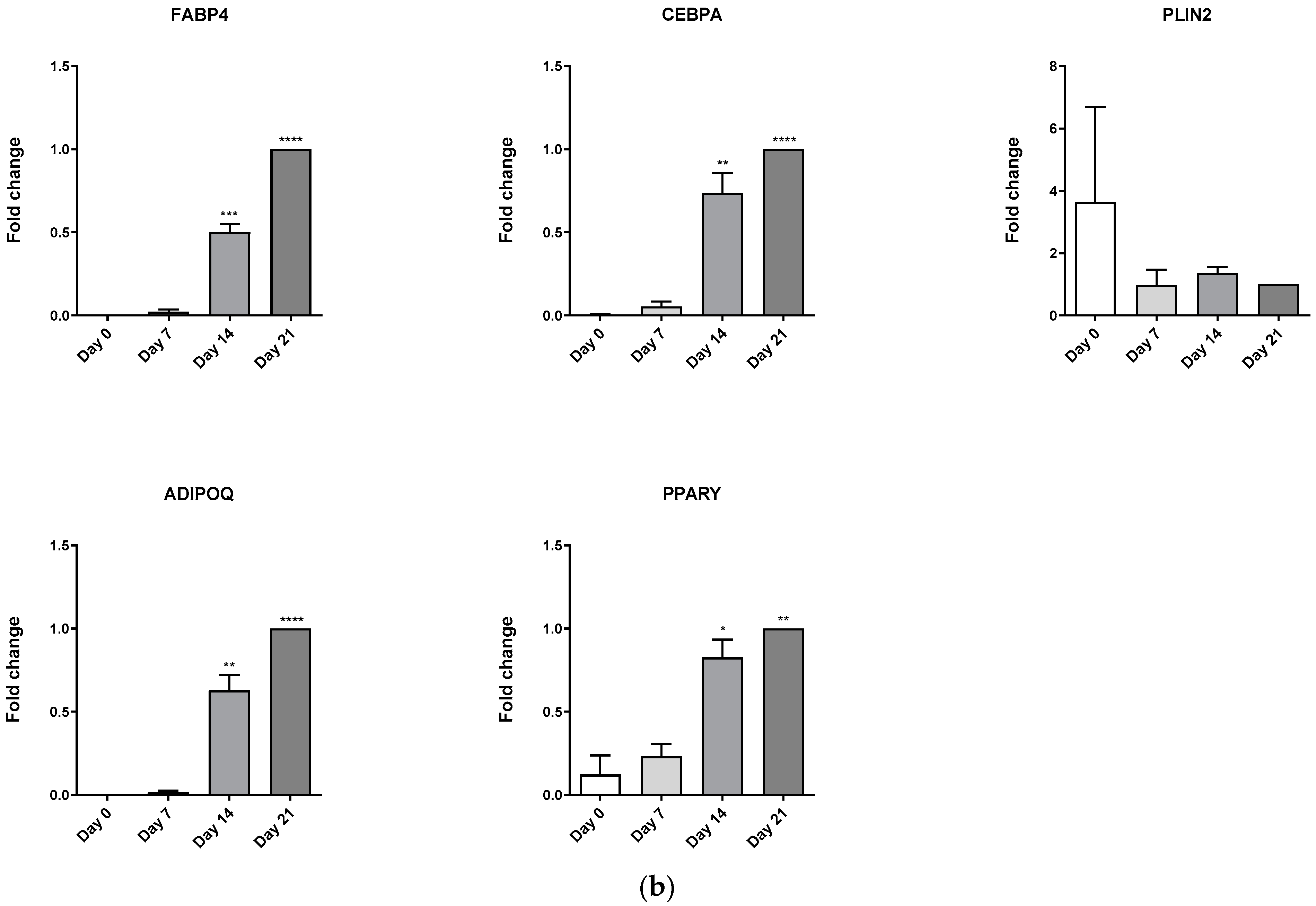
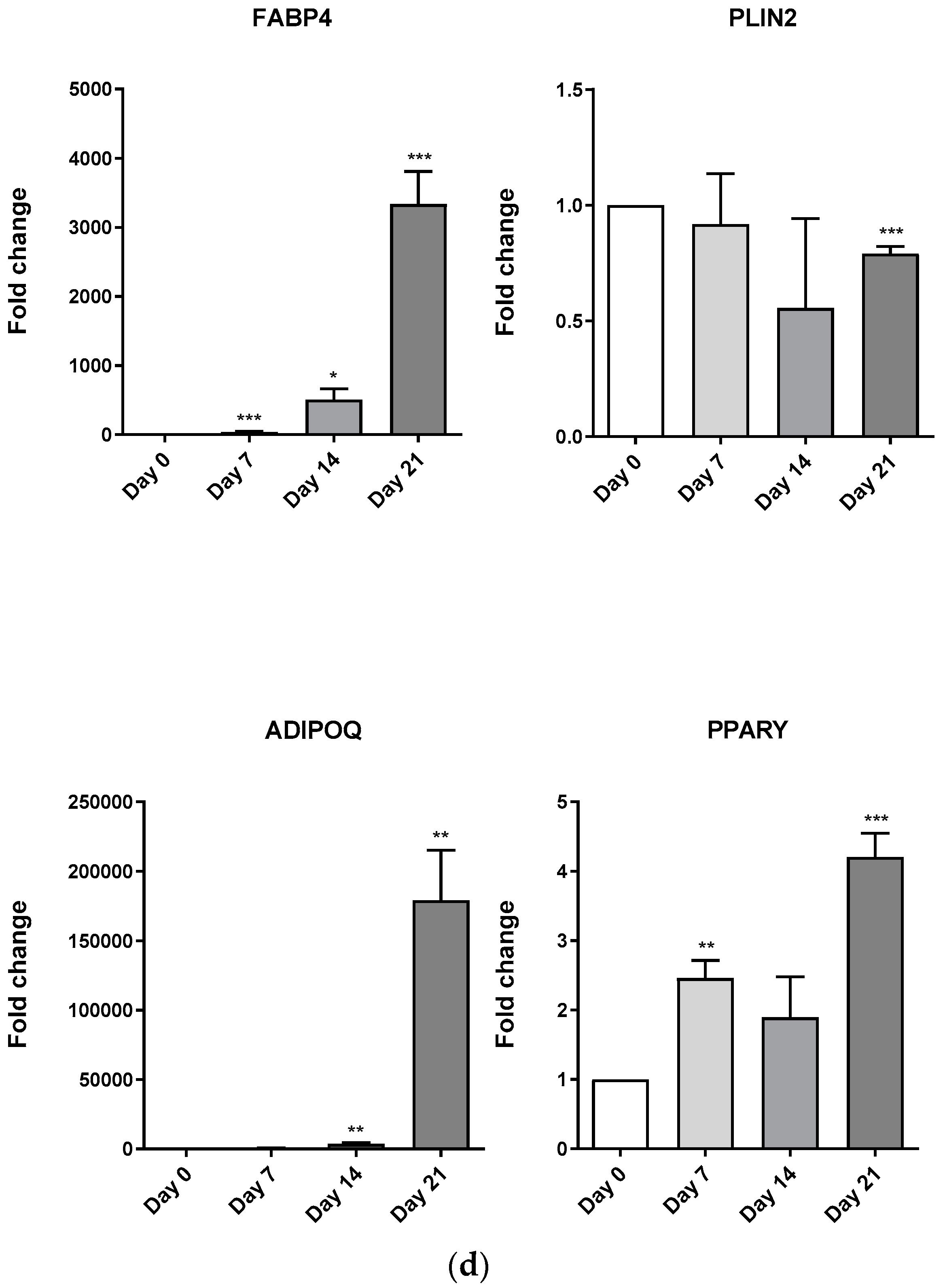
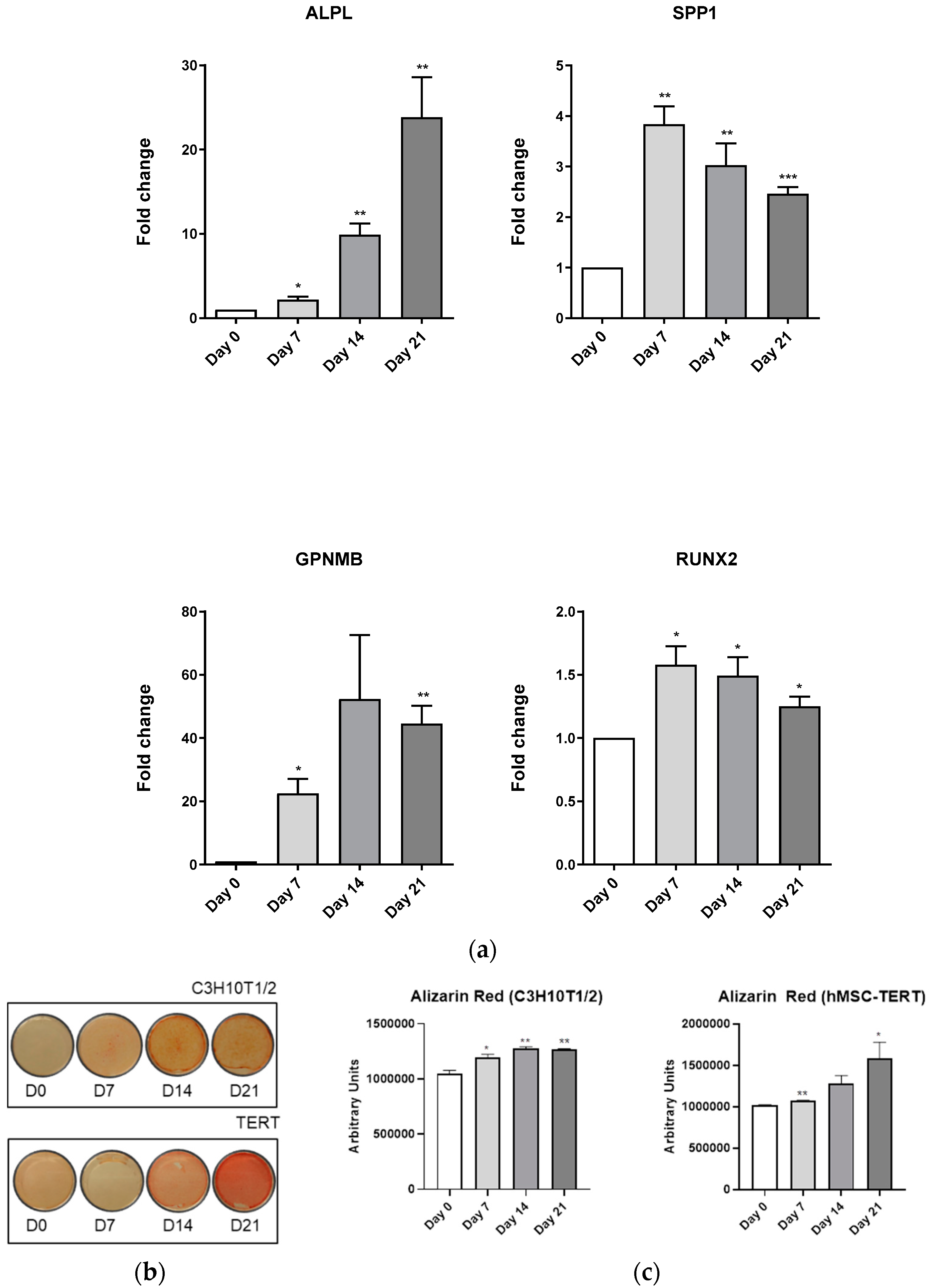
3.4. Evaluation of Osteoblast-Adipocyte Commitment of Primary Aged Osteoblast-like Cells
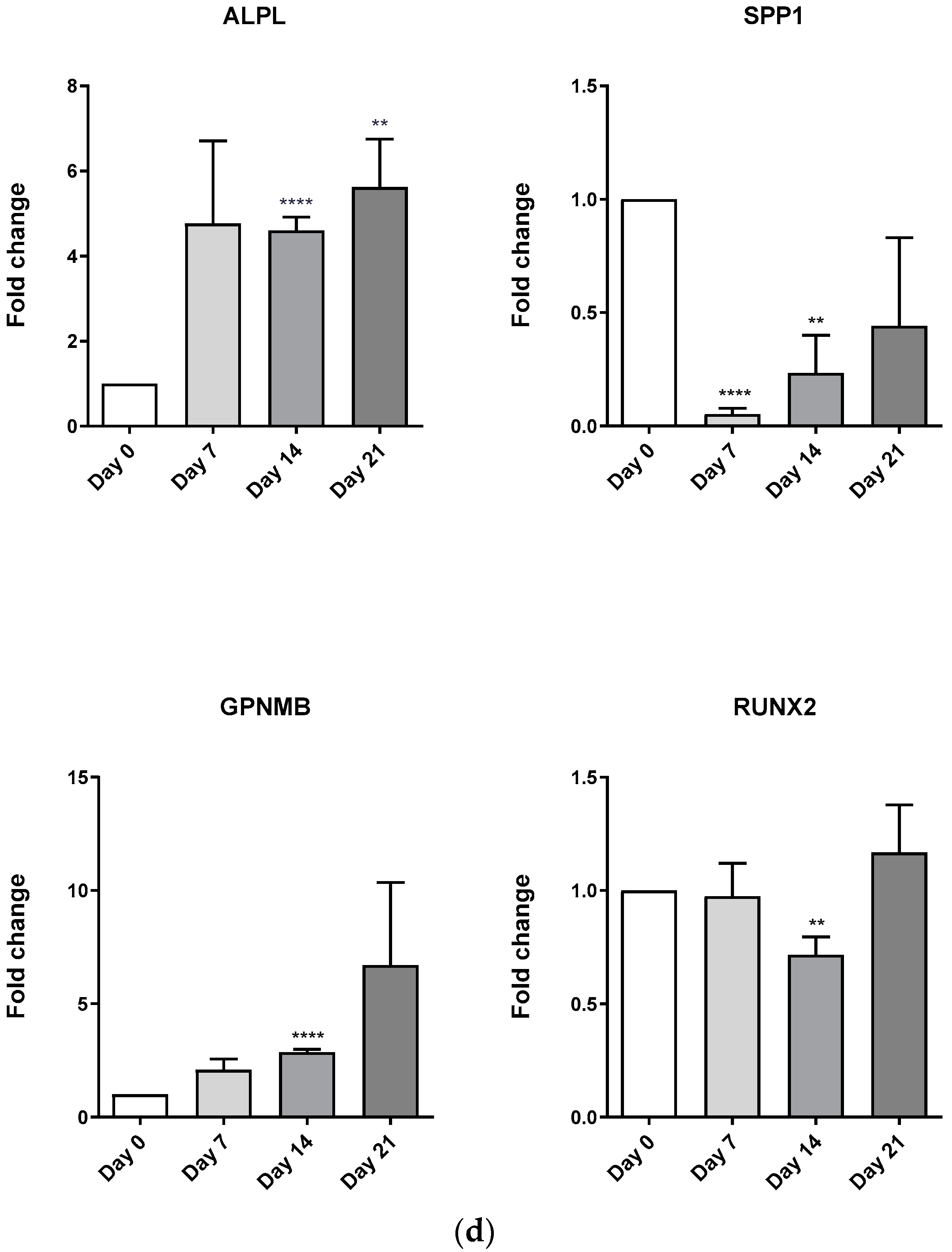
3.5. Differentiation Media Evaluation in MSC Lines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO|Chronic Rheumatic Conditions. WHO. 2016. Available online: http://www.who.int/chp/topics/rheumatic/en/ (accessed on 27 July 2020).
- Yamamoto, Y.; Turkiewicz, A.; Wingstrand, H.; Englund, M.; Englund, M. Fragility Fractures in Patients with Rheumatoid Arthritis and Osteoarthritis Compared with the General Population. J. Rheumatol. 2015, 42, 2055–2058. [Google Scholar] [CrossRef] [PubMed]
- Goldring, S.R.; Purdue, P.E.; Crotti, T.N.; Shen, Z.; Flannery, M.R.; Binder, N.B.; Ross, F.P.; McHugh, K.P.; Edward Purdue, P.; Crotti, T.N.; et al. Bone remodelling in inflammatory arthritis. Ann. Rheum. Dis. 2013, 72 (Suppl. 2), ii52–ii55. [Google Scholar] [CrossRef] [PubMed]
- Paskins, Z.; Whittle, R.; Abdul Sultan, A.; Muller, S.; Blagojevic-Bucknall, M.; Helliwell, T.; Packham, J.; Hider, S.; Roddy, E.; Mallen, C. Risk of fragility fracture among patients with late-onset psoriasis: A UK population-based study. Osteoporos. Int. 2018, 29, 1659–1664. [Google Scholar] [CrossRef]
- Ozdil, K.; Kahraman, R.; Sahin, A.; Calhan, T.; Gozden, E.H.; Akyuz, U.; Erer, B.; Sokmen, M.H. Bone density in proton pump inhibitors users: A prospective study. Rheumatol. Int. 2013, 33, 2255–2260. [Google Scholar] [CrossRef]
- Chiba, K.; Uetani, M.; Kido, Y.; Ito, M.; Okazaki, N.; Taguchi, K.; Shindo, H. Osteoporotic changes of subchondral trabecular bone in osteoarthritis of the knee: A 3-T MRI study. Osteoporos. Int. 2012, 23, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Justesen, J.; Stenderup, K.; Ebbesen, E.N.; Mosekilde, L.; Steiniche, T.; Kassem, M. Adipocyte tissue volume in bone marrow is increased with aging and in patients with osteoporosis. Biogerontology 2001, 2, 165–171. [Google Scholar] [CrossRef]
- Moerman, E.J.; Teng, K.; Lipschitz, D.A.; Lecka-Czernik, B. Aging activates adipogenic and suppresses osteogenic programs in mesenchymal marrow stroma/stem cells: The role of PPAR-γ 2 transcription factor and TGF-β/BMP signaling pathways. Aging Cell 2004, 3, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Atashi, F.; Modarressi, A.; Pepper, M.S. The Role of Reactive Oxygen Species in Mesenchymal Stem Cell Adipogenic and Osteogenic Differentiation: A Review. Stem Cells Dev. 2015, 24, 1150–1163. [Google Scholar] [CrossRef] [Green Version]
- Rosen, C.J.; Bouxsein, M.L. Mechanisms of disease: Is osteoporosis the obesity of bone? Nat. Clin. Pract. Rheumatol. 2006, 2, 35–43. [Google Scholar] [CrossRef]
- Takada, I.; Kouzmenko, A.P.; Kato, S. Wnt and PPARγ signaling in osteoblastogenesis and adipogenesis. Nat. Rev. Rheumatol. 2009, 5, 442–447. [Google Scholar] [CrossRef]
- Yang, D.-C.C.; Tsay, H.-J.J.; Lin, S.-Y.Y.; Chiou, S.-H.H.; Li, M.-J.J.; Chang, T.-J.J.; Hung, S.-C.C. cAMP/PKA Regulates Osteogenesis, Adipogenesis and Ratio of RANKL/OPG mRNA Expression in Mesenchymal Stem Cells by Suppressing Leptin. PLoS ONE 2008, 3, e1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulos, S.P.; Dodson, M.V.; Hausman, G.J. Cell line models for differentiation: Preadipocytes and adipocytes. Exp. Biol. Med. 2010, 235, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Czekanska, E.M.; Stoddart, M.J.; Richards, R.G.; Hayes, J.S. In search of an osteoblast cell model for in vitro research. Eur. Cells Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Helfrich, S.H.; Ralston, M.H. Bone Research Protocols, 1st ed.; Humana Press Inc.: Totowa, NJ, USA, 2003. [Google Scholar]
- Levi, B.; James, A.W.; Wan, D.C.; Glotzbach, J.P.; Commons, G.W.; Longaker, M.T. Regulation of Human Adipose-Derived Stromal Cell Osteogenic Differentiation by Insulin-like Growth Factor-1 and Platelet-Derived Growth Factor-Alpha. Plast. Reconstr. Surg. 2010, 126, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nueda, M.L.; García-Ramírez, J.J.; Laborda, J.; Baladrón, V. dlk1 Specifically Interacts with Insulin-Like Growth Factor Binding Protein 1 to Modulate Adipogenesis of 3T3-L1 Cells. J. Mol. Biol. 2008, 379, 428–442. [Google Scholar] [CrossRef]
- Zhao, P.; Deng, Y.; Gu, P.; Wang, Y.; Zhou, H.; Hu, Y.; Chen, P.; Fan, X. Insulin-like growth factor 1 promotes the proliferation and adipogenesis of orbital adipose-derived stromal cells in thyroid-associated ophthalmopathy. Exp. Eye Res. 2013, 107, 65–73. [Google Scholar] [CrossRef]
- MacDougald, O.A.; Lane, M.D. Transcriptional regulation of gene expression during adipocyte differentiation. Annu. Rev. Biochem. 1995, 64, 345–373. [Google Scholar] [CrossRef]
- Rharass, T.; Lucas, S. MECHANISMS IN ENDOCRINOLOGY: Bone marrow adiposity and bone, a bad romance? Eur. J. Endocrinol. 2018, 179, R165–R182. [Google Scholar] [CrossRef] [Green Version]
- Bellows, C.G.; Aubin, J.E.; Heersche, J.N.M. Initiation and progression of mineralization of bone nodules formed in vitro: The role of alkaline phosphatase and organic phosphate. Bone Miner. 1991, 14, 27–40. [Google Scholar] [CrossRef]
- Franceschi, R.T.; Iyer, B.S. Relationship between collagen synthesis and expression of the osteoblast phenotype in MC3T3-E1 cells. J. Bone Miner. Res. 1992, 7, 235–246. [Google Scholar] [CrossRef]
- Okazaki, K.; Yamaguchi, T.; Tanaka, K.I.; Notsu, M.; Ogawa, N.; Yano, S.; Sugimoto, T. Advanced glycation end products (AGEs), but not high glucose, inhibit the osteoblastic differentiation of mouse stromal ST2 cells through the suppression of osterix expression, and inhibit cell growth and increasing cell apoptosis. Calcif. Tissue Int. 2012, 91, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, C.S.; Johncola, A.J.; Batzdorf, A.S.; Jones, B.C.; Al Mukaddam, M.; Sexton, K.; Shults, J.; Leonard, M.B.; Snyder, P.J.; Wehrli, F.W. Effect of Low-Intensity Vibration on Bone Strength, Microstructure, and Adiposity in Pre-Osteoporotic Postmenopausal Women: A Randomized Placebo-Controlled Trial. J. Bone Miner. Res. 2021, 36, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Rauch, A.; Haakonsson, A.K.; Madsen, J.G.S.; Larsen, M.; Forss, I.; Madsen, M.R.; Van Hauwaert, E.L.; Wiwie, C.; Jespersen, N.Z.; Tencerova, M.; et al. Osteogenesis depends on commissioning of a network of stem cell transcription factors that act as repressors of adipogenesis. Nat. Genet. 2019, 51, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Ciuffreda, M.C.; Malpasso, G.; Musarò, P.; Turco, V.; Gnecchi, M. Protocols for in vitro differentiation of human mesenchymal stem cells into osteogenic, chondrogenic and adipogenic lineages. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2016; Volume 1416, pp. 149–158. [Google Scholar] [CrossRef]
- Reed, B.C.; Lane, M.D. Insulin receptor synthesis and turnover in differentiating 3T3-L1 preadipocytes. Proc. Natl. Acad. Sci. USA 1980, 77, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-J.J.; Fried, S.K. Optimal protocol for the differentiation and metabolic analysis of human adipose stromal cells. In Methods in Enzymology; Academic Press Inc.: Cambridge, MA, USA, 2014; Volume 538, pp. 49–65. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.-Q.; Otto, T.C.; Lane, M.D. Commitment of C3H10T1/2 pluripotent stem cells to the adipocyte lineage. Proc. Natl. Acad. Sci. USA 2004, 101, 9607–9611. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.B.; Benkusky, N.A.; Sen, B.; Rubin, J.; Pike, J.W. Epigenetic Plasticity Drives Adipogenic and Osteogenic Differentiation of Marrow-derived Mesenchymal Stem Cells. J. Biol. Chem. 2016, 291, 17829–17847. [Google Scholar] [CrossRef] [Green Version]
- Qian, S.-W.; Li, X.; Zhang, Y.-Y.; Huang, H.-Y.; Liu, Y.; Sun, X.; Tang, Q.-Q. Characterization of adipocyte differentiation from human mesenchymal stem cells in bone marrow. BMC Dev. Biol. 2010, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Hanna, H.; Mir, L.M.; Andre, F.M. In vitro osteoblastic differentiation of mesenchymal stem cells generates cell layers with distinct properties. Stem Cell Res. Ther. 2018, 9, 203. [Google Scholar] [CrossRef]
- Liu, B.; Lu, Y.; Wang, Y.; Ge, L.; Zhai, N.; Han, J. A protocol for isolation and identification and comparative characterization of primary osteoblasts from mouse and rat calvaria. Cell Tissue Bank. 2019, 20, 173. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. TGF-β/BMP signaling and other molecular events: Regulation of osteoblastogenesis and bone formation. Bone Res. 2015, 3, 15005. [Google Scholar] [CrossRef] [Green Version]
- Reddi, A.H. Cell Biology and Biochemistry of Endochondral Bone Development. Coll. Relat. Res. 1981, 1, 209–226. [Google Scholar] [CrossRef] [PubMed]
- Date, T.; Doiguchi, Y.; Nobuta, M.; Shindo, H. Bone morphogenetic protein-2 induces differentiation of multipotent C3H10T1/2 cells into osteoblasts, chondrocytes, and adipocytes in vivo and in vitro. J. Orthop. Sci. 2004, 9, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Liu, W.; Kuang, S. Fatty acid binding protein 4 expression marks a population of adipocyte progenitors in white and brown adipose tissues. FASEB J. 2013, 27, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heid, H.W.; Molí, H.; Schwetlick, I.; Rackwitz´thomas, H.-R.; Rackwitz´thomas, R.; Keenan, W.; Heid, H.W.; Rackwitz, H.-R.; Moll, R.; Schwetlick, I.; et al. Adipophilin is a specific marker of lipid accumulation in diverse cell types and diseases. Cell Tissue Res. 1998, 294, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, C.; Gao, Y.; Li, F.; Zhang, Y.; Dai, L.; Jiang, H.; Liu, H.; Zhang, S. Silencing of ADIPOQ Efficiently Suppresses Preadipocyte Differentiation in Porcine. Cell Physiol. Biochem. 2013, 31, 452–461. [Google Scholar] [CrossRef]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPARγ is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Murshed, M.; Harmey, D.; Millán, J.L.; Mckee, M.D.; Karsenty, G. Unique coexpression in osteoblasts of broadly expressed genes accounts for the spatial restriction of ECM mineralization to bone. GENES Dev. 2005, 19, 1093–1104. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.A.R.; Tye, C.E.; Sampaio, A.V.; Underhill, T.M.; Hunter, G.K.; Goldberg, H.A. Bone sialoprotein expression enhances osteoblast differentiation and matrix mineralization in vitro. Bone 2007, 41, 462–473. [Google Scholar] [CrossRef]
- Abdelmagid, S.M.; Belcher, J.Y.; Moussa, F.M.; Lababidi, S.L.; Sondag, G.R.; Novak, K.M.; Sanyurah, A.S.; Frara, N.A.; Razmpour, R.; Del Carpio-Cano, F.E.; et al. Mutation in Osteoactivin Decreases Bone Formation in Vivo and Osteoblast Differentiation in Vitro. Am. J. Pathol. 2014, 184, 697. [Google Scholar] [CrossRef] [Green Version]
- Rawadi, G.; Atrice Vayssière, B.É.; Vayssière, V.; Dunn, F.; Baron, R.; Roman-Roman, S. BMP-2 Controls Alkaline Phosphatase Expression and Osteoblast Mineralization by a Wnt Autocrine Loop. J. Bone Miner. Res. 2003, 18, 1842–1853. [Google Scholar] [CrossRef]
- Baniwal, S.K.; Shah, P.K.; Shi, Y.; Haduong, J.H.; Declerck, Y.A.; Gabet, Y.; Frenkel, B. Runx2 promotes both osteoblastogenesis and novel osteoclastogenic signals in ST2 mesenchymal progenitor cells. Osteoporos. Int. 2012, 23, 1399–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gossiel, F.; Altaher, H.; Reid, D.M.; Roux, C.; Felsenberg, D.; Glüer, C.C.; Eastell, R. Bone turnover markers after the menopause: T-score approach. Bone 2018, 111, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, B.M.; Haack-Sørensen, M.; Burns, J.S.; Elsnab, B.; Jakob, F.; Hokland, P.; Kassem, M. Maintenance of differentiation potential of human bone marrow mesenchymal stem cells immortalized by human telomerase reverse transcriptase gene despite of extensive proliferation. Biochem. Biophys. Res. Commun. 2005, 326, 527–538. [Google Scholar] [CrossRef]
- Altmann, S.; Mut, J.; Wolf, N.; Meißner-Weigl, J.; Rudert, M.; Jakob, F.; Gutmann, M.; Lühmann, T.; Seibel, J.; Ebert, R. Metabolic glycoengineering in hmsc-tert as a model for skeletal precursors by using modified azide/alkyne monosaccharides. Int. J. Mol. Sci. 2021, 22, 2820. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yu, Q.; Lai, A.; Tian, J. Pulsed electromagnetic field induces Ca2+-dependent osteoblastogenesis in C3H10T1/2 mesenchymal cells through the Wnt-Ca2+/Wnt-β-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2018, 503, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xing, J.; Hu, X.; Chen, L.; Lv, H.; Xu, C.; Hong, D.; Wu, X. Inhibition of heat shock protein 90 rescues glucocorticoid-induced bone loss through enhancing bone formation. J. Steroid Biochem. Mol. Biol. 2017, 171, 236–246. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Symbol | Forward Primer (5′-3′) Sequence | Reverse Primer (5′-3′) Sequence |
|---|---|---|---|
| Human Alkaline phosphatase | ALPL | TCTTCACATTTGGTGGATAC | ATGGAGACATTCTCTCGTTC |
| Human Osteopontin | SPP1 | GACCAAGGAAAACTCACTAC | CTGTTTAACTGGTATGGCAC |
| Human Osteoactivin | GPNMB | CAGATCAGATTCCTGTGTTTG | ACAGTATGATTGGTGGAAAC |
| Human RUNT-related transcription factor 2 | RUNX2 | AAGCTTGATGACTCTAAACC | TCTGTAATCTGACTCTGTCC |
| Human Bone morphogenic protein 2 | BMP2 | TCCACCATGAAGAATCTTTG | TAATTCGGTGATGGAAACTG |
| Human Fatty acid binding protein 4 | FABP4 | CAAGAGCACCATAACCTTAG | CTCGTTTTCTCTTTATGGTGG |
| Human CCAAT Enhancer Binding Protein Alpha | CEBPA | AGCCTTGTTTGTACTGTATG | AAAATGGTGGTTTAGCAGAG |
| Human Perilipin 2 | PLIN2 | GTTCACCTGATTGAATTTGC | GAGGTAGAGCTTATCCTGAG |
| Human Adiponectin | ADIPOQ | GGTCTTATTGGTCCTAAGGG | GTAGAAGATCTTGGTAAAGCG |
| Human Peroxisome proliferator activator receptor γ | PPARG | AAAGAAGCCAACACTAAACC | TGGTCATTTCGTTAAAGGC |
| Human Hypoxanthine Phosphoribosyl transferase 1 | HPRT | ATAAGCCAGACTTTGTTGG | ATAGGACTCCAGATGTTTCC |
| Mouse Alkaline phosphatase | ALPL | ATTCCCACTATGTCTGGAAC | CTCAAAGAGACCTAAGAGGTAG |
| Mouse Osteopontin | SPP1 | GGATGAATCTGACGAATCTC | GCATCAGGATACTGTTCATC |
| Mouse Osteoactivin | GPNMB | CTCTTTAATGCCTACTGGTTAC | GCCATATCTGTTTATTCGGC |
| Mouse RUNT-related transcription factor 2 | RUNX2 | ACAAGGACAGAGTCAGATTAC | CAGTGTCATCATCTGAAATACG |
| Mouse Fatty acid binding protein 4 | FABP4 | GTAAATGGGGATTTGGTCAC | TATGATGCTCTTCACCTTCC |
| Mouse Perilipin 2 | PLIN2 | ATAAGCTCTATGTCTCGTGG | GCCTGATCTTGAATGTTCTG |
| Mouse Adiponectin | ADIPOQ | CCACTTTCTCCTCATTTCTG | CTAGCTCTTCAGTTGTAGTAAC |
| Mouse Peroxisome proliferator activator receptor γ | PPARG | AAAGACAACGGACAAATCAC | GGGATATTTTTGGCATACTCTG |
| Mouse Hypoxanthine Phosphoribosyl transferase 1 | HPRT | AGGGATTTGAATCACGTTTG | TTTACTGGCAACATCAACAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso-Pérez, A.; Guillán-Fresco, M.; Franco-Trepat, E.; Jorge-Mora, A.; López-Fagúndez, M.; Pazos-Pérez, A.; Crespo-Golmar, A.; Caeiro-Rey, J.R.; Gómez, R. Improved Protocol to Study Osteoblast and Adipocyte Differentiation Balance. Biomedicines 2023, 11, 31. https://doi.org/10.3390/biomedicines11010031
Alonso-Pérez A, Guillán-Fresco M, Franco-Trepat E, Jorge-Mora A, López-Fagúndez M, Pazos-Pérez A, Crespo-Golmar A, Caeiro-Rey JR, Gómez R. Improved Protocol to Study Osteoblast and Adipocyte Differentiation Balance. Biomedicines. 2023; 11(1):31. https://doi.org/10.3390/biomedicines11010031
Chicago/Turabian StyleAlonso-Pérez, Ana, María Guillán-Fresco, Eloi Franco-Trepat, Alberto Jorge-Mora, Miriam López-Fagúndez, Andrés Pazos-Pérez, Antía Crespo-Golmar, José R. Caeiro-Rey, and Rodolfo Gómez. 2023. "Improved Protocol to Study Osteoblast and Adipocyte Differentiation Balance" Biomedicines 11, no. 1: 31. https://doi.org/10.3390/biomedicines11010031