
Biofilms on Indwelling Artificial Urinary Sphincter Devices Harbor Complex Microbe–Metabolite Interaction Networks and Reconstitute Differentially In Vitro by Material Type
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Demographics
2.2. Sample Processing
2.3. Sequencing
2.4. Antibiotic Resistance and Biofilm Gene Detection
2.5. Metabolomics
2.6. Bioinformatics
2.7. Continuous-Flow Stir Tank Bioreactor Biofilm Assays and Scanning Electron Microscopy
3. Results
3.1. Biofilm Microbial Composition
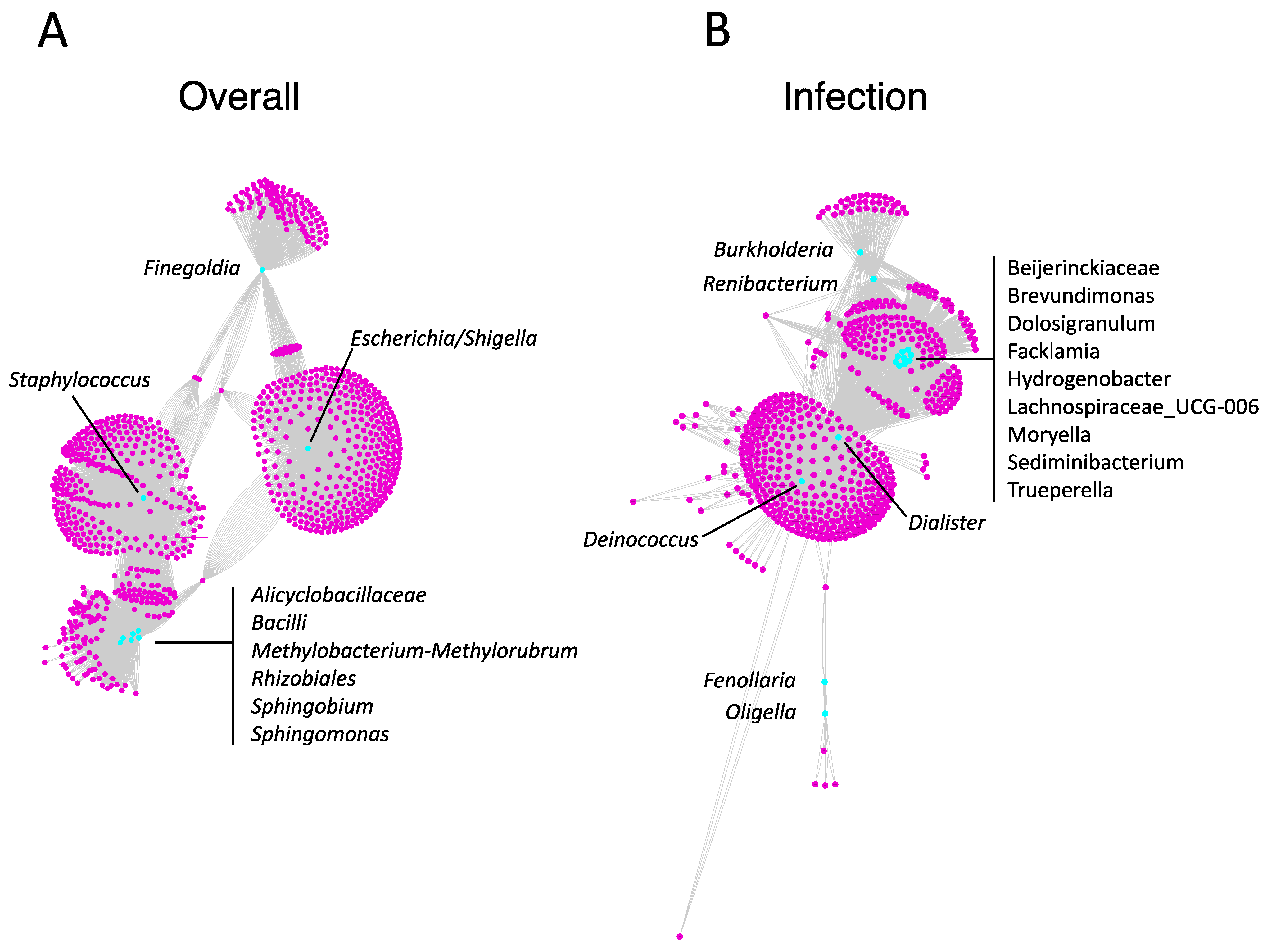
3.2. Biofilm Metabolite Composition and Microbe–Metabolite Interaction Networks
3.3. Reconstitution of Bacterial Biofilm In Vitro
3.4. Biofilm and Antibiotic Resistance Gene Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Cógáin, M.R.; Elliott, D.S. The impact of an antibiotic coating on the artificial urinary sphincter infection rate. J. Urol. 2013, 190, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Kastler, E.C.; Genevois, S.; Gamé, X.; Denys, P.; Richard, F.; Leriche, A.; Saramon, J.P.; Ruffion, A. Treatment of neurogenic male urinary incontinence related to intrinsic sphincter insufficiency with an artificial urinary sphincter: A French retrospective multicentre study. BJU Int. 2011, 107, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.; Poinas, G.; Naoum, K.B.; Bouzoubaa, K.; Wagner, L.; Soustelle, L.; Boukaram, M.; Droupy, S. Long-term results of artificial urinary sphincter for women with type III stress urinary incontinence. Eur. Urol. 2013, 63, 753–758. [Google Scholar] [CrossRef]
- Hüsch, T.; Kretschmer, A.; Thomsen, F.; Kronlachner, D.; Kurosch, M.; Obaje, A.; Anding, R.; Pottek, T.; Rose, A.; Olianas, R. Antibiotic coating of the artificial urinary sphincter (AMS 800): Is it worthwhile? Urology 2017, 103, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Ziegelmann, M.J.; Linder, B.J.; Avant, R.A.; Elliott, D.S. Bacterial cultures at the time of artificial urinary sphincter revision surgery in clinically uninfected devices: A contemporary series. J. Urol. 2019, 201, 1152–1157. [Google Scholar] [CrossRef]
- Elliott, D.S.; Barrett, D.M. Mayo Clinic long-term analysis of the functional durability of the AMS 800 artificial urinary sphincter: A review of 323 cases. J. Urol. 1998, 159, 1206–1208. [Google Scholar] [CrossRef]
- Malaeb, B.S.; Elliott, S.P.; Lee, J.; Anderson, D.W.; Timm, G.W. Novel artificial urinary sphincter in the canine model: The tape mechanical occlusive device. Urology 2011, 77, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Larson, T.; Jhaveri, H.; Yeung, L.L. Adjustable continence therapy (ProACT) for the treatment of male stress urinary incontinence: A systematic review and meta-analysis. Neurourol. Urodyn. 2019, 38, 2051–2059. [Google Scholar] [CrossRef]
- Vilar, F.D.O.; Araújo, L.A.; Lima, S.V. Periurethral constrictor in pediatric urology: Long-term followup. J. Urol. 2004, 171, 2626–2628. [Google Scholar] [CrossRef]
- Schiavini, J.L.; Damião, R.; de Resende Júnior, J.A.D.; Dornas, M.C.; da Costa, D.S.C.L.; Barros, C.B. Treatment of post-prostate surgery urinary incontinence with the periurethral constrictor: A retrospective analysis. Urology 2010, 75, 1488–1492. [Google Scholar] [CrossRef]
- Zampini, A.; Nguyen, A.H.; Rose, E.; Monga, M.; Miller, A.W. Defining dysbiosis in patients with urolithiasis. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kachroo, N.; Lange, D.; Penniston, K.L.; Stern, J.; Tasian, G.; Bajic, P.; Wolfe, A.J.; Suryavanshi, M.; Ticinesi, A.; Meschi, T. Standardization of microbiome studies for urolithiasis: An international consensus agreement. Nat. Rev. Urol. 2021, 18, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Werneburg, G.T.; Adler, A.; Zhang, A.; Mukherjee, S.D.; Haywood, S.; Miller, A.W.; Klein, E.A. Transperineal prostate biopsy is associated with lower tissue core pathogen burden relative to transrectal biopsy: Mechanistic underpinnings for lower infection risk in the transperineal approach. Urology 2022, 165, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Goudarzi, M.; Weber, W.; Mak, T.D.; Chung, J.; Doyle-Eisele, M.; Melo, D.; Brenner, D.J.; Guilmette, R.A.; Fornace, A.J. Development of urinary biomarkers for internal exposure by cesium-137 using a metabolomics approach in mice. Radiat. Res. 2014, 181, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Agudelo, J.; Fedrigon, D.; Faris, A.; Wilkins, L.; Monga, M.; Miller, A.W. Delineating the role of the urinary metabolome in the lithogenesis of calcium-based kidney stones. Urology 2022, 167, 49–55. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Stedtfeld, R.D.; Guo, X.; Stedtfeld, T.M.; Sheng, H.; Williams, M.R.; Hauschild, K.; Gunturu, S.; Tift, L.; Wang, F.; Howe, A. Primer set 2.0 for highly parallel qPCR array targeting antibiotic resistance genes and mobile genetic elements. FEMS Microbiol. 2018, 94, fiy130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, M.T.; Taylor, L.T.; DeLong, E.F. Quantitative analysis of small-subunit rRNA genes in mixed microbial populations via 5′-nuclease assays. Appl. Environ. Microbiol. 2000, 66, 4605–4614. [Google Scholar] [CrossRef] [Green Version]
- Pusz, P.; Bok, E.; Mazurek, J.; Stosik, M.; Baldy-Chudzik, K. Type 1 fimbriae in commensal Escherichia coli derived from healthy humans. Acta Biochim. Pol. 2014, 61, 389–392. [Google Scholar] [CrossRef] [Green Version]
- Nõlvak, H.; Truu, M.; Tiirik, K.; Oopkaup, K.; Sildvee, T.; Kaasik, A.; Mander, Ü.; Truu, J. Dynamics of antibiotic resistance genes and their relationships with system treatment efficiency in a horizontal subsurface flow constructed wetland. Sci. Total Environ. 2013, 461, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.J.; Criado, A.; Palomares, J.C.; Aznar, J. Use of real-time PCR and fluorimetry for rapid detection of rifampin and isoniazid resistance-associated mutations in Mycobacterium tuberculosis. J. Clin. Microbiol. 2000, 38, 3194–3199. [Google Scholar] [CrossRef]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [Green Version]
- Mak, T.D.; Laiakis, E.C.; Goudarzi, M.; Fornace, A.J., Jr. MetaboLyzer: A novel statistical workflow for analyzing postprocessed LC–MS metabolomics data. Anal. Chem. 2014, 86, 506–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2019, 47, D590–D595. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Alvarez-Jarreta, J.; Brasher, C.J.; Nguyen, A.; Hawksworth, J.I.; Rodrigues, P.; Meckelmann, S.; Allen, S.M.; O’Donnell, V.B. LipidFinder on LIPID MAPS: Peak filtering, MS searching and statistical analysis for lipidomics. Bioinformatics 2019, 35, 685–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspi, R.; Altman, T.; Billington, R.; Dreher, K.; Foerster, H.; Fulcher, C.A.; Holland, T.A.; Keseler, I.M.; Kothari, A.; Kubo, A. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of Pathway/Genome Databases. Nucleic Acids Res. 2014, 42, D459–D471. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Hrabek-Smith, J.M.; Carroll, K.K. A comparative study of serum lipoproteins in rabbits fed a natural ingredient diet or low-fat, cholesterol-free, semipurified diets containing casein or isolated soy protein. Biochem. Cell Biol. 1987, 65, 610–616. [Google Scholar] [CrossRef]
- Bodenhofer, U.; Bonatesta, E.; Horejš-Kainrath, C.; Hochreiter, S. msa: An R package for multiple sequence alignment. Bioinformatics 2015, 31, 3997–3999. [Google Scholar] [CrossRef]
- Schliep, K.P. phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: A bioconductor package for handling and analysis of high-throughput phylogenetic sequence data. In Biocomputing 2012; World Scientific: Singapore, 2012; pp. 235–246. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.; Simpson, G.; Solymos, P. Vegan: Community Ecology Package, version 2.5-6; The Comprehensive R Archive Network, 2019.
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, R.G.; Wu, J.R.; Gloor, G.B. Expanding the UniFrac toolbox. PLoS ONE 2016, 11, e0161196. [Google Scholar] [CrossRef] [Green Version]
- Saama, P.M.; Patel, O.V.; Bettegowda, A.; Ireland, J.J.; Smith, G.W. Novel algorithm for transcriptome analysis. Physiol. Genomics. 2006, 28, 62–66. [Google Scholar] [CrossRef]
- Bararpour, N.; Gilardi, F.; Carmeli, C.; Sidibe, J.; Ivanisevic, J.; Caputo, T.; Augsburger, M.; Grabherr, S.; Desvergne, B.; Guex, N. DBnorm as an R package for the comparison and selection of appropriate statistical methods for batch effect correction in metabolomic studies. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.-L.; Ideker, T. Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, T.; Schultz, L.N.; Lauchnor, E.G.; Gerlach, R.; Lange, D. Evaluation of biofilm induced urinary infection stone formation in a novel laboratory model system. J. Urol. 2018, 199, 178–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magera, J.S.; Elliott, D.S. Artificial urinary sphincter infection: Causative organisms in a contemporary series. J. Urol. 2008, 180, 2475–2478. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.K.; Gross, M.S. Biofilm and penile prosthesis infections in the era of coated implants: 2021 update. Int. J. Impot. Res. 2022, 34, 411–415. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef]
- Wilson, S.K.; Costerton, J.W. Biofilm and penile prosthesis infections in the era of coated implants: A review. J. Sex. Med. 2012, 9, 44–53. [Google Scholar] [CrossRef]
- Khouri, R.K.; Ortiz, N.M.; Dropkin, B.M.; Joice, G.A.; Baumgarten, A.S.; Morey, A.F.; Hudak, S.J. Artificial Urinary Sphincter Complications: Risk Factors, Workup, and Clinical Approach. Curr. Urol. Rep. 2021, 22, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Savage, V.J.; Chopra, I.; O’Neill, A.J. Staphylococcus aureus biofilms promote horizontal transfer of antibiotic resistance. Antimicrob. Agents Chemother. 2013, 57, 1968–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darouiche, R.O.; Wall, M.J., Jr.; Itani, K.M.; Otterson, M.F.; Webb, A.L.; Carrick, M.M.; Miller, H.J.; Awad, S.S.; Crosby, C.T.; Mosier, M.C. Chlorhexidine–alcohol versus povidone–iodine for surgical-site antisepsis. N. Engl. J. Med. 2010, 362, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, L.L.; Grewal, S.; Bullock, A.; Lai, H.H.; Brandes, S.B. A comparison of chlorhexidine-alcohol versus povidone-iodine for eliminating skin flora before genitourinary prosthetic surgery: A randomized controlled trial. J. Urol. 2013, 189, 136–140. [Google Scholar] [CrossRef]
- Kavoussi, N.L.; Viers, B.R.; Pagilara, T.J.; Siegel, J.A.; Hofer, M.D.; Cordon, B.; Shakir, N.; Scott, J.; Morey, A.F. Are urine cultures necessary prior to urologic prosthetic surgery? Sex Med. Rev. 2018, 6, 157–161. [Google Scholar] [CrossRef]
- Kavoussi, N.L.; Siegel, J.A.; Viers, B.R.; Pagliara, T.J.; Hofer, M.D.; Cordon, B.H.; Shakir, N.; Scott, J.M.; Morey, A.F. Preoperative urine culture results correlate poorly with bacteriology of urologic prosthetic device infections. J. Sex Med. 2017, 14, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Tosh, P.K. A clinician’s primer on the role of the microbiome in human health and disease. Mayo Clin. Proc. 2014, 89, 107–114. [Google Scholar] [CrossRef]
- Tate, R., 3rd; Alexander, M. Microbial formation and degradation of dimethylamine. Appl. Environ. Microbiol. 1976, 31, 399–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fothergill, J.; Guest, J. Catabolism of L-lysine by Pseudomonas aeruginosa. J. Gen. Microbiol. 1977, 99, 139–155. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Zhang, H.; Lv, M.; Hu, M.; Li, Z.; Gao, C.; Xu, P.; Ma, C. Enzymatic production of 5-aminovalerate from L-lysine using L-lysine monooxygenase and 5-aminovaleramide amidohydrolase. Sci. Rep. 2014, 4, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Carson, C.C. Efficacy of antibiotic impregnation of inflatable penile prostheses in decreasing infection in original implants. J. Urol. 2004, 171, 1611–1614. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Mean (SD) or n (%) (n = 18) |
|---|---|
| Age (years) | 73 (10.2) |
| Sex (male) | 18 (100) |
| Race | |
| –White | 15 (83.3) |
| –Black | 2 (11.1) |
| –Hispanic | 1 (5.6) |
| Body mass index (kg/m2) | 28.9 (5.1) |
| Diabetes mellitus | 2 (11.1) |
| Cardiac disease | 6 (33.3) |
| Current smoker | 0 (0) |
| Device indwelling time (years) | 6.4 (4.8) |
| Operative time at placement (minutes) | 115 (47.1) 1 |
| Indication for device removal | |
| –Device malfunction | 13 (72.2) |
| –Device-associated infection | 1 (5.6) |
| –Cuff erosion | 3 (16.7) |
| –Clinically ineffective | 1 (5.6) |
| Antibiotics during 30 days prior to device removal | 6 (33.3) |
| Strain 1 | Number of Isolates |
|---|---|
| Staphylococcus epidermidis | 2 |
| Staphylococcus lugdunensis | 2 |
| Staphylococcus hyicus | 1 |
| Staphylococcus warneri | 1 |
| Micrococcus luteus | 1 |
| Micrococcus yunnanensis | 1 |
| Enterococcus faecalis | 1 |
| Brevibacterium sp | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Werneburg, G.T.; Hettel, D.; Adler, A.; Mukherjee, S.D.; Lundy, S.D.; Angermeier, K.W.; Wood, H.M.; Gill, B.C.; Vasavada, S.P.; Goldman, H.B.; et al. Biofilms on Indwelling Artificial Urinary Sphincter Devices Harbor Complex Microbe–Metabolite Interaction Networks and Reconstitute Differentially In Vitro by Material Type. Biomedicines 2023, 11, 215. https://doi.org/10.3390/biomedicines11010215
Werneburg GT, Hettel D, Adler A, Mukherjee SD, Lundy SD, Angermeier KW, Wood HM, Gill BC, Vasavada SP, Goldman HB, et al. Biofilms on Indwelling Artificial Urinary Sphincter Devices Harbor Complex Microbe–Metabolite Interaction Networks and Reconstitute Differentially In Vitro by Material Type. Biomedicines. 2023; 11(1):215. https://doi.org/10.3390/biomedicines11010215
Chicago/Turabian StyleWerneburg, Glenn T., Daniel Hettel, Ava Adler, Sromona D. Mukherjee, Scott D. Lundy, Kenneth W. Angermeier, Hadley M. Wood, Bradley C. Gill, Sandip P. Vasavada, Howard B. Goldman, and et al. 2023. "Biofilms on Indwelling Artificial Urinary Sphincter Devices Harbor Complex Microbe–Metabolite Interaction Networks and Reconstitute Differentially In Vitro by Material Type" Biomedicines 11, no. 1: 215. https://doi.org/10.3390/biomedicines11010215