
The Many Faces of Immune Activation in HIV-1 Infection: A Multifactorial Interconnection

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Role of Immune Activation in Progression to AIDS
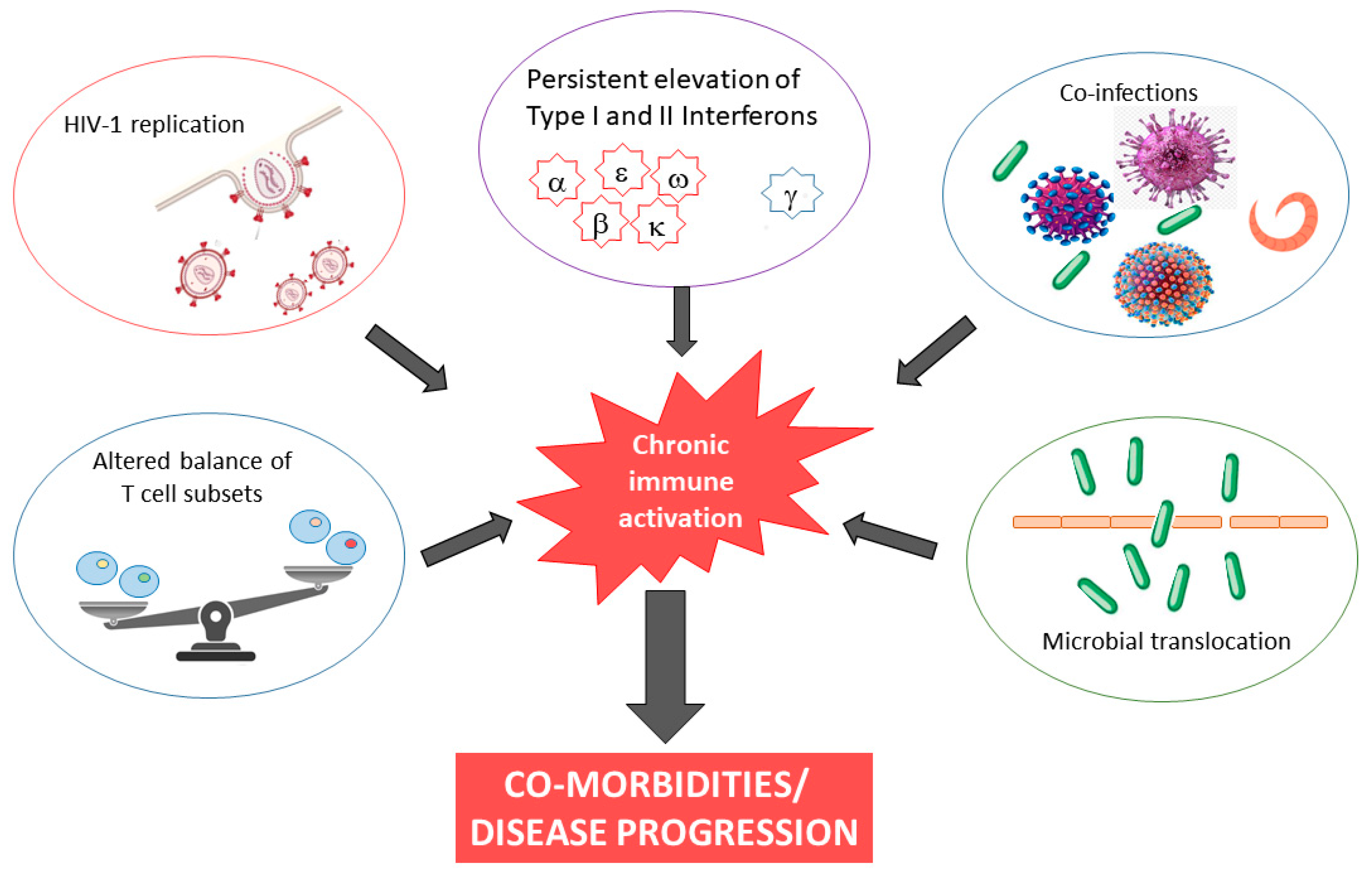
3. Proposed Mechanisms Inducing Chronic Immune Activation
4. The Detrimental Consequences of Systemic Immune Activation
5. Effect of cART on HIV-1 Associated Immune Activation
6. Gender Differences in HIV-1 Associated Immune Activation
7. Experimental and Clinical Approaches to Decrease Chronic Inflammation and Immune Activation
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Douek, D.C. Immune activation, HIV persistence, and the cure. Top. Antivir. Med. 2013, 21, 128–132. [Google Scholar]
- Sokoya, T.; Steel, H.C.; Nieuwoudt, M.; Rossouw, T.M. HIV as a Cause of Immune Activation and Immunosenescence. Mediat. Inflamm. 2017, 2017, 6825493. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, F.; May, M.; Phillips, A. Life expectancy living with HIV: Recent estimates and future implications. Curr. Opin. Infect. Dis. 2013, 26, 17–25. [Google Scholar] [CrossRef]
- Nasi, M.; Pinti, M.; Mussini, C.; Cossarizza, A. Persistent inflammation in HIV infection: Established concepts, new perspectives. Immunol. Lett. 2014, 161, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Nasi, M.; De Biasi, S.; Gibellini, L.; Bianchini, E.; Pecorini, S.; Bacca, V.; Guaraldi, G.; Mussini, C.; Pinti, M.; Cossarizza, A. Ageing and inflammation in patients with HIV infection. Clin. Exp. Immunol. 2017, 187, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Strategies for Management of Antiretroviral Therapy (SMART) Study Group. CD4+ count-guided interruption of antiretroviral treatment. N. Engl. J. Med. 2006, 355, 2283–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatano, H. Immune activation and HIV persistence: Considerations for novel therapeutic interventions. Curr. Opin. HIV AIDS 2013, 8, 211–216. [Google Scholar] [CrossRef] [Green Version]
- d’Ettorre, G.; Paiardini, M.; Ceccarelli, G.; Silvestri, G.; Vullo, V. HIV-associated immune activation: From bench to bedside. AIDS Res. Hum. Retrovir. 2011, 27, 355–364. [Google Scholar] [CrossRef]
- Klatt, N.R.; Silvestri, G.; Hirsch, V. Nonpathogenic Simian Immunodeficiency Virus Infections. Cold Spring Harb. Perspect. Med. 2012, 2, a007153. [Google Scholar] [CrossRef] [Green Version]
- Dunham, R.; Pagliardini, P.; Gordon, S.; Sumpter, B.; Engram, J.; Moanna, A.; Paiardini, M.; Mandl, J.N.; Lawson, B.; Garg, S.; et al. The AIDS resistance of naturally SIV-infected sooty mangabeys is independent of cellular immunity to the virus. Blood 2006, 108, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Silvestri, G. Naturally SIV-infected sooty mangabeys: Are we closer to understanding why they do not develop AIDS? J. Med. Primatol. 2005, 34, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, G.; Paiardini, M.; Pandrea, I.; Lederman, M.M.; Sodora, D.L. Understanding the benign nature of SIV infection in natural hosts. J. Clin. Investig. 2007, 117, 3148–3154. [Google Scholar] [CrossRef]
- Silvestri, G.; Sodora, D.L.; Koup, R.A.; Paiardini, M.; O’Neil, S.P.; McClure, H.M.; Staprans, S.I.; Feinberg, M.B. Nonpathogenic SIV infection of sooty mangabeys is characterized by limited bystander immunopathology despite chronic high-level viremia. Immunity 2003, 18, 441–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paiardini, M.; Cervasi, B.; Sumpter, B.; McClure, H.M.; Sodora, D.L.; Magnani, M.; Staprans, S.I.; Piedimonte, G.; Silvestri, G. Perturbations of cell cycle control in T cells contribute to the different outcomes of simian immunodeficiency virus infection in rhesus macaques and sooty mangabeys. J. Virol. 2006, 80, 634–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumpter, B.; Dunham, R.; Gordon, S.; Engram, J.; Hennessy, M.; Kinter, A.; Paiardini, M.; Cervasi, B.; Klatt, N.; McClure, H.; et al. Correlates of preserved CD4+ T cell homeostasis during natural, nonpathogenic simian immunodeficiency virus infection of sooty mangabeys: Implications for AIDS pathogenesis. J. Immunol. 2007, 178, 1680–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 2006, 12, 1365–1371. [Google Scholar] [CrossRef] [Green Version]
- Pandrea, I.V.; Gautam, R.; Ribeiro, R.M.; Brenchley, J.M.; Butler, I.F.; Pattison, M.; Rasmussen, T.; Marx, P.A.; Silvestri, G.; Lackner, A.A.; et al. Acute loss of intestinal CD4+ T cells is not predictive of simian immunodeficiency virus virulence. J. Immunol. 2007, 179, 3035–3046. [Google Scholar] [CrossRef] [Green Version]
- Pandrea, I.; Gaufin, T.; Brenchley, J.M.; Gautam, R.; Monjure, C.; Gautam, A.; Coleman, C.; Lackner, A.A.; Ribeiro, R.M.; Douek, D.C.; et al. Cutting edge: Experimentally induced immune activation in natural hosts of SIV induces significant increases in viral replication and CD4+ T cell depletion. J. Immunol. 2008, 181, 6687–6691. [Google Scholar] [CrossRef] [Green Version]
- Deeks, S.G.; Walker, B.D. Human immunodeficiency virus controllers: Mechanisms of durable virus control in the absence of antiretroviral therapy. Immunity 2007, 27, 406–416. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, S.; Wilson, E.M.P.; Sheikh, V.; Rupert, A.; Mendoza, D.; Yang, J.; Lempicki, R.; Migueles, S.A.; Sereti, I. Evidence for Innate Immune System Activation in HIV Type 1–Infected Elite Controllers. J. Infect. Dis. 2014, 209, 931–939. [Google Scholar] [CrossRef] [Green Version]
- Okulicz, J.F.; Lambotte, O. Epidemiology and clinical characteristics of elite controllers. Curr. Opin. HIV AIDS 2011, 6, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Gil, E.; Ikediobi, U.; Sutton, R.E. Mechanisms of Virologic Control and Clinical Characteristics of HIV+ Elite/Viremic Controllers. Yale J. Biol. Med. 2017, 90, 245–259. [Google Scholar] [PubMed]
- Genovese, L.; Nebuloni, M.; Alfano, M. Cell-Mediated Immunity in Elite Controllers Naturally Controlling HIV Viral Load. Front. Immunol. 2013, 4, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereyra, F.; Addo, M.M.; Kaufmann, D.E.; Liu, Y.; Miura, T.; Rathod, A.; Baker, B.; Trocha, A.; Rosenberg, R.; Mackey, E.; et al. Genetic and immunologic heterogeneity among persons who control HIV infection in the absence of therapy. J. Infect. Dis. 2008, 197, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Pereyra, F.; Palmer, S.; Miura, T.; Block, B.L.; Wiegand, A.; Rothchild, A.C.; Baker, B.; Rosenberg, R.; Cutrell, E.; Seamanet, M.S.; et al. Persistent low-level viremia in HIV-1 elite controllers and relationship to immunologic parameters. J. Infect. Dis. 2009, 200, 984–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajadi, M.M.; Constantine, N.T.; Mann, D.L.; Charurat, M.; Dadzan, E.; Kadlecik, P.; Redfield, R.R. Epidemiologic characteristics and natural history of HIV-1 natural viral suppressors. J. Acquir. Immune Defic. Syndr. 2009, 50, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Hunt, P.W.; Brenchley, J.; Sinclair, E.; McCune, J.M.; Roland, M.; Page-Shafer, K.; Hsue, P.; Emu, B.; Krone, M.; Lampiris, H.; et al. Relationship between T cell activation and CD4+ T cell count in HIV-seropositive individuals with undetectable plasma HIV RNA levels in the absence of therapy. J. Infect. Dis. 2008, 197, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Sousa, A.E.; Carneiro, J.; Meier-Schellersheim, M.; Grossman, Z.; Victorino, R.M. CD4 T cell depletion is linked directly to immune activation in the pathogenesis of HIV-1 and HIV-2 but only indirectly to the viral load. J. Immunol. 2002, 169, 3400–3406. [Google Scholar] [CrossRef] [Green Version]
- Rajasuriar, R.; Khoury, G.; Kamarulzaman, A.; French, M.A.; Cameron, P.U.; Lewin, S.R. Persistent immune activation in chronic HIV infection: Do any interventions work? AIDS 2013, 27, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Duffy, P.; Wang, X.; Lin, P.H.; Yao, Q.; Chen, C. HIV Nef protein causes endothelial dysfunction in porcine pulmonary arteries and human pulmonary artery endothelial cells. J. Surg. Res. 2009, 156, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Green, L.A.; Gupta, S.K.; Kim, C.; Wang, L.; Almodovar, S.; Flores, S.C.; Prudovsky, I.A.; Jolicoeur, P.; Liu, Z.; et al. Transfer of Intracellular HIV Nef to Endothelium Causes Endothelial Dysfunction. PLoS ONE 2014, 9, e91063. [Google Scholar] [CrossRef] [PubMed]
- Betts, M.R.; Ambrozak, D.R.; Douek, D.C.; Bonhoeffer, S.; Brenchley, J.M.; Casazza, J.P.; Koup, R.A.; Picker, L.J. Analysis of total human immunodeficiency virus (HIV)-specific CD4(+) and CD8(+) T-cell responses: Relationship to viral load in untreated HIV infection. J. Virol. 2001, 75, 11983–11991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, A.; Alter, G.; Frahm, N.; Sidhu, H.; Li, B.; Bagchi, A.; Teigen, N.; Streeck, H.; Stellbrink, H.J.; Hellman, J.; et al. MyD88-dependent immune activation mediated by human immunodeficiency virus type 1-encoded Toll-like receptor ligands. J. Virol. 2007, 81, 8180–8191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appay, V.; Fastenackels, S.; Katlama, C.; Ait-Mohand, H.; Schneider, L.; Guihot, A.; Keller, M.; Grubeck-Loebenstein, B.; Simon, A.; Lambotte, O.; et al. Old age and anti-cytomegalovirus immunity are associated with altered T-cell reconstitution in HIV-1-infected patients. AIDS 2011, 25, 1813–1822. [Google Scholar] [CrossRef] [Green Version]
- Del Cornò, M.; Cappon, A.; Donninelli, G.; Varano, B.; Marra, F.; Gessani, S. HIV-1 gp120 signaling through TLR4 modulates innate immune activation in human macrophages and the biology of hepatic stellate cells. J. Leukoc. Biol. 2016, 100, 599–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douek, D.C.; Brenchley, J.M.; Betts, M.R.; Ambrozak, D.R.; Hill, B.J.; Okamoto, Y.; Casazza, J.P.; Kuruppu, J.; Kunstman, K.; Wolinsky, S.; et al. HIV preferentially infects HIV-specific CD4+ T cells. Nature 2002, 417, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Beignon, A.S.; McKenna, K.; Skoberne, M.; Manches, O.; DaSilva, I.; Kavanagh, D.G.; Larsson, M.; Gorelick, R.J.; Lifson, J.D.; Bhardwaj, N. Endocytosis of HIV- 1 activates plasmacytoid dendritic cells via Toll-like receptor-viral RNA interactions. J. Clin. Invest. 2005, 115, 3265–3275. [Google Scholar] [CrossRef] [Green Version]
- Falasca, F.; Di Carlo, D.; De Vito, C.; Bon, I.; d’Ettorre, G.; Fantauzzi, A.; Mezzaroma, I.; Fimiani, C.; Re, M.C.; Vullo, V.; et al. Evaluation of HIV-DNA and inflammatory markers in HIV-infected individuals with different viral load patterns. BMC Infect. Dis. 2017, 17, 581. [Google Scholar] [CrossRef] [Green Version]
- Olson, A.; Coote, C.; Snyder-Cappione, J.E.; Lin, N.; Sagar, M. HIV-1 Transcription but Not Intact Provirus Levels are Associated With Systemic Inflammation. J. Infect. Dis. 2021, 223, 1934–1942. [Google Scholar] [CrossRef]
- Mexas, A.M.; Graf, E.H.; Pace, M.J.; Yu, J.J.; Papasavvas, E.; Azzoni, L.; Busch, M.P.; Di Mascio, M.; Foulkes, A.S.; Migueles, S.A.; et al. Concurrent measures of total and integrated HIV DNA monitor reservoirs and ongoing replication in eradication trials. AIDS 2012, 26, 2295–2306. [Google Scholar] [CrossRef]
- Allavena, C.; Rodallec, A.; Sécher, S.; Reliquet, V.; Baffoin, S.; André-Garnier, E.; Billaud, E.; Raffi, F.; Ferré, V. Evaluation of residual viremia and quantitation of soluble CD14 in a large cohort of HIV-infected adults on a long-term non-nucleoside reverse transcriptase inhibitor-based regimen. J. Med. Virol. 2013, 85, 1878–1882. [Google Scholar] [CrossRef]
- Wiesmann, F.; Braun, P.; Knickmann, M.; Knechten, H. Low level HIV viremia is more frequent under protease-inhibitor containing first line therapy than under NNRTI-regimens. J. Int. AIDS Soc. 2014, 17, 19828. [Google Scholar] [CrossRef] [PubMed]
- Righetti, E.; Ballon, G.; Ometto, L.; Cattelan, A.M.; Menin, C.; Zanchetta, M.; Chieco-Bianchi, L.; De Rossi, A. Dynamics of Epstein-Barr virus in HIV-1- infected subjects on highly active antiretroviral therapy. AIDS 2002, 16, 63–73. [Google Scholar] [CrossRef]
- Petrara, M.R.; Cattelan, A.M.; Zanchetta, M.; Sasset, L.; Freguja, R.; Gianesin, K.; Cecchetto, M.G.; Carmona, F.; De Rossi, A. Epstein-Barr virus load and immune activation in human immunodeficiency virus type 1-infected patients. J. Clin. Virol. 2012, 53, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, G.; Tincati, C.; Silvestri, G. Microbial Translocation in the Pathogenesis of HIV Infection and AIDS. Clin. Microbiol. Rev. 2013, 26, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Copeland, N.K.; Eller, M.A.; Kim, D.; Creegan, M.; Esber, A.; Eller, L.A.; Semwogerere, M.; Kibuuka, H.; Kiweewa, F.; Crowell, T.A.; et al. Brief Report: Increased Inflammation and Liver Disease in HIV/HBV-Coinfected Individuals. J. Acquir. Immune Defic. Syndr. 2021, 88, 310–313. [Google Scholar] [CrossRef]
- Shmagel, K.V.; Saidakova, E.V.; Shmagel, N.G.; Korolevskaya, L.B.; Chereshnev, V.A.; Robinson, J.; Grivel, J.C.; Douek, D.C.; Margolis, L.; Anthony, D.D.; et al. Systemic inflammation and liver damage in HIV/hepatitis C virus coinfection. HIV Med. 2016, 17, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Liu, X.; Duan, S.; Tang, R.; Zhou, S.; Ye, R.; Yang, Y.; Wang, J.; Yao, S.; He, N. Plasma Inflammatory Biomarkers Associated with Advanced Liver Fibrosis in HIV-HCV-Coinfected Individuals. Int. J. Environ. Res. Public Health 2020, 17, 9474. [Google Scholar] [CrossRef] [PubMed]
- Borkow, G.; Bentwich, Z. Chronic immune activation associated with chronic helminthic and human immunodeficiency virus infections: Role of hyporesponsiveness and anergy. Clin. Microbiol. Rev. 2004, 17, 1012–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenchley, J.M.; Douek, D.C. Microbial Translocation Across the GI Tract. Annu. Rev. Immunol. 2012, 30, 149–173. [Google Scholar] [CrossRef] [Green Version]
- Shan, L.; Siliciano, R.F. Unraveling the relationship between microbial translocation and systemic immune activation in HIV infection. J. Clin. Invest. 2014, 124, 2368–2371. [Google Scholar] [CrossRef] [PubMed]
- Streeck, H.; Nixon, D.F. T cell immunity in acute HIV-1 infection. J. Infect. Dis. 2010, 202 (Suppl. 2), S302–S308. [Google Scholar] [CrossRef] [Green Version]
- Guihot, A.; Dentone, C.; Assoumou, L.; Parizot, C.; Calin, R.; Seang, S.; Soulié, C.; Marcelin, A.G.; Calvez, V.; Autran, B.; et al. Residual immune activation in combined antiretroviral therapy-treated patients with maximally suppressed viremia. AIDS 2016, 30, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Lagares, G.; Romero-Sánchez, M.C.; Ruiz-Mateos, E.; Genebat, M.; Ferrando-Martínez, S.; Muñoz-Fernández, M.Á.; Pacheco, Y.M.; Leal, M. Long- term suppressive combined antiretroviral treatment does not normalize the serum level of soluble CD14. J. Infect. Dis. 2013, 207, 1221–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudd, J.C.; Brenchley, J.M. Gut mucosal barrier dysfunction, microbial dysbiosis, and their role in HIV-1 disease progression. J. Infect. Dis. 2016, 214, S58–S66. [Google Scholar] [CrossRef] [Green Version]
- Müller-Trutwin, M.; Hosmalin, A. Role for plasmacytoid dendritic cells in anti-HIV innate immunity. Immunol. Cell Biol. 2005, 83, 578–583. [Google Scholar] [CrossRef]
- Grossman, Z.; Meier-Schellersheim, M.; Sousa, A.E.; Victorino, R.M.; Paul, W.E. CD4+ T-cell depletion in HIV infection: Are we closer to understanding the cause? Nat. Med. 2002, 8, 319–323. [Google Scholar] [CrossRef]
- Cohen Stuart, J.W.; Hazebergh, M.D.; Hamann, D.; Otto, S.A.; Borleffs, J.C.; Miedema, F.; Boucher, C.A.; de Boer, R.J. The dominant source of CD4+ and CD8+ T-cell activation in HIV infection is antigenic stimulation. J. Acquir. Immune Defic. Syndr. 2000, 25, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Wolf, K.; Tsakiris, D.A.; Weber, R.; Erb, P.; Battegay, M.; Swiss HIV Cohort Study. Antiretroviral therapy reduces markers of endothelial and coagulation activation in patients infected with human immunodeficiency virus type 1. J. Infect. Dis. 2002, 185, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Vandergeeten, C.; Fromentin, R.; Chomont, N. The role of cytokines in the establishment, persistence and eradication of the HIV reservoir. Cytokine Growth Factor Rev. 2012, 23, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Catalfamo, M.; Le Saout, C.; Lane, H.C. The role of cytokines in the pathogenesis and treatment of HIV infection. Cytokine Growth Factor Rev. 2012, 23, 207–214. [Google Scholar] [CrossRef]
- Keating, S.M.; Jacobs, E.S.; Norris, P.J. Soluble mediators of inflammation in HIV and their implications for therapeutics and vaccine development. Cytokine Growth Factor Rev. 2012, 23, 193–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kedzierska, K.; Crowe, S.M. Cytokines and HIV-1: Interactions and clinical implications. Antivir. Chem. Chemother. 2001, 12, 133–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolte, L. Thymic function in HIV-infection. Dan. Med. J. 2013, 60, B4622. [Google Scholar] [PubMed]
- Bandera, A.; Ferrario, G.; Saresella, M.; Marventano, I.; Soria, A.; Zanini, F.; Sabbatini, F.; Airoldi, M.; Marchetti, G.; Franzetti, F.; et al. CD4+ T cell depletion, immune activation and increased production of regulatory T cells in the thymus of HIV-infected individuals. PLoS ONE 2010, 5, e10788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho Tsong Fang, R.; Colantonio, A.D.; Uittenbogaart, C.H. The role of the thymus in HIV infection: A 10 year perspective. AIDS 2008, 22, 171–184. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Chun, T.W.; Davey, R.T., Jr.; Engel, D.; Lane, H.C.; Fauci, A.S. Re-emergence of HIV after stopping therapy. Nature 1999, 401, 874–875. [Google Scholar] [CrossRef]
- Conway, J.M.; Perelson, A.S. Residual Viremia in Treated HIV+ Individuals. PLoS Comput. Biol. 2016, 12, e1004677. [Google Scholar] [CrossRef] [Green Version]
- Luo, R.; Cardozo, E.F.; Piovoso, M.J.; Wu, H.; Buzón, M.J.; Martinez-Picado, J.; Zurakowski, R. Modeling HIV-1 2-LTR dynamics following raltegravir intensification. J. R. Soc. Interface 2013, 10, 20130186. [Google Scholar] [CrossRef]
- Nou, E.; Lo, J.; Grinspoon, S.K. Inflammation, immune activation, and cardiovascular disease in HIV. AIDS 2016, 30, 1495–1509. [Google Scholar] [CrossRef]
- Deeks, S.G.; Phillips, A.N. HIV infection, antiretroviral treatment, ageing, and non-AIDS related morbidity. BMJ 2009, 338, a3172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannetta, M.; Savinelli, S.; Rossi, R.; Mascia, C.; Marocco, R.; Vita, S.; Zuccalà, P.; Zingaropoli, M.A.; Mengoni, F.; Massetti, A.P.; et al. Myeloid and lymphoid activation markers in AIDS and non-AIDS presenters. Immunobiology 2019, 224, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, J.; Jacobs, D.R., Jr.; Baker, J.V.; Calmy, A.; Duprez, D.; La Rosa, A.; Kuller, L.H.; Pett, S.L.; Ristola, M.; Ross, M.J.; et al. Markers of inflammation, coagulation, and renal function are elevated in adults with HIV infection. J. Infect. Dis. 2010, 201, 1788–1795. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.W.; Martin, J.N.; Sinclair, E.; Bredt, B.; Hagos, E.; Lampiris, H.; Deeks, S.G. T cell activation is associated with lower CD4+ T cell gains in human immunodeficiency virus-infected patients with sustained viral suppression during antiretroviral therapy. J. Infect. Dis. 2003, 187, 1534–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdez, H.; Connick, E.; Smith, K.Y.; Lederman, M.M.; Bosch, R.J.; Kim, R.S.; St Clair, M.; Kuritzkes, D.R.; Kessler, H.; Fox, L.; et al. Limited immune restoration after 3 years’ suppression of HIV-1 replication in patients with moderately advanced disease. AIDS 2002, 16, 1859–1866. [Google Scholar] [CrossRef]
- Kuller, L.H.; Tracy, R.; Belloso, W.; De Wit, S.; Drummond, F.; Lane, H.C.; Ledergerber, B.; Lundgren, J.; Neuhaus, J.; Nixon, D.; et al. Inflammatory and coagulation biomarkers and mortality in patients with HIV infection. PLoS Med. 2008, 5, e203. [Google Scholar] [CrossRef]
- Tien, P.C.; Choi, A.I.; Zolopa, A.R.; Benson, C.; Tracy, R.; Scherzer, R.; Bacchetti, P.; Shlipak, M.; Grunfeld, C. Inflammation and mortality in HIV-infected adults: Analysis of the FRAM study cohort. J. Acquir. Immune Defic. Syndr. 2010, 55, 316–322. [Google Scholar] [CrossRef] [Green Version]
- Andrade, B.B.; Hullsiek, K.H.; Boulware, D.R.; Rupert, A.; French, M.A.; Ruxrungtham, K.; Montes, M.L.; Price, H.; Barreiro, P.; Audsley, J.; et al. Biomarkers of inflammation and coagulation are associated with mortality and hepatitis flares in persons coinfected with HIV and hepatitis viruses. J. Infect. Dis. 2013, 207, 1379–1388. [Google Scholar] [CrossRef] [Green Version]
- Sandler, N.G.; Wand, H.; Roque, A.; Law, M.; Nason, M.C.; Nixon, D.E.; Pedersen, C.; Ruxrungtham, K.; Lewin, S.R.; Emery, S.; et al. Plasma levels of soluble CD14 independently predict mortality in HIV infection. J. Infect. Dis. 2011, 203, 780–790. [Google Scholar] [CrossRef] [Green Version]
- Hunt, P.W.; Cao, H.L.; Muzoora, C.; Ssewanyana, I.; Bennett, J.; Emenyonu, N.; Kembabazi, A.; Neilands, T.B.; Bangsberg, D.R.; Deeks, S.G.; et al. Impact of CD8+ T-cell activation on CD4+ T-cell recovery and mortality in HIV- infected Ugandans initiating antiretroviral therapy. AIDS 2011, 25, 2123–2131. [Google Scholar] [CrossRef] [PubMed]
- Duprez, D.A.; Neuhaus, J.; Kuller, L.H.; Tracy, R.; Belloso, W.; De Wit, S.; Drummond, F.; Lane, H.C.; Ledergerber, B.; Lundgren, J.; et al. Inflammation, coagulation and cardiovascular disease in HIV-infected individuals. PLoS ONE 2012, 7, e44454. [Google Scholar] [CrossRef] [PubMed]
- Chun, T.W.; Murray, D.; Justement, J.S.; Hallahan, C.W.; Moir, S.; Kovacs, C.; Fauci, A.S. Relationship between residual plasma viremia and the size of HIV proviral DNA reservoirs in infected individuals receiving effective antiretroviral therapy. J. Infect. Dis. 2011, 204, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Ford, E.S.; Greenwald, J.H.; Richterman, A.G.; Rupert, A.; Dutcher, L.; Badralmaa, Y.; Natarajan, V.; Rehm, C.; Hadigan, C.; Sereti, I. Traditional risk factors and D-dimer predict incident cardiovascular disease events in chronic HIV infection. AIDS 2010, 24, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Borges, Á.H.; Silverberg, M.J.; Wentworth, D.; Grulich, A.E.; Fätkenheuer, G.; Mitsuyasu, R.; Tambussi, G.; Sabin, C.A.; Neaton, J.D.; Lundgren, J.D. Predicting risk of cancer during HIV infection: The role of inflammatory and coagulation biomarkers. AIDS 2013, 27, 1433–1441. [Google Scholar] [CrossRef] [Green Version]
- McComsey, G.A.; Kitch, D.; Sax, P.E.; Tierney, C.; Jahed, N.C.; Melbourne, K.; Ha, B.; Brown, T.T.; Bloom, A.; Fedarko, N.; et al. Associations of inflammatory markers with AIDS and non-AIDS clinical events after initiation of antiretroviral therapy: AIDS clinical trials group A5224s, a substudy of ACTG A5202. J. Acquir. Immune Defic. Syndr. 2014, 65, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Jain, V.; Hartogensis, W.; Bacchetti, P.; Hunt, P.W.; Hatano, H.; Sinclair, E.; Epling, L.; Lee, T.H.; Busch, M.P.; McCune, J.M.; et al. Antiretroviral therapy initiated within 6 months of HIV infection is associated with lower T-cell activation and smaller HIV reservoir size. J. Infect. Dis. 2013, 208, 1202–1211. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, S.; Altfeld, M. Sex differences in HIV-1-mediated immunopathology. Curr. Opin. HIV AIDS 2016, 11, 209–215. [Google Scholar] [CrossRef]
- Ballesteros-Zebadúa, P.; Villarreal, C.; Cocho, G.; Huerta, L.; Estrada, J.L. Differences in HIV-1 viral loads between male and female antiretroviral- untreated Mexican patients. Arch. Med. Res. 2013, 44, 296–301. [Google Scholar] [CrossRef]
- Loupa, C.V.; Rodriguez, B.; McComsey, G.; Gripshover, B.; Salata, R.A.; Valdez, H.; Lisgaris, M.V.; Fulton, S.A.; Lederman, M.M. Gender differences in human immunodeficiency virus (HIV) RNA and CD4 cell counts among new entrants to HIV care. Clin. Microbiol. Infect. 2006, 12, 389–391. [Google Scholar] [CrossRef] [Green Version]
- Collazos, J. Sexual dysfunction in the highly active antiretroviral therapy era. AIDS Rev. 2007, 9, 237–245. [Google Scholar]
- Sáez-Cirión, A.; Bacchus, C.; Hocqueloux, L.; Avettand-Fenoel, V.; Girault, I.; Lecuroux, C.; Potard, V.; Versmisse, P.; Melard, A.; Prazuck, T.; et al. Post-treatment HIV-1 controllers with a long- term virological remission after the interruption of early initiated antiretroviral therapy ANRS VISCONTI Study. PLoS Pathog. 2013, 9, e1003211. [Google Scholar] [CrossRef] [PubMed]
- Fitch, K.V.; Srinivasa, S.; Abbara, S.; Burdo, T.H.; Williams, K.C.; Eneh, P.; Lo, J.; Grinspoon, S.K. Noncalcified coronary atherosclerotic plaque and immune activation in HIV-infected women. J. Infect. Dis. 2013, 208, 1737–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurkovetskiy, L.; Burrows, M.; Khan, A.A.; Graham, L.; Volchkov, P.; Becker, L.; Antonopoulos, D.; Umesaki, Y.; Chervonsky, A.V. Gender bias in autoimmunity is influenced by microbiota. Immunity 2013, 39, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Seillet, C.; Laffont, S.; Trémollières, F.; Rouquié, N.; Ribot, C.; Arnal, J.F.; Douin- Echinard, V.; Gourdy, P.; Guéry, J.C. The TLR-mediated response of plasmacytoid dendritic cells is positively regulated by estradiol in vivo through cell-intrinsic estrogen receptor α signaling. Blood 2012, 119, 454–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libert, C.; Dejager, L.; Pinheiro, I. The X chromosome in immune functions: When a chromosome makes the difference. Nat. Rev. Immunol. 2010, 10, 594–604. [Google Scholar] [CrossRef]
- Carrel, L.; Brown, C.J. When the Lyon(ized chromosome) roars: Ongoing expression from an inactive X chromosome. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160355. [Google Scholar] [CrossRef] [Green Version]
- Mamrut, S.; Avidan, N.; Staun-Ram, E.; Ginzburg, E.; Truffault, F.; Berrih-Aknin, S.; Miller, A. Integrative analysis of methylome and transcriptome in human blood identifies extensive sex- and immune cell-specific differentially methylated regions. Epigenetics 2015, 10, 943–957. [Google Scholar] [CrossRef] [Green Version]
- Butler, K.; Ritter, J.M.; Ellis, S.; Morris, M.R.; Hanson, D.L.; McNicholl, J.M.; Kersh, E.N. A Depot Medroxyprogesterone Acetate Dose That Models Human Use and Its Effect on Vaginal SHIV Acquisition Risk. J. Acquir. Immune Defic. Syndr. 2016, 72, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Prakash, M.; Kapembwa, M.S.; Gotch, F.; Patterson, S. Oral contraceptive use induces upregulation of the CCR5 chemokine receptor on CD4(+) T cells in the cervical epithelium of healthy women. J. Reprod. Immunol. 2002, 54, 117–131. [Google Scholar] [CrossRef]
- Griesbeck, M.; Ziegler, S.; Laffont, S.; Smith, N.; Chauveau, L.; Tomezsko, P.; Sharei, A.; Kourjian, G.; Porichis, F.; Hart, M.; et al. Sex Differences in Plasmacytoid Dendritic Cell Levels of IRF5 Drive Higher IFN-α Production in Women. J. Immunol. 2015, 195, 5327–5336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellors, J.W.; Margolick, J.B.; Phair, J.P.; Rinaldo, C.R.; Detels, R.; Jacobson, L.P.; Muñoz, A. Prognostic value of HIV-1 RNA, CD4 cell count, and CD4 Cell count slope for progression to AIDS and death in untreated HIV-1 infection. JAMA 2007, 297, 2349–2350. [Google Scholar] [CrossRef]
- Wallis, R.S.; Kalayjian, R.; Jacobson, J.M.; Fox, L.; Purdue, L.; Shikuma, C.M.; Arakaki, R.; Snyder, S.; Coombs, R.W.; Bosch, R.J.; et al. A study of the immunology, virology, and safety of prednisone in HIV-1-infected subjects with CD4 cell counts of 200 to 700 mm (−3). J. Acquir. Immune Defic. Syndr. 2003, 32, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Kasang, C.; Kalluvya, S.; Majinge, C.; Kongola, G.; Mlewa, M.; Massawe, I.; Kabyemera, R.; Magambo, K.; Ulmer, A.; Klinker, H.; et al. Effects of prednisolone on disease progression in antiretroviral-untreated HIV infection: A 2-year randomized, double-blind placebo controlled clinical trial. PLoS ONE 2016, 11, e0146678. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, A.; Muller, M.; Bertisch-Mollenhoff, B.; Frietsch, B. Low dose prednisolone reduces CD4+ T cell loss in therapy naive HIV-patients without antiretroviral therapy. Eur. J. Med. Res. 2005, 10, 105–109. [Google Scholar]
- Andrieu, J.M.; Lu, W.; Levy, R. Sustained increases in CD4 cell counts in asymptomatic human. J. Infect. Dis. 1995, 171, 523–530. [Google Scholar] [CrossRef]
- Rizzardi, G.P.; Harari, A.; Capiluppi, B.; Tambussi, G.; Ellefsen, K.; Ciuffreda, D.; Champagne, P.; Bart, P.A.; Chave, J.P.; Lazzarin, A.; et al. Treatment of primary HIV-1 infection with cyclosporin A coupled with highly active antiretroviral therapy. J. Clin. Investig. 2002, 109, 681–688. [Google Scholar] [CrossRef]
- Heredia, A.; Amoroso, A.; Davis, C.L.E.N.; Le, N.; Reardon, E.; Dominique, J.K.; Klingebiel, E.; Gallo, R.C.; Redfield, R.R. Rapamycin causes down-regulation of CCR5 and accumulation of anti-HIV βchemokines: An approach to suppress R5 strains of HIV-1. Proc. Natl. Acad. Sci. USA 2003, 100, 10411–10416. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.; Harris, T.; Childs, G.; Prystowsky, M.B. Rapamycin blocks IL-2-driven T cell cycle progression while preserving T cell survival. Blood Cells Mol. Dis. 2001, 27, 572–585. [Google Scholar] [CrossRef]
- Roy, J.; Paquette, J.S.; Fortin, J.F.; Tremblay, M.J. The immunosuppressant rapamycin represses human immunodeficiency virus type 1 replication. Antimicrob. Agents Chemother. 2002, 46, 3447–3455. [Google Scholar] [CrossRef] [Green Version]
- Di Benedetto, F.; Di Sandro, S.; De Ruvo, N.; Montalti, R.; Ballarin, R.; Guerrini, G.P.; Spaggiari, M.; Guaraldi, G.; Gerunda, G. First report on a series of HIV patients undergoing rapamycin monotherapy after liver transplantation. Transplantation 2010, 89, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Sperber, K.; Kalb, T.H.; Stecher, V.J.; Banerjee, R.; Mayer, L. Inhibition of human immunodeficiency virus type 1 replication by hydroxychloroquine in T cells and monocytes. AIDS Res. Hum. Retrovir. 1993, 9, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Piconi, S.; Parisotto, S.; Rizzardini, G.; Passerini, S.; Terzi, R.; Argenteri, B.; Meraviglia, P.; Capetti, A.; Biasin, M.; Trabattoni, D.; et al. Hydroxychloroquine drastically reduces immune activation in HIV-infected, antiretroviral therapy-treated immunologic nonresponders. Blood 2011, 118, 3263–3272. [Google Scholar] [CrossRef] [PubMed]
- Paton, N.I.; Goodall, R.L.; Dunn, D.T.; Franzen, S.; Collaco-Moraes, Y.; Gazzard, B.G.; Williams, I.G.; Fisher, M.J.; Winston, A.; Fox, J.; et al. Effects of hydroxychloroquine on immune activation and disease progression among HIV-infected patients not receiving antiretroviral Therapy. JAMA 2012, 308, 353–361. [Google Scholar] [CrossRef]
- Routy, J.P.; Angel, J.B.; Patel, M.; Kanagaratham, C.; Radzioch, D.; Kema, I.; Gilmore, N.; Ancuta, P.; Singer, J.; Jenabian, M.A. Assessment of chloroquine as a modulator of immune activation to improve CD4 recovery in immune nonresponding HIV-infected patients receiving antiretroviral therapy. HIV Med. 2015, 16, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, A.; Crum-Cianflone, N.; Higgins, J.; Qin, J.; Rehm, C.; Metcalf, J.; Brandt, C.; Vita, J.; Decker, C.F.; Sklar, P.; et al. High dose atorvastatin decreases cellular markers of immune activation without affecting HIV-1 RNA levels: Results of a double-blind randomized placebo controlled clinical trial. J. Infect. Dis. 2011, 203, 756–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckard, A.R.; Jiang, Y.; Debanne, S.M.; Funderburg, N.T.; McComsey, G.A. Effect of 24 weeks of statin therapy on systemic and vascular inflammation in HIV-infected subjects receiving antiretroviral therapy. J. Infect. Dis. 2014, 209, 1156–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funderburg, N.T.; Jiang, Y.; Debanne, S.M.; Labbato, D.; Juchnowski, S.; Ferrari, B.; Clagett, B.; Robinson, J.; Lederman, M.M.; McComsey, G.A. Rosuvastatin reduces vascular inflammation and T-cell and monocyte activation in HIV-infected subjects on antiretroviral therapy. J. Acquir. Immune Defic. Syndr. 2015, 68, 396–404. [Google Scholar] [CrossRef] [Green Version]
- d’Ettorre, G.; Ceccarelli, G.; Giustini, N.; Serafino, S.; Calantone, N.; De Girolamo, G.; Bianchi, L.; Bellelli, V.; Ascoli-Bartoli, T.; Marcellini, S.; et al. Probiotics reduce inflammation in antiretroviral treated, HIV-infected individuals: Results of the “Probio-HIV” clinical trial. PLoS ONE 2015, 10, e0137200. [Google Scholar] [CrossRef] [Green Version]
- Hummelen, R.; Changalucha, J.; Butamanya, N.L.; Koyama, T.E.; Cook, A.; Habbema, J.D.F.; Reid, G. Effect of 25 weeks probiotic supplementation on immune function of HIV patients. Gut Microbes 2011, 2, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Schunter, M.; Chu, H.; Hayes, T.L.; McConnell, D.; Crawford, S.S.; Luciw, P.A.; Bengmark, S.; Asmuth, D.M.; Brown, J.; Bevins, C.L.; et al. Randomized pilot trial of a synbiotic dietary supplement in chronic HIV-1 infection. BMC Complement. Altern. Med. 2012, 12, 84. [Google Scholar] [CrossRef] [Green Version]
- González-Hernández, L.A.; Jave-Suarez, L.F.; Fafutis-Morris, M.; Montes-Salcedo, K.E.; Valle-Gutierrez, L.G.; Campos-Loza, A.E.; Enciso-Gómez, L.F.; Andrade-Villanueva, J.F. Synbiotic therapy decreases microbial translocation and inflammation and improves immunological status in HIV-infected patients: A double-blind randomized controlled pilot trial. Nutr. J. 2012, 11, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villar-García, J.; Hernández, J.J.; Güerri-Fernández, R.; González, A.; Lerma, E.; Guelar, A.; Saenz, D.; Sorlí, L.; Montero, M.; Horcajada, J.P.; et al. Effect of probiotics (Saccharomyces boulardii) on microbial translocation and inflammation in HIV-treated Patients. J. Acquir. Immune Defic. Syndr. 2015, 68, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Fitch, K.; Abbara, S.; Lee, H.; Stavrou, E.; Sacks, R.; Michel, T.; Hemphill, L.; Torriani, M.; Grinspoon, S. Effects of lifestyle modification and metformin on atherosclerotic indices among HIVinfected patients with the metabolic syndrome. AIDS 2012, 26, 587–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikuma, C.M.; Chew, G.M.; Kohorn, L.; Souza, S.A.; Chow, D.; SahBandar, I.N.; Park, E.Y.; Hanks, N.; Gangcuangco, L.M.A.; Gerschenson, M.; et al. Short communication: Metformin reduces CD4 T cell exhaustion in HIVinfected adults on suppressive antiretroviral therapy. AIDS Res. Hum. Retrovir. 2020, 36, 303–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planas, D.; Pagliuzza, A.; Ponte, R.; Fert, A.; Marchand, L.R.; Massanella, M.; Gosselin, A.; Mehraj, V.; Dupuy, F.P.; Isnard, S.; et al. LILAC pilot study: Effects of metformin on mTOR activation and HIV reservoir persistence during antiretroviral therapy. EBioMedicine 2021, 65, 103270. [Google Scholar] [CrossRef]
- Routy, J.P.; Isnard, S.; Mehraj, V.; Ostrowski, M.; Chomont, N.; Ancuta, P.; Ponte, R.; Planas, D.; Dupuy, F.P.; Angel, J.B. Effect of metformin on the size of the HIV reservoir in non-diabetic ART-treated individuals: Single-arm non-randomised Lilac pilot study protocol. BMJ Open 2019, 9, e028444. [Google Scholar] [CrossRef] [Green Version]
- Best, C.; Struthers, H.; Laciny, E.; Royal, M.; Reeds, D.N.; Yarasheski, K.E. Sitagliptin reduces inflammation and chronic immune cell activation in HIV+ adults with impaired glucose tolerance. J. Clin. Endocrinol. Metab. 2015, 100, 2621–2629. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, M.; Montenont, E.; Hu, L.; Nardi, M.A.; Valdes, V.; Merolla, M.; Gettenberg, G.; Cavanagh, K.; Aberg, J.A.; Bhardwaj, N.; et al. Aspirin attenuates platelet activation and immune activation in HIV-1-infected subjects on antiretroviral therapy. J. Acquir. Immune Defic. Syndr. 2013, 63, 280–288. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, M.P.; Zafar, M.U.; Rodriguez, J.C.; Okoroafor, I.; Heyison, A.; Cavanagh, K.; Rodriguez-Caprio, G.; Weinberg, A.; Escolar, G.; Aberg, J.A.; et al. Targeting thrombogenicity and inflammation in chronic HIV infection. Sci. Adv. 2019, 5, eaav5463. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, F.O.; Torheim, E.A.; Dahm, A.E.; Aaberge, I.S.; Lind, A.; Holm, M.; Aandahl, E.M.; Sandset, P.M.; Taskén, K.; Kvale, D. An exploratory trial of cyclooxygenase type 2 inhibitor in HIV-1 infection: Downregulated immune activation and improved T celldependent vaccine responses. J. Virol. 2011, 85, 6557–6566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prebensen, C.; Trøseid, M.; Ueland, T.; Dahm, A.; Sandset, P.M.; Aaberge, I.; Waalen, K.; Dyrhol-Riise, A.M.; Tasken, K.; Kvale, D. Immune activation and HIV-specific T cell responses are modulated by a cyclooxygenase-2 inhibitor in untreated HIV-infected individuals: An exploratory clinical trial. PLoS ONE 2017, 12, e0176527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuzak, J.A.; Gott, T.M.; Kirkwood, J.S.; Coronado, E.; Hensley-McBain, T.; Miller, C.; Cheu, R.K.; Collier, A.C.; Funderburg, N.T.; Martin, J.N.; et al. Heavy Cannabis use associated with reduction in activated and inflammatory immune cell frequencies in antiretroviral therapy treated human immunodeficiency virus-infected individuals. Clin. Infect. Dis. 2018, 66, 1872–1882. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Peterson, S.N.; Li, Y.; Schrier, R.; Iudicello, J.; Letendre, S.; Morgan, E.; Tang, B.; Grant, I.; Cherner, M. Recent cannabis use in HIV is associated with reduced inflammatory markers in CSF and blood. Neurol.-Neuroimmunol. Neuroinflamm. 2020, 7, e809. [Google Scholar] [CrossRef]
- Valdés-Ferrer, S.I.; Crispin, J.C.; Belaunzarán, P.F.; Cantú-Brito, C.G.; Sierra-Madero, J.; Alcocer-Varela, J. Acetylcholine-esterase inhibitor pyridostigmine decreases T cell overactivation in patients infected by HIV. AIDS Res. Hum. Retrovir. 2009, 25, 749–755. [Google Scholar] [CrossRef]
- Macatangay, B.J.; Jackson, E.K.; Abebe, K.Z.; Comer, D.; Cyktor, J.; Klamar-Blain, C.; Borowski, L.; Gillespie, D.G.; Mellors, J.W.; Rinaldo, C.R.; et al. A randomized, placebo-controlled, pilot clinical trial of dipyridamole to decrease human immunodeficiency virus-associated chronic inflammation. J. Infect. Dis. 2020, 221, 1598–1606. [Google Scholar] [CrossRef]
- Somsouk, M.; Dunham, R.M.; Cohen, M.; Albright, R.; Abdel-Mohsen, M.; Liegler, T.; Lifson, J.; Piatak, M.; Gorelick, R.; Huang, Y.; et al. The immunologic effects of mesalamine in treated HIV-infected individuals with incomplete CD4+ T cell recovery: A randomized crossover trial. PLoS ONE 2014, 9, e116306. [Google Scholar] [CrossRef] [Green Version]
- Read, S.W.; DeGrezia, M.; Ciccone, E.J.; DerSimonian, R.; Higgins, J.; Adelsberger, J.W.; Starling, J.M.; Rehm, C.; Sereti, I. The effect of leflunomide on cycling and activation of T-cells in HIV-1-infected participants. PLoS ONE 2010, 5, e11937. [Google Scholar] [CrossRef]
- Available online: https//:www.eacsociety.org/media/guidelines-11.1_final_09-10.pdf (accessed on 29 October 2022).


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazzuti, L.; Turriziani, O.; Mezzaroma, I. The Many Faces of Immune Activation in HIV-1 Infection: A Multifactorial Interconnection. Biomedicines 2023, 11, 159. https://doi.org/10.3390/biomedicines11010159
Mazzuti L, Turriziani O, Mezzaroma I. The Many Faces of Immune Activation in HIV-1 Infection: A Multifactorial Interconnection. Biomedicines. 2023; 11(1):159. https://doi.org/10.3390/biomedicines11010159
Chicago/Turabian StyleMazzuti, Laura, Ombretta Turriziani, and Ivano Mezzaroma. 2023. "The Many Faces of Immune Activation in HIV-1 Infection: A Multifactorial Interconnection" Biomedicines 11, no. 1: 159. https://doi.org/10.3390/biomedicines11010159