
Patient-Derived Organoids: The Beginning of a New Era in Ovarian Cancer Disease Modeling and Drug Sensitivity Testing
 , ,
, ,
Abstract
:1. Introduction
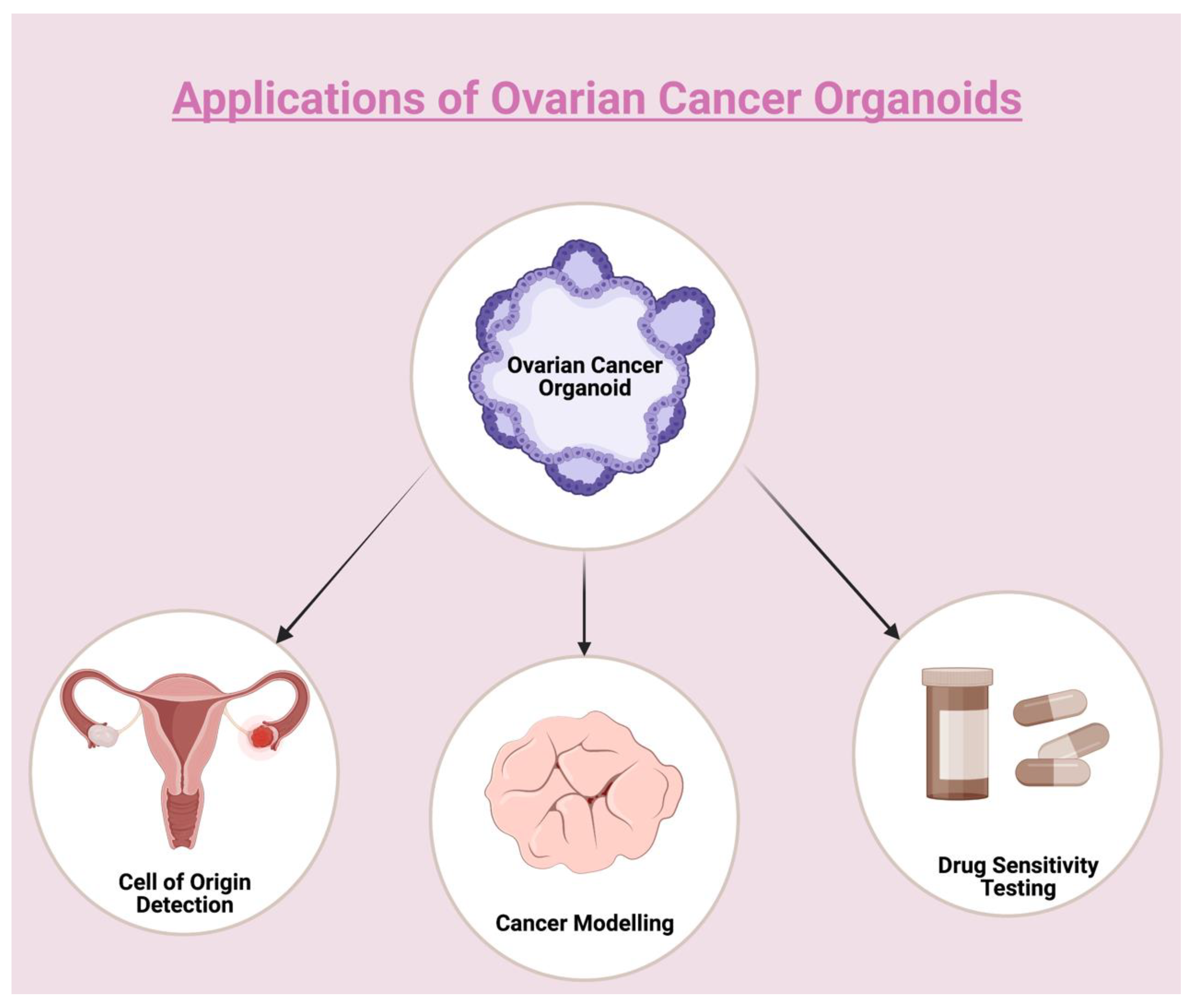
2. The Role of Organoids in the Assessment of HGSOC Origins
3. Efficient Use of Organoids as a Preclinical Model for OC
4. Organoid Culture of OC for Disease Modeling and Drug Sensitivity Testing
5. Advantages and Limitations of OC Organoids-Future Directions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Cancer Society. Key Statistics for Ovarian Cancer; American Cancer Society: Atlanta, GA, USA, 2022. [Google Scholar]
- Prat, J. Ovarian carcinomas: Five distinct diseases with different origins, genetic alterations, and clinicopathological features. Virchows Arch. 2012, 460, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Seebacher, V.; Reinthaller, A.; Koelbl, H.; Concin, N.; Nehoda, R.; Polterauer, S. The Impact of the Duration of Adjuvant Chemo-therapy on Survival in Patients with Epithelial Ovarian Cancer—A Retrospective Study. PLoS ONE 2017, 12, e0169272. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Survival Rates for Ovarian Cancer; American Cancer Society: Atlanta, GA, USA, 2022. [Google Scholar]
- American Cancer Society. Tests for Ovarian Cancer; American Cancer Society: Atlanta, GA, USA, 2020. [Google Scholar]
- American Cancer Society. Can Ovarian Cancer Be Found Early? American Cancer Society: Atlanta, GA, USA, 2020. [Google Scholar]
- American Cancer Society. Treatment of Invasive Epithelial Ovarian Cancers, by Stage; American Cancer Society: Atlanta, GA, USA, 2020. [Google Scholar]
- Qin, T.; Fan, J.; Lu, F.; Zhang, L.; Liu, C.; Xiong, Q.; Zhao, Y.; Chen, G.; Sun, C. Harnessing preclinical models for the interrogation of ovarian cancer. J. Exp. Clin. Cancer Res. 2022, 41, 277. [Google Scholar] [CrossRef] [PubMed]
- Ciucci, A.; Buttarelli, M.; Fagotti, A.; Scambia, G.; Gallo, D. Preclinical models of epithelial ovarian cancer: Practical considerations and challenges for a meaningful application. Cell. Mol. Life Sci. 2022, 79, 364. [Google Scholar] [CrossRef] [PubMed]
- Barnes, B.M.; Nelson, L.; Tighe, A.; Burghel, G.J.; Lin, I.; Desai, S.; McGrail, J.C.; Morgan, R.D.; Taylor, S.S. Distinct transcriptional programs stratify ovarian cancer cell lines into the five major histological subtypes. Genome Med. 2021, 13, 140. [Google Scholar] [CrossRef] [PubMed]
- Beaufort, C.M.; Helmijr, J.C.; Piskorz, A.M.; Hoogstraat, M.; Ruigrok-Ritstier, K.; Besselink, N.; Murtaza, M.; van IJcken, W.F.; Heine, A.A.; Smid, M. Ovarian cancer cell line panel (OCCP): Clinical importance of in vitro morphological subtypes. PLoS ONE 2014, 9, e103988. [Google Scholar] [CrossRef]
- Domcke, S.; Sinha, R.; Levine, D.A.; Sander, C.; Schultz, N. Evaluating cell lines as tumour models by comparison of genomic profiles. Nat. Commun. 2013, 4, 2126. [Google Scholar] [CrossRef] [Green Version]
- Ricci, F.; Bizzaro, F.; Cesca, M.; Guffanti, F.; Ganzinelli, M.; Decio, A.; Ghilardi, C.; Perego, P.; Fruscio, R.; Buda, A. Patient-derived ovarian tumor xenografts recapitulate human clinicopathology and genetic alterations. Cancer Res. 2014, 74, 6980–6990. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, M.; Amant, F.; Biankin, A.V.; Budinská, E.; Byrne, A.T.; Caldas, C.; Clarke, R.B.; de Jong, S.; Jonkers, J.; Mælandsmo, G.M. Patient-derived xenograft models: An emerging platform for translational cancer research. Cancer Discov. 2014, 4, 998–1013. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, U.; Ha, G.; Tseng, Y.-Y.; Greenwald, N.F.; Oh, C.; Shih, J.; McFarland, J.M.; Wong, B.; Boehm, J.S.; Beroukhim, R. Patient-derived xenografts undergo mouse-specific tumor evolution. Nat. Genet. 2017, 49, 1567–1575. [Google Scholar] [CrossRef]
- Powley, I.R.; Patel, M.; Miles, G.; Pringle, H.; Howells, L.; Thomas, A.; Kettleborough, C.; Bryans, J.; Hammonds, T.; MacFarlane, M. Patient-derived explants (PDEs) as a powerful preclinical platform for anti-cancer drug and biomarker discovery. Br. J. Cancer 2020, 122, 735–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, V.M. Genetically engineered mouse models for epithelial ovarian cancer: Are we there yet? In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2014; pp. 106–117. [Google Scholar]
- Zhang, S.; Dolgalev, I.; Zhang, T.; Ran, H.; Levine, D.A.; Neel, B.G. Both fallopian tube and ovarian surface epithelium are cells-of-origin for high-grade serous ovarian carcinoma. Nat. Commun. 2019, 10, 5367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Vries, R.G.; Snippert, H.J.; Van De Wetering, M.; Barker, N.; Stange, D.E.; Van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.; Van Es, J.H.; Van Den Brink, S.; Van Houdt, W.J.; Pronk, A.; Van Gorp, J.; Siersema, P.D. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Lohmussaar, K.; Boretto, M.; Clevers, H. Human-Derived Model Systems in Gynecological Cancer Research. Trends Cancer 2020, 6, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Chen, X.; Dowbaj, A.M.; Sljukic, A.; Bratlie, K.; Lin, L.; Fong, E.L.S.; Balachander, G.M.; Chen, Z.; Soragni, A.; et al. Organoids. Nat. Rev. Methods Prim. 2022, 2, 94. [Google Scholar] [CrossRef]
- de Souza, N. Organoids. Nat. Methods 2018, 15, 23. [Google Scholar] [CrossRef]
- Benton, G.; Kleinman, H.K.; George, J.; Arnaoutova, I. Multiple uses of basement membrane-like matrix (BME/Matrigel) in vitro and in vivo with cancer cells. Int. J. Cancer 2011, 128, 1751–1757. [Google Scholar] [CrossRef] [PubMed]
- Chumduri, C.; Turco, M.Y. Organoids of the female reproductive tract. J. Mol. Med. 2021, 99, 531–553. [Google Scholar] [CrossRef]
- Kopper, O.; De Witte, C.J.; Lõhmussaar, K.; Valle-Inclan, J.E.; Hami, N.; Kester, L.; Balgobind, A.V.; Korving, J.; Proost, N.; Begthel, H.; et al. An organoid platform for ovarian cancer captures intra- and interpatient heter-ogeneity. Nat. Med. 2019, 25, 838–849. [Google Scholar] [CrossRef]
- Joo, S.; Oh, S.-H.; Sittadjody, S.; Opara, E.C.; Jackson, J.D.; Lee, S.J.; Yoo, J.J.; Atala, A. The effect of collagen hydrogel on 3D culture of ovarian follicles. Biomed. Mater. 2016, 11, 065009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Huang, S.; Cheng, S.; Jin, Y.; Zhang, N.; Wang, Y. Application of Ovarian Cancer Organoids in Precision Medicine: Key Challenges and Current Opportunities. Front. Cell Dev. Biol. 2021, 9, 2004. [Google Scholar] [CrossRef] [PubMed]
- Es, H.A.; Montazeri, L.; Aref, A.R.; Vosough, M.; Baharvand, H. Personalized Cancer Medicine: An Organoid Approach. Trends Biotechnol. 2018, 36, 358–371. [Google Scholar]
- Hill, S.J.; Decker, B.; Roberts, E.A. Prediction of DNA Repair Inhibitor Response in Short-Term Patient-Derived Ovarian Cancer Organoids. Cancer Discov. 2018, 8, 1404–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, K.; Berger, H.; Kulbe, H.; Thillainadarasan, S.; Mollenkopf, H.; Zemojtel, T.; Taube, E.; Darb-Esfahani, S.; Mangler, M.; Sehouli, J.; et al. Stable expansion of high-grade serous ovarian cancer organoids requires a low-Wnt environment. EMBO J. 2020, 39, e104013. [Google Scholar] [CrossRef]
- Maru, Y.; Tanaka, N.; Itami, M.; Hippo, Y. Efficient use of patient-derived organoids as a preclinical model for gynecologic tumors. Gynecol. Oncol. 2019, 154, 189–198. [Google Scholar] [CrossRef]
- Nanki, Y.; Chiyoda, T.; Hirasawa, A.; Ookubo, A.; Itoh, M.; Ueno, M.; Akahane, T.; Kameyama, K.; Yamagami, W.; Kataoka, F.; et al. Patient-derived ovarian cancer organoids capture the genomic profiles of primary tumours applicable for drug sensitivity and resistance testing. Sci. Rep. 2020, 10, 12581. [Google Scholar] [CrossRef]
- Maenhoudt, N.; Defraye, C.; Boretto, M.; Jan, Z.; Heremans, R.; Boeckx, B.; Hermans, F.; Arijs, I.; Cox, B.; Van Nieuwenhuysen, E.; et al. Developing organoids from ovarian cancer as experimental and preclinical models. Stem Cell Rep. 2020, 14, 717–729. [Google Scholar] [CrossRef]
- Maenhoudt, N.; Vankelecom, H. Protocol for establishing organoids from human ovarian cancer biopsies. STAR Protoc. 2021, 2, 100429. [Google Scholar] [CrossRef]
- Kim, J.; Park, E.Y.; Kim, O.; Schilder, J.M.; Coffey, D.M.; Cho, C.-H.; Bast, R.C., Jr. Cell Origins of High-Grade Serous Ovarian Cancer. Cancers 2018, 10, 433. [Google Scholar] [CrossRef] [Green Version]
- Kessler, M.; Hoffmann, K.; Brinkmann, V.; Thieck, O.; Jackisch, S.; Toelle, B.; Berger, H.; Mollenkopf, H.-J.; Mangler, M.; Sehouli, J.; et al. The Notch and Wnt pathways regulate stemness and differentiation in human fallopian tube organoids. Nat. Commun. 2015, 6, 8989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lõhmussaar, K.; Kopper, O.; Korving, J.; Begthel, H.; Vreuls, C.P.H.; van Es, J.H.; Clevers, H. Assessing the origin of high-grade serous ovarian cancer using CRISPR-modification of mouse organoids. Nat. Commun. 2020, 11, 2660. [Google Scholar] [CrossRef] [PubMed]
- Maru, Y.; Tanaka, N.; Tatsumi, Y.; Nakamura, Y.; Yao, R.; Noda, T.; Itami, M.; Hippo, Y. Probing the tumorigenic potential of genetic interactions reconstituted in murine fallopian tube organoids. J. Pathol. 2021, 255, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Park, E.S.; Xiang, D.; Li, Z. Long-term organoid culture reveals enrichment of organoid-forming epithelial cells in the fimbrial portion of mouse fallopian tube. Stem Cell Res. 2018, 32, 51–60. [Google Scholar] [CrossRef]
- Yucer, N.; Holzapfel, M.; Vogel, T.J.; Lenaeus, L.; Ornelas, L.; Laury, A.; Sareen, D.; Barrett, R.; Karlan, B.Y.; Svendsen, C.N. Directed Differentiation of Human Induced Pluripotent Stem Cells into Fallopian Tube Epithelium. Sci. Rep. 2017, 7, 10741. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.K.Y.; Lau, T.S.; Chung, K.Y.; Tam, C.; Cheung, T.H.; Yim, S.F.; Lee, J.H.S.; Leung, R.W.T.; Qin, J.; Or, Y.Y.Y.; et al. Short-Form Thymic Stromal Lymphopoietin (sfTSLP) Is the Predominant Isoform Ex-pressed by Gynaecologic Cancers and Promotes Tumour Growth. Cancers 2021, 13, 980. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Gotimer, K.; De Souza, C.; Tepper, C.G.; Karnezis, A.N.; Leiserowitz, G.S.; Chien, J.; Smith, L.H. Short-term organoid culture for drug sensitivity testing of high-grade serous carcinoma. Gynecol. Oncol. 2020, 157, 783–792. [Google Scholar] [CrossRef]
- Yucer, N.; Ahdoot, R.; Workman, M.J.; Laperle, A.H.; Recouvreux, M.S.; Kurowski, K.; Naboulsi, D.J.; Liang, V.; Qu, Y.; Plummer, J.T.; et al. Human iPSC-derived fallopian tube organoids with BRCA1 mutation recapitulate early-stage carcinogenesis. Cell Rep. 2021, 37, 110146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Iyer, S.; Ran, H.; Dolgalev, I.; Gu, S.; Wei, W.; Foster, C.J.; Loomis, C.A.; Olvera, N.; Dao, F.; et al. Genetically Defined, Syngeneic Organoid Platform for Developing Combination Therapies for Ovarian Cancer. Cancer Discov. 2021, 11, 362–383. [Google Scholar] [CrossRef]
- Chen, Y.-A.; Lu, C.-Y.; Cheng, W.-F.; Kuo, K.-T.; Yu, C.-W.; Ho, H.-N.; Chen, H.-F.; Pan, S.-H. An experimental model for ovarian cancer: Propagation of ovarian cancer initiating cells and generation of ovarian cancer organoids. BMC Cancer 2022, 22, 967. [Google Scholar] [CrossRef]
- Wang, W.; Jo, H.; Park, S.; Kim, H.; Kim, S.I.; Han, Y.; Lee, J.; Seol, A.; Kim, J.; Lee, M.; et al. Integrated analysis of ascites and plasma extracellular vesicles identifies a miRNA-based diagnostic signature in ovarian cancer. Cancer Lett. 2022, 542, 215735. [Google Scholar] [CrossRef] [PubMed]
- Franzese, E.; Diana, A.; Centonze, S.; Pignata, S.; De Vita, F.; Ciardiello, F.; Orditura, M. PARP Inhibitors in First-Line Therapy of Ovarian Cancer: Are There Any Doubts? Front. Oncol. 2020, 10, 782. [Google Scholar] [CrossRef] [PubMed]
- Pokhriyal, R.; Hariprasad, R.; Kumar, L.; Hariprasad, G. Chemotherapy Resistance in Advanced Ovarian Cancer Patients. Biomark. Cancer 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Newtson, A.; Zhang, Y.; Devor, E.; Samuelson, M.; Thiel, K.; Leslie, K. Successful Patient-Derived Organoid Culture of Gynecologic Cancers for Disease Modeling and Drug Sensitivity Testing. Cancers 2021, 13, 2901. [Google Scholar] [CrossRef] [PubMed]
- Jabs, J.; Zickgraf, F.M.; Park, J.; Wagner, S.; Jiang, X.; Jechow, K.; Kleinheinz, K.; Toprak, U.H.; Schneider, M.A.; Meister, M.; et al. Screening drug effects in patient-derived cancer cells links organoid responses to genome alterations. Mol. Syst. Biol. 2017, 13, 955. [Google Scholar] [CrossRef]
- Phan, N.; Hong, J.J.; Tofig, B.; Mapua, M.; Elashoff, D.; Moatamed, N.A.; Huang, J.; Memarzadeh, S.; Damoiseaux, R.; Soragni, A. A simple high-throughput approach identifies actionable drug sensitivities in patient-derived tumor organoids. Commun. Biol. 2019, 2, 78. [Google Scholar] [CrossRef]
- de Witte, C.J.; Valle-Inclan, J.E.; Hami, N.; Lõhmussaar, K.; Kopper, O.; Vreuls, C.P.H.; Jonges, G.N.; van Diest, P.; Nguyen, L.; Clevers, H.; et al. Patient-Derived Ovarian Cancer Organoids Mimic Clinical Response and Exhibit Heterogeneous Inter- and Intrapatient Drug Responses. Cell Rep. 2020, 31, 107762. [Google Scholar] [CrossRef]
- Cappuccio, S.; Distefano, M.G.; Ghizzoni, V.; Fagotti, A.; Scambia, G. Trametinib response in heavily pretreated high-grade ovarian cancer: One step towards precision medicine. Gynecol. Oncol. Rep. 2020, 32, 100547. [Google Scholar] [CrossRef]
- Chesnokov, M.S.; Khan, I.; Park, Y.; Ezell, J.; Mehta, G.; Yousif, A.; Hong, L.J.; Buckanovich, R.J.; Takahashi, A.; Chefetz, I. The MEK1/2 pathway as a therapeutic target in high-grade serous ovarian carcinoma. Cancers 2021, 13, 1369. [Google Scholar] [CrossRef]
- D’Amora, P.; Silva, I.D.C.; Tewari, K.S.; Bristow, R.E.; Cappuccini, F.; Evans, S.S.; Salzgeber, M.B.; Addis-Bernard, P.J.; Palma, A.M.; Marchioni, D.M.; et al. Platinum resistance in gynecologic malignancies: Response, disease free and overall sur-vival are predicted by biochemical signature: A metabolomic analysis. Gynecol. Oncol. 2021, 163, 162–170. [Google Scholar] [CrossRef]
- Gorski, J.W.; Zhang, Z.; McCorkle, J.R.; DeJohn, J.M.; Wang, C.; Miller, R.W.; Gallion, H.H.; Dietrich, C.S.; Ueland, F.R.; Kolesar, J.M. Utilizing Patient-Derived Epithelial Ovarian Cancer Tumor Organoids to Predict Carboplatin Resistance. Biomedicines 2021, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- McCorkle, J.R.; Gorski, J.W.; Liu, J.; Riggs, M.B.; McDowell, A.B.; Lin, N.; Wang, C.; Ueland, F.R.; Kolesar, J.M. Lapatinib and poziotinib overcome ABCB1-mediated paclitaxel resistance in ovarian cancer. PLoS ONE 2021, 16, e0254205. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cho, U.; Yoo, A.; Jung, C.L.; Kim, B.; Kim, H.; Lee, J.; Jo, H.; Han, Y.; Song, M.H.; et al. Wnt/beta-Catenin Inhibition by CWP232291 as a Novel Therapeutic Strategy in Ovarian Cancer. Front. Oncol. 2022, 12, 852260. [Google Scholar] [CrossRef] [PubMed]
- Pietilä, E.A.; Gonzalez-Molina, J.; Moyano-Galceran, L.; Jamalzadeh, S.; Zhang, K.; Lehtinen, L.; Turunen, S.P.; Martins, T.A.; Gultekin, O.; Lamminen, T.; et al. Co-evolution of matrisome and adaptive adhesion dynamics drives ovarian cancer chemoresistance. Nat. Commun. 2021, 12, 3904. [Google Scholar] [CrossRef]
- Sun, H.; Wang, H.; Wang, X.; Aoki, Y.; Wang, X.; Yang, Y.; Cheng, X.; Wang, Z.; Wang, X. Aurora-A/SOX8/FOXK1 signaling axis promotes chemoresistance via suppression of cell se-nescence and induction of glucose metabolism in ovarian cancer organoids and cells. Theranostics 2020, 10, 6928–6945. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, W.; Zuo, L.; Xu, M.; Wu, Y.; Huang, J.; Zhang, X.; Li, Y.; Wang, J.; Chen, J.; et al. The Fibrillin-1/VEGFR2/STAT2 signaling axis promotes chemoresistance via modulating gly-colysis and angiogenesis in ovarian cancer organoids and cells. Cancer Commun. 2022, 42, 245–265. [Google Scholar] [CrossRef]
- Neal, A.; Lai, T.; Singh, T.; Rahseparian, N.; Grogan, T.; Elashoff, D.; Scott, P.; Pellegrini, M.; Memarzadeh, S. Combining ReACp53 with Carboplatin to Target High-Grade Serous Ovarian Cancers. Cancers 2021, 13, 5908. [Google Scholar] [CrossRef]
- Soragni, A.; Janzen, D.M.; Johnson, L.M.; Lindgren, A.G.; Nguyen, A.T.-Q.; Tiourin, E.; Soriaga, A.B.; Lu, J.; Jiang, L.; Faull, K.F.; et al. A Designed Inhibitor of p53 Aggregation Rescues p53 Tumor Suppression in Ovarian Carcinomas. Cancer Cell 2016, 29, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Singh, T.; Neal, A.; Dibernardo, G.; Raheseparian, N.; Moatamed, N.A.; Memarzadeh, S. Efficacy of birinapant in combination with carboplatin in targeting platinumresistant epithelial ovarian cancers. Int. J. Oncol. 2022, 60, 35. [Google Scholar] [CrossRef]
- Wambecke, A.; Ahmad, M.; Morice, P.M.; Lambert, B.; Weiswald, L.B.; Vernon, M.; Vigneron, N.; Abeilard, E.; Brotin, E.; Figeac, M.; et al. The lncRNA ‘UCA1’ modulates the response to chemotherapy of ovarian cancer through direct binding to miR-27a-5p and control of UBE2N levels. Mol. Oncol. 2021, 15, 3659–3678. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, M.; Sang, X.; Liu, P.; Gao, J.; Jiang, K.; Cheng, H. Phenethyl Isothiocyanate Enhances the Cytotoxic Effects of PARP Inhibitors in High-Grade Serous Ovarian Cancer Cells. Front. Oncol. 2021, 11, 812264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, L.; Chen, S.; Huang, P.; Ma, L.; Ding, H.; Basappa, B.; Zhu, T.; Lobie, P.E.; Pandey, V. Combined inhibition of BADSer99 phosphorylation and PARP ablates models of recurrent ovarian carcinoma. Commun. Med. 2022, 2, 82. [Google Scholar] [CrossRef] [PubMed]
- Tao, M.; Sun, F.; Wang, J.; Wang, Y.; Zhu, H.; Chen, M.; Liu, L.; Liu, L.; Lin, H.; Wu, X. Developing patient-derived organoids to predict PARP inhibitor response and explore resistance overcoming strategies in ovarian cancer. Pharmacol. Res. 2022, 179, 106232. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Thiel, K.W.; Litman, J.M.; Zhang, Y.; Devor, E.J.; Newtson, A.M.; Schnieders, M.J.; Bosquet, J.G.; Leslie, K.K. Characterization of a TP53 Somatic Variant of Unknown Function From an Ovarian Cancer Patient Using Organoid Culture and Computational Modeling. Clin. Obstet. Gynecol. 2020, 63, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Zhang, Y.; Malmrose, P.K.; Losh, H.A.; Newtson, A.M.; Devor, E.J.; Thiel, K.W.; Leslie, K.K. Blocking autophagy overcomes resistance to dual histone deacetylase and proteasome in-hibition in gynecologic cancer. Cell Death Dis. 2022, 13, 59. [Google Scholar] [CrossRef]
- Qian, X.; LaRochelle, W.J.; Ara, G.; Wu, F.; Petersen, K.D.; Thougaard, A.; Sehested, M.; Lichenstein, H.S.; Jeffers, M. Activity of PXD101, a histone deacetylase inhibitor, in preclinical ovarian cancer studies. Mol. Cancer Ther. 2006, 5, 2086–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, K.; Zhang, G.; Zhang, X.; Yang, M.; Wang, Y.; He, M.; Lu, J.; Liu, H. Stromal infiltrating mast cells identify immunoevasive subtype high-grade serous ovarian cancer with poor prognosis and inferior immunotherapeutic response. Oncoimmunology 2021, 10, 1969075. [Google Scholar] [CrossRef]
- Wan, C.; Keany, M.P.; Dong, H.; Al-Alem, L.F.; Pandya, U.M.; Lazo, S.; Boehnke, K.; Lynch, K.N.; Xu, R.; Zarrella, D.T.; et al. Enhanced Efficacy of Simultaneous PD-1 and PD-L1 Immune Checkpoint Blockade in High-Grade Serous Ovarian Cancer. Cancer Res. 2021, 81, 158–173. [Google Scholar] [CrossRef]
- Florent, R.; Weiswald, L.-B.; Lambert, B.; Brotin, E.; Abeilard, E.; Louis, M.-H.; Babin, G.; Poulain, L.; N’Diaye, M. Bim, Puma and Noxa upregulation by Naftopidil sensitizes ovarian cancer to the BH3-mimetic ABT-737 and the MEK inhibitor Trametinib. Cell Death Dis. 2020, 11, 380. [Google Scholar] [CrossRef]
- Liu, F.; Tang, L.; Tao, M.; Cui, C.; He, D.; Li, L.; Liao, Y.; Gao, Y.; He, J.; Sun, F.; et al. Stichoposide C Exerts Anticancer Effects on Ovarian Cancer by Inducing Autophagy via Inhibiting AKT/mTOR Pathway. OncoTargets Ther. 2022, 15, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Huang, Y.; Qin, T.; You, L.; Lu, F.; Hu, D.; Xiao, R.; Qin, X.; Guo, E.; Yang, B.; et al. AZD5153 reverses palbociclib resistance in ovarian cancer by inhibiting cell cycle-related proteins and the MAPK/PI3K-AKT pathway. Cancer Lett. 2022, 528, 31–44. [Google Scholar] [CrossRef] [PubMed]
- McDowell, A., Jr.; Hill, K.S.; McCorkle, J.R.; Gorski, J.; Zhang, Y.; Salahudeen, A.; Ueland, F.; Kolesar, J. Preclinical Evaluation of Artesunate as an Antineoplastic Agent in Ovarian Cancer Treatment. Diagnostics 2021, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Scattolin, T.; Bortolamiol, E.; Visentin, F. Palladium(II)-eta(3) -Allyl Complexes Bearing N-Trifluoromethyl N-Heterocyclic Carbenes: A New Generation of Anticancer Agents that Restrain the Growth of High-Grade Serous Ovarian Cancer Tumor-oids. Chemistry 2020, 26, 11868–11876. [Google Scholar] [CrossRef] [PubMed]
- Shigeta, S.; Lui, G.Y.; Shaw, R.; Moser, R.; Gurley, K.E.; Durenberger, G.; Rosati, R.; Diaz, R.L.; Ince, T.A.; Swisher, E.M.; et al. Targeting BET Proteins BRD2 and BRD3 in Combination with PI3K-AKT Inhibition as a Therapeutic Strategy for Ovarian Clear Cell Carcinoma. Mol. Cancer Ther. 2021, 20, 691–703. [Google Scholar] [CrossRef]
- Vernon, M.; Lambert, B.; Meryet-Figuière, M.; Brotin, E.; Weiswald, L.-B.; Paysant, H.; Vigneron, N.; Wambecke, A.; Abeilard, E.; Giffard, F.; et al. Functional miRNA Screening Identifies Wide-ranging Antitumor Properties of miR-3622b-5p and Reveals a New Therapeutic Combination Strategy in Ovarian Tumor Organoids. Mol. Cancer Ther. 2020, 19, 1506–1519. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Wu, K.-C.; Harnod, T.; Ding, D.-C. The organoid: A research model for ovarian cancer. Tzu-Chi Med. J. 2022, 34, 255. [Google Scholar]
- Liu, X.; Fang, J.; Huang, S.; Wu, X.; Xie, X.; Wang, J.; Liu, F.; Zhang, M.; Peng, Z.; Hu, N. Tumor-on-a-chip: From bioinspired design to biomedical application. Microsyst. Nanoeng. 2021, 7, 50. [Google Scholar] [CrossRef]
- Saha, B.; Mathur, T.; Tronolone, J.J.; Chokshi, M.; Lokhande, G.K.; Selahi, A.; Gaharwar, A.K.; Afshar-Kharghan, V.; Sood, A.K.; Bao, G. Human tumor microenvironment chip evaluates the consequences of platelet extravasation and combinatorial antitumor-antiplatelet therapy in ovarian cancer. Sci. Adv. 2021, 7, eabg5283. [Google Scholar] [CrossRef]
- Drost, J.; Clevers, H. Organoids in cancer research. Nat. Rev. Cancer 2018, 18, 407–418. [Google Scholar] [CrossRef]
- Psilopatis, I.; Kokkali, S.; Palamaris, K.; Digklia, A.; Vrettou, K.; Theocharis, S. Organoids: A New Chapter in Sarcoma Diagnosis and Treatment. Int. J. Mol. Sci. 2022, 23, 11271. [Google Scholar] [CrossRef]


{kind=link}
| Organoids | Methods | Main Results | References |
|---|---|---|---|
| Human-derived organoids | |||
| Three dimensional organoid cultures from normal human fallopian tubes | Immunohistochemistry (IHC), Microarray expression profiling, Real-time PCR (RT-PCR) |
| [37] |
| Fifteen organoid lines from human peritoneal and omental HGSOC | Flow cytometry (FC), Luminescent cell viability assay, IHC, Immunofluorescence (IF) staining, Quantitative reverse transcription–PCR (qRT-PCR), Western blotting (WB), Drug sensitivity array, Next-generation sequencing (NGS) |
| [31] |
| Human fallopian tube epithelium organoid in vitro model | RT-PCR, IHC, IF |
| [41] |
| Mouse-derived organoids | |||
| Murine fallopian-tube-epithelium-derived and ovarian- surface-epithelium-derived organoids | IF, WB, RNA-sequencing (RNA-seq) |
| [18] |
| Organoid-based tumor progression models of HGSOC from murine fallopian tube and ovarian surface epithelium tissues | Organoid growth assay, FC, IHC, Organoid transfection and genotyping, WB, qRT-PCR, In vitro drug screen, In vivo transplantation assays |
| [38] |
| Murine fallopian tube organoids, tumor-derived organoids | Tumorigenicity assay, WB, Histopathological analysis, Transcriptome analysis, Drug sensitivity assay |
| [39] |
| Murine healthy fallopian tube organoids | RNA quantification, RT-PCR, IF |
| [40] |
| Organoids | Methods | Main Results | References |
|---|---|---|---|
| Human-derived organoids | |||
| Cancer organoid formation of A2780 and IGROV-1 human cancer cells with sfTSLP overexpression or empty-vector expression | Tumor Growth Assay |
| [42] |
| Organoid culture from human HGSOCmalignant effusions | Short-term organoid growth assay, RNA-seq |
| [43] |
| Fifty-six organoid lines from 32 patients, representing all main subtypes of OC | Scanning electron microscopy, Genomic analysis, RNA-seq, Methylation analysis |
| [26] |
| Nine human OC-derived organoids | Targeted next- generation sequencing analysis, Cell proliferation assay, Drug sensitivity assay, Tumorigenicity assay |
| [32] |
| Induced-pluripotent-stem-cell-derived, fallopian tube epithelium organoids from healthy women and OC patients with germline pathogenic BRCA1 mutation | WB, RT-PCR, IHC, Transcriptional Analysis |
| [44] |
| Organoids generated from human induced OC initiating cells | In vitro differentiation assays |
| [46] |
| Human ascites-derived OC organoids | Organoid growth assays |
| [47] |
| Mouse-derived organoids | |||
| Murine fallopian-tube- epithelium- derived organoid-based platform | Chemotaxis assays, FC, IF, IHC, WB, RNA-seq, Shallow Whole Genome Sequencing |
| [45] |
| Therapeutic Agent | Organoids | Main Results | References |
|---|---|---|---|
| Human-derived organoids | |||
| Carboplatin | Two neoadjuvant- carboplatin- exposed and four chemo- naïve HGSOC organoid lines from tissue obtained during debulking surgery |
| [57] |
| Cisplatin | Organoids from cisplatin-sensitive and -resistant human OC tissue samples |
| [61] |
| Multiple agents | Patient-derived ovarian and endometrial cancer organoids |
| [50] |
| Paclitaxel | Primary tumor organoid cell lines from seven unique OC patients |
| [58] |
| Palladium (II)-η3-allyl 4c complex bearing N-trifluoromethyl N-heterocyclic carbenes | Patient-derived OC organoids |
| [79] |
| Platinum-based chemotherapy | Patient-derived organoids from 47 patients with adenocarcinoma of the ovary or uterus |
| [56] |
| Platinum-based chemotherapy | Fifty-six organoid lines from thirty-two patients, representing all main subtypes of OC |
| [26] |
| Platinum-based chemotherapy | Seven human HGSOC organoids |
| [60] |
| Platinum drugs, Paclitaxel, Olaparib | Patient-derived OC organoids |
| [33] |
| Therapeutic Agent | Organoids | Main Results | References |
|---|---|---|---|
| Human-derived organoids | |||
| Anti-PD1 therapy | Short-term human HGSOC organoids |
| [73] |
| Anti-PD-1/PD-L1 antibody | Human HGSOC organoid/immune cell co-cultures |
| [74] |
| Apatinib | Patient-derived HGSOC organoids |
| [62] |
| Artesunate | Patient-derived OC organoids |
| [78] |
| AZD5153 | Patient-derived OC organoids |
| [77] |
| BET/PI3K-AKT-mTOR inhibitors | Patient-derived clear cell OC organoids |
| [80] |
| Birinapant | Organoid panel of 7 epithelial OC cell lines and 10 platinum- resistant primary patient OC samples |
| [65] |
| Carboplatin, PARP inhibitor, CHK1 inhibitor, ATR inhibitor | Short-term patient- derived HGSOC organoids |
| [30] |
| CWP232291 | Organoids from cisplatin-sensitive and cisplatin-resistant patients |
| [59] |
| Erlotinib-ABT-737-combination | Patient-derived serous OC organoids |
| [81] |
| Two hundred and forty kinase inhibitors | Four patient-derived tumor organoids established from two ovarian and one peritoneal high-grade serous carcinomas and one carcinosarcoma of the ovary |
| [52] |
| Multiple agents | Organoid culture from HGSOC malignant effusions |
| [43] |
| Multiple agents | Patient-derived serous OC organoids |
| [51] |
| Multiple agents | Thirty-six whole-genome- characterized organoids from twenty-three OC patients |
| [53] |
| Naftopidil | Patient-derived HGSOC organoids |
| [75] |
| NPB, PARPi | Recurrent human epithelial OC organoids |
| [68] |
| PARPi Platinum-based chemotherapy | Primary and metastatic OC organoids including all histological subtypes (PARPi untreated or post-PARPi treatment recurrent OC) |
| [69] |
| PEITC, PARP inhibitor | Patient-derived HGSOC organoids |
| [67] |
| Proteasome/ HDAC inhibitor combination | Ovarian-carcinosarcoma-patient-derived organoid |
| [70] |
| Proteasome/ HDAC inhibitor combination | Patient-derived ovarian and endometrial cancer organoids |
| [71] |
| PXD101 | Primary OC clinical specimens grown in three-dimensional organoid culture |
| [72] |
| ReACp53 | Human OVCAR3 organoids |
| [63] |
| ReACp53 | Human HGSOC organoids |
| [64] |
| Stichoposide C | Patient-derived HGSOC and endometrioid OC organoids |
| [76] |
| UBE2N | Patient-derived HGSOC organoids |
| [66] |
| Mouse-derived organoids | |||
| Gemcitabine, g-MDSCs, anti-PD-L1 antibodies | Murine fallopian-tube-epithelium-derived organoid-based platform |
| [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Psilopatis, I.; Sykaras, A.G.; Mandrakis, G.; Vrettou, K.; Theocharis, S. Patient-Derived Organoids: The Beginning of a New Era in Ovarian Cancer Disease Modeling and Drug Sensitivity Testing. Biomedicines 2023, 11, 1. https://doi.org/10.3390/biomedicines11010001
Psilopatis I, Sykaras AG, Mandrakis G, Vrettou K, Theocharis S. Patient-Derived Organoids: The Beginning of a New Era in Ovarian Cancer Disease Modeling and Drug Sensitivity Testing. Biomedicines. 2023; 11(1):1. https://doi.org/10.3390/biomedicines11010001
Chicago/Turabian StylePsilopatis, Iason, Alexandros G. Sykaras, Georgios Mandrakis, Kleio Vrettou, and Stamatios Theocharis. 2023. "Patient-Derived Organoids: The Beginning of a New Era in Ovarian Cancer Disease Modeling and Drug Sensitivity Testing" Biomedicines 11, no. 1: 1. https://doi.org/10.3390/biomedicines11010001