
An Update on In Vitro Folliculogenesis: A New Technique for Post-Cancer Fertility

Abstract
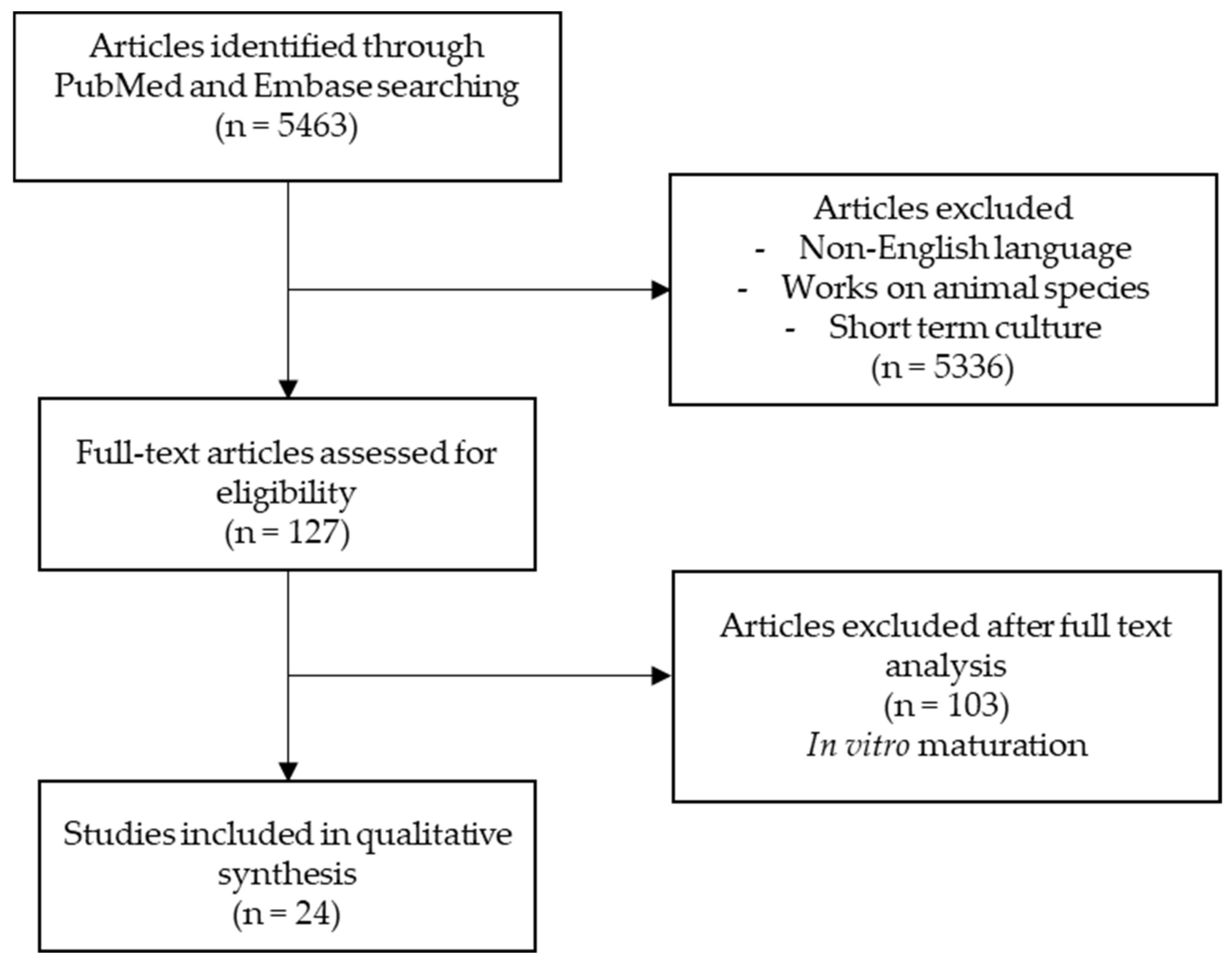
:1. Introduction
2. Methods
3. Description of Culture Media Used for In Vitro Folliculogenesis
4. From the Reserve Follicle to the Secondary Follicle
4.1. Tissue or Isolated Follicle Culture
4.2. Culture Systems
4.3. Supplementation of Culture Systems: Use of Activators
4.3.1. Spontaneous Activation
4.3.2. Activators
4.3.3. Growth Factors
5. From the Secondary Follicle to the Antral Follicle
5.1. Isolation of Follicles
5.2. Culture Systems
5.3. Supplementation of the Culture Medium
6. In Vitro Maturation of Cumulus Oocyte Complexes and Oocyte Quality
7. Discussion
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data availability statement
Acknowledgments
Conflicts of Interest
References
- Takae, S.; Suzuki, N. Current state and future possibilities of ovarian tissue transplantation. Reprod. Med. Biol. 2019, 18, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Dolmans, M.M.; Demylle, D.; Jadoul, P.; Pirard, C.; Squifflet, J.; Martinez-Madrid, B.; van Langendonckt, A. Livebirth after orthotopic transplantation of cryopreserved ovarian tissue. Lancet 2004, 364, 1405–1410. [Google Scholar] [CrossRef]
- Donnez, J.; Dolmans, M.-M. Fertility Preservation in Women. N. Engl. J. Med. 2018, 378, 400–401. [Google Scholar] [CrossRef]
- Marin, L.; Bedoschi, G.; Kawahara, T.; Oktay, K.H. History, Evolution and Current State of Ovarian Tissue Auto-Transplantation with Cryopreserved Tissue: A Successful Translational Research Journey from 1999 to 2020. Reprod. Sci. 2020, 27, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Dolmans, M.-M.; Luyckx, V.; Donnez, J.; Andersen, C.Y.; Greve, T. Risk of transferring malignant cells with transplanted frozen-thawed ovarian tissue. Fertil. Steril. 2013, 99, 1514–1522. [Google Scholar] [CrossRef]
- Masciangelo, R.; Bosisio, C.; Donnez, J.; Amorim, C.A.; Dolmans, M.-M. Safety of ovarian tissue transplantation in patients with borderline ovarian tumors. Hum. Reprod. 2018, 33, 212–219. [Google Scholar] [CrossRef]
- Laronda, M.M.; Duncan, F.E.; Hornick, J.E.; Xu, M.; Pahnke, J.E.; Whelan, K.A.; Shea, L.D.; Woodruff, T.K. Alginate encapsulation supports the growth and differentiation of human primordial follicles within ovarian cortical tissue. J. Assist. Reprod. Genet. 2014, 31, 1013–1028. [Google Scholar] [CrossRef] [PubMed]
- Tagler, D.; Makanji, Y.; Tu, T.; Bernabé, B.P.; Lee, R.; Zhu, J.; Kniazeva, E.; Hornick, J.E.; Woodruff, T.K.; Shea, L.D. Promoting extracellular matrix remodeling via ascorbic acid enhances the survival of primary ovarian follicles encapsulated in alginate hydrogels. Biotechnol. Bioeng. 2014, 111, 1417–1429. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J.; O’Brien, M.; Wigglesworth, K. Mammalian oocyte growth and development in vitro. Mol. Reprod. Dev. 1996, 44, 260–273. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Pendola, J.K.; Eppig, J.J. A revised protocol for in vitro development of mouse oocytes from primordial follicles dramatically improves their developmental competence. Biol. Reprod. 2003, 68, 1682–1686. [Google Scholar] [CrossRef] [Green Version]
- Derrar, N.; Price, C.A.; Sirard, M.A. Effect of growth factors and co-culture with ovarian medulla on the activation of primordial follicles in explants of bovine ovarian cortex. Theriogenology 2000, 54, 587–598. [Google Scholar] [CrossRef]
- Silva, G.M.; Rossetto, R.; Chaves, R.N.; Duarte, A.B.G.; Araújo, V.R.; Feltrin, C.; Bernuci, M.P.; Anselmo-Franci, J.A.; Xu, M.; Woodruff, T.K.; et al. In vitro development of secondary follicles from pre-pubertal and adult goats cultured in two-dimensional or three-dimensional systems. Zygote 2015, 23, 475–484. [Google Scholar] [CrossRef]
- Wandji, S.A.; Srsen, V.; Voss, A.K.; Eppig, J.J.; Fortune, J.E. Initiation in vitro of growth of bovine primordial follicles. Biol. Reprod. 1996, 55, 942–948. [Google Scholar] [CrossRef]
- Wandji, S.A.; Srsen, V.; Nathanielsz, P.W.; Eppig, J.J.; Fortune, J.E. Initiation of growth of baboon primordial follicles in vitro. Hum. Reprod. 1997, 12, 1993–2001. [Google Scholar] [CrossRef]
- Daniel, S.A.; Armstrong, D.T.; Gore-Langton, R.E. Growth and development of rat oocytes in vitro. Gamete Res. 1989, 24, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Carrell, D.T.; Wilcox, A.L. Development of in vitro-matured oocytes from porcine preantral follicles following intracytoplasmic sperm injection. Biol. Reprod. 2001, 65, 1579–1585. [Google Scholar] [CrossRef]
- Gupta, P.S.P.; Ramesh, H.S.; Manjunatha, B.M.; Nandi, S.; Ravindra, J.P. Production of buffalo embryos using oocytes from in vitro grown preantral follicles. Zygote 2008, 16, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Arunakumari, G.; Shanmugasundaram, N.; Rao, V.H. Development of morulae from the oocytes of cultured sheep preantral follicles. Theriogenology 2010, 74, 884–894. [Google Scholar] [CrossRef]
- Magalhães, D.M.; Duarte, A.B.G.; Araújo, V.R.; Brito, I.R.; Soares, T.G.; Lima, I.M.T.; Lopes, C.A.P.; Campello, C.C.; Rodrigues, A.P.R.; Figueiredo, J.R. In vitro production of a caprine embryo from a preantral follicle cultured in media supplemented with growth hormone. Theriogenology 2011, 75, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Peluffo, M.C.; Ting, A.Y.; Zamah, A.M.; Conti, M.; Stouffer, R.L.; Zelinski, M.B.; Hennebold, J.D. Amphiregulin promotes the maturation of oocytes isolated from the small antral follicles of the rhesus macaque. Hum. Reprod. 2012, 27, 2430–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sá, N.A.R.; Ferreira, A.C.A.; Sousa, F.G.C.; Duarte, A.B.G.; Paes, V.M.; Cadenas, J.; Anjos, J.C.; Fernandes, C.C.L.; Rosseto, R.; Cibin, F.W.S.; et al. First pregnancy after in vitro culture of early antral follicles in goats: Positive effects of anethole on follicle development and steroidogenesis. Mol. Reprod. Dev. 2020, 87, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, A. Dynamics of follicular growth in the human: A model from preliminary results. Hum. Reprod. 1986, 1, 81–87. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, J.; Romero, M.M.; Smith, K.N.; Shea, L.D.; Woodruff, T.K. In vitro follicle growth supports human oocyte meiotic maturation. Sci. Rep. 2015, 5, 17323. [Google Scholar] [CrossRef]
- McLaughlin, M.; Albertini, D.F.; Wallace, W.H.B.; Anderson, R.A.; Telfer, E.E. Metaphase II oocytes from human unilaminar follicles grown in a multi-step culture system. Mol. Hum. Reprod. 2018, 24, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Lawson, M.S.; Bean, Y.; Ting, A.Y.; Pejovic, T.; De Geest, K.; Moffitt, M.; Mitalipov, S.M.; Xu, J. Matrix-free 3D culture supports human follicular development from the unilaminar to the antral stage in vitro yielding morphologically normal metaphase II oocytes. Hum. Reprod. 2021, 36, 1326–1338. [Google Scholar] [CrossRef] [PubMed]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef]
- Hovatta, O.; Silye, R.; Abir, R.; Krausz, T.; Winston, R.M. Extracellular matrix improves survival of both stored and fresh human primordial and primary ovarian follicles in long-term culture. Hum. Reprod. 1997, 12, 1032–1036. [Google Scholar] [CrossRef]
- Abir, R.; Franks, S.; Mobberley, M.A.; Moore, P.A.; Margara, R.A.; Winston, R.M. Mechanical isolation and in vitro growth of preantral and small antral human follicles. Fertil. Steril. 1997, 68, 682–688. [Google Scholar] [CrossRef]
- Hreinsson, J.G.; Scott, J.E.; Rasmussen, C.; Swahn, M.L.; Hsueh, A.J.W.; Hovatta, O. Growth differentiation factor-9 promotes the growth, development, and survival of human ovarian follicles in organ culture. J. Clin. Endocrinol. Metab. 2002, 87, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Barrett, S.L.; West-Farrell, E.; Kondapalli, L.A.; Kiesewetter, S.E.; Shea, L.D.; Woodruff, T.K. In vitro grown human ovarian follicles from cancer patients support oocyte growth. Hum. Reprod. 2009, 24, 2531–2540. [Google Scholar] [CrossRef]
- Lerer-Serfaty, G.; Samara, N.; Fisch, B.; Shachar, M.; Kossover, O.; Seliktar, D.; Ben-Haroush, A.; Abir, R. Attempted application of bioengineered/biosynthetic supporting matrices with phosphatidylinositol-trisphosphate-enhancing substances to organ culture of human primordial follicles. J. Assist. Reprod. Genet. 2013, 30, 1279–1288. [Google Scholar] [CrossRef]
- Wang, T.; Yan, L.; Yan, J.; Lu, C.; Xia, X.; Yin, T.; Zhu, X.; Gao, J.; Ding, T.; Hu, W.; et al. Basic fibroblast growth factor promotes the development of human ovarian early follicles during growth in vitro. Hum. Reprod. 2014, 29, 568–576. [Google Scholar] [CrossRef]
- Yin, H.; Kristensen, S.G.; Jiang, H.; Rasmussen, A.; Andersen, C.Y. Survival and growth of isolated pre-antral follicles from human ovarian medulla tissue during long-term 3D culture. Hum. Reprod. 2016, 31, 1531–1539. [Google Scholar] [CrossRef]
- Hosseini, L.; Shirazi, A.; Naderi, M.M.; Shams-Esfandabadi, N.; Borjian Boroujeni, S.; Sarvari, A.; Sadeghnia, S.; Behzadi, B.; Akhondi, M.M. Platelet-rich plasma promotes the development of isolated human primordial and primary follicles to the preantral stage. Reprod. Biomed. Online 2017, 35, 343–350. [Google Scholar] [CrossRef]
- Grosbois, J.; Demeestere, I. Dynamics of PI3K and Hippo signaling pathways during in vitro human follicle activation. Hum. Reprod. 2018, 33, 1705–1714. [Google Scholar] [CrossRef]
- Asadi, E.; Najafi, A.; Moeini, A.; Pirjani, R.; Hassanzadeh, G.; Mikaeili, S.; Salehi, E.; Adutwum, E.; Soleimani, M.; Khosravi, F.; et al. Ovarian tissue culture in the presence of VEGF and fetuin stimulates follicle growth and steroidogenesis. J. Endocrinol. 2017, 232, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, F.; Reid, R.L.; Moini, A.; Abolhassani, F.; Valojerdi, M.R.; Kan, F.W.K. In vitro development of human primordial follicles to preantral stage after vitrification. J. Assist. Reprod. Genet. 2013, 30, 1397–1406. [Google Scholar] [CrossRef] [PubMed]
- Telfer, E.E.; McLaughlin, M.; Ding, C.; Thong, K.J. A two-step serum-free culture system supports development of human oocytes from primordial follicles in the presence of activin. Hum. Reprod. 2008, 23, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, M.; Kinnell, H.L.; Anderson, R.A.; Telfer, E.E. Inhibition of phosphatase and tensin homologue (PTEN) in human ovary in vitro results in increased activation of primordial follicles but compromises development of growing follicles. Mol. Hum. Reprod. 2014, 20, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, R.; Lima-Verde, I.B.; Matos, M.H.T.; Saraiva, M.V.A.; Martins, F.S.; Faustino, L.R.; Araújo, V.R.; Silva, C.M.G.; Name, K.P.O.; Sn, S.N.B.; et al. Interaction between ascorbic acid and follicle-stimulating hormone maintains follicular viability after long-term in vitro culture of caprine preantral follicles. Domest. Anim. Endocrinol. 2009, 37, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Wright, C.S.; Hovatta, O.; Margara, R.; Trew, G.; Winston, R.M.L.; Franks, S.; Hardy, K. Effects of follicle-stimulating hormone and serum substitution on the in-vitro growth of human ovarian follicles. Hum. Reprod. 1999, 14, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, J.; Xu, K.P.; Liu, B.; DiMattina, M. Extracorporeal development and ultrarapid freezing of human fetal ova. J. Assist. Reprod. Genet. 1995, 12, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Hovatta, O.; Wright, C.; Krausz, T.; Hardy, K.; Winston, R.M. Human primordial, primary and secondary ovarian follicles in long-term culture: Effect of partial isolation. Hum. Reprod. 1999, 14, 2519–2524. [Google Scholar] [CrossRef]
- Abir, R.; Roizman, P.; Fisch, B.; Nitke, S.; Okon, E.; Orvieto, R.; Ben Rafael, Z. Pilot study of isolated early human follicles cultured in collagen gels for 24 hours. Hum. Reprod. 1999, 14, 1299–1301. [Google Scholar] [CrossRef]
- Krotz, S.P.; Robins, J.C.; Ferruccio, T.-M.; Moore, R.; Steinhoff, M.M.; Morgan, J.R.; Carson, S. In vitro maturation of oocytes via the pre-fabricated self-assembled artificial human ovary. J. Assist. Reprod. Genet. 2010, 27, 743–750. [Google Scholar] [CrossRef]
- Roy, S.K.; Treacy, B.J. Isolation and long-term culture of human preantral follicles. Fertil. Steril. 1993, 59, 783–790. [Google Scholar] [CrossRef]
- Xia, X.; Wang, T.; Yin, T.; Yan, L.; Yan, J.; Lu, C.; Zhao, L.; Li, M.; Zhang, Y.; Jin, H.; et al. Mesenchymal Stem Cells Facilitate In Vitro Development of Human Preantral Follicle. Reprod. Sci. 2015, 22, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Chiti, M.-C.; Vanacker, J.; Ouni, E.; Tatic, N.; Viswanath, A.; des Rieux, A.; Dolmans, M.-M.; White, L.J.; Amorim, C.A. Ovarian extracellular matrix-based hydrogel for human ovarian follicle survival in vivo: A pilot work. J. Biomed. Mater. Res. B Appl. Biomater. 2022, 110, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kawamura, K.; Cheng, Y.; Liu, S.; Klein, C.; Liu, S.; Duan, E.-K.; Hsueh, A.J.W. Activation of dormant ovarian follicles to generate mature eggs. Proc. Natl. Acad. Sci. USA 2010, 107, 10280–10284. [Google Scholar] [CrossRef]
- Yaseen, M.A.; Wrenzycki, C.; Herrmann, D.; Carnwath, J.W.; Niemann, H. Changes in the relative abundance of mRNA transcripts for insulin-like growth factor (IGF-I and IGF-II) ligands and their receptors (IGF-IR/IGF-IIR) in preimplantation bovine embryos derived from different in vitro systems. Reproduction 2001, 122, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Louhio, H.; Hovatta, O.; Sjöberg, J.; Tuuri, T. The effects of insulin, and insulin-like growth factors I and II on human ovarian follicles in long-term culture. Mol. Hum. Reprod. 2000, 6, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.H.; Leask, R.; Srsen, V.; Riley, S.C.; Spears, N.; Telfer, E.E. Effect of ascorbic acid on health and morphology of bovine preantral follicles during long-term culture. Reproduction 2001, 122, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Gomes, R.G.; Lisboa, L.A.; Silva, C.B.; Max, M.C.; Marino, P.C.; Oliveira, R.L.; González, S.M.; Barreiros, T.R.R.; Marinho, L.S.R.; Seneda, M.M. Improvement of development of equine preantral follicles after 6 days of in vitro culture with ascorbic acid supplementation. Theriogenology 2015, 84, 750–755. [Google Scholar] [CrossRef]
- Lewis, N.; Hinrichs, K.; Leese, H.J.; McGregor Argo, C.; Brison, D.R.; Sturmey, R.G. Glucose concentration during equine in vitro maturation alters mitochondrial function. Reproduction 2020, 160, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Sadeu, J.C.; Smitz, J. Growth differentiation factor-9 and anti-Müllerian hormone expression in cultured human follicles from frozen-thawed ovarian tissue. Reprod. Biomed. Online 2008, 17, 537–548. [Google Scholar] [CrossRef]
- Varelas, X. The Hippo pathway effectors TAZ and YAP in development, homeostasis and disease. Development 2014, 141, 1614–1626. [Google Scholar] [CrossRef]
- Holbourn, K.P.; Acharya, K.R.; Perbal, B. The CCN family of proteins: Structure-function relationships. Trends Biochem. Sci. 2008, 33, 461–473. [Google Scholar] [CrossRef]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef]
- Masciangelo, R.; Hossay, C.; Chiti, M.C.; Manavella, D.D.; Amorim, C.A.; Donnez, J.; Dolmans, M.-M. Role of the PI3K and Hippo pathways in follicle activation after grafting of human ovarian tissue. J. Assist. Reprod. Genet. 2020, 37, 101–108. [Google Scholar] [CrossRef]
- Novella-Maestre, E.; Herraiz, S.; Rodríguez-Iglesias, B.; Díaz-García, C.; Pellicer, A. Short-Term PTEN Inhibition Improves In Vitro Activation of Primordial Follicles, Preserves Follicular Viability, and Restores AMH Levels in Cryopreserved Ovarian Tissue From Cancer Patients. PLoS ONE 2015, 10, e0127786. [Google Scholar] [CrossRef]
- Suzuki, N.; Yoshioka, N.; Takae, S.; Sugishita, Y.; Tamura, M.; Hashimoto, S.; Morimoto, Y.; Kawamura, K. Successful fertility preservation following ovarian tissue vitrification in patients with primary ovarian insufficiency. Hum. Reprod. 2015, 30, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Liu, L.; Adhikari, D.; Jagarlamudi, K.; Rajareddy, S.; Shen, Y.; Du, C.; Tang, W.; Hämäläinen, T.; Peng, S.L.; et al. Oocyte-specific deletion of Pten causes premature activation of the primordial follicle pool. Science 2008, 319, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.-Y.; Liu, Z.; Cahill, N.; Richards, J.S. Targeted disruption of Pten in ovarian granulosa cells enhances ovulation and extends the life span of luteal cells. Mol. Endocrinol. 2008, 22, 2128–2140. [Google Scholar] [CrossRef]
- Caires, K.C.; de Avila, J.M.; Cupp, A.S.; McLean, D.J. VEGFA family isoforms regulate spermatogonial stem cell homeostasis in vivo. Endocrinology 2012, 153, 887–900. [Google Scholar] [CrossRef]
- Abir, R.; Ao, A.; Zhang, X.Y.; Garor, R.; Nitke, S.; Fisch, B. Vascular endothelial growth factor A and its two receptors in human preantral follicles from fetuses, girls, and women. Fertil. Steril. 2010, 93, 2337–2347. [Google Scholar] [CrossRef]
- Yang, M.Y.; Fortune, J.E. Vascular endothelial growth factor stimulates the primary to secondary follicle transition in bovine follicles in vitro. Mol. Reprod. Dev. 2007, 74, 1095–1104. [Google Scholar] [CrossRef]
- Gao, J.-M.; Yan, J.; Li, R.; Li, M.; Yan, L.-Y.; Wang, T.-R.; Zhao, H.-C.; Zhao, Y.; Yu, Y.; Qiao, J. Improvement in the quality of heterotopic allotransplanted mouse ovarian tissues with basic fibroblast growth factor and fibrin hydrogel. Hum. Reprod. 2013, 28, 2784–2793. [Google Scholar] [CrossRef]
- Pors, S.E.; Ramløse, M.; Nikiforov, D.; Lundsgaard, K.; Cheng, J.; Andersen, C.Y.; Kristensen, S.G. Initial steps in reconstruction of the human ovary: Survival of pre-antral stage follicles in a decellularized human ovarian scaffold. Hum. Reprod. 2019, 34, 1523–1535. [Google Scholar] [CrossRef]
- Vanacker, J.; Camboni, A.; Dath, C.; Van Langendonckt, A.; Dolmans, M.-M.; Donnez, J.; Amorim, C.A. Enzymatic isolation of human primordial and primary ovarian follicles with Liberase DH: Protocol for application in a clinical setting. Fertil. Steril. 2011, 96, 379–383.e3. [Google Scholar] [CrossRef]
- Dolmans, M.-M.; Michaux, N.; Camboni, A.; Martinez-Madrid, B.; Van Langendonckt, A.; Nottola, S.A.; Donnez, J. Evaluation of Liberase, a purified enzyme blend, for the isolation of human primordial and primary ovarian follicles. Hum. Reprod. 2006, 21, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Chiti, M.C.; Dolmans, M.-M.; Hobeika, M.; Cernogoraz, A.; Donnez, J.; Amorim, C.A. A modified and tailored human follicle isolation procedure improves follicle recovery and survival. J. Ovarian Res. 2017, 10, 71. [Google Scholar] [CrossRef]
- Shea, L.D.; Woodruff, T.K.; Shikanov, A. Bioengineering the ovarian follicle microenvironment. Annu. Rev. Biomed. Eng. 2014, 16, 29–52. [Google Scholar] [CrossRef]
- Laronda, M.M.; Rutz, A.L.; Xiao, S.; Whelan, K.A.; Duncan, F.E.; Roth, E.W.; Woodruff, T.K.; Shah, R.N. A bioprosthetic ovary created using 3D printed microporous scaffolds restores ovarian function in sterilized mice. Nat. Commun. 2017, 8, 15261. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, V.; Dolmans, M.-M.; Vanacker, J.; Scalercio, S.R.; Donnez, J.; Amorim, C.A. First step in developing a 3D biodegradable fibrin scaffold for an artificial ovary. J. Ovarian Res. 2013, 6, 83. [Google Scholar] [CrossRef]
- Hassani, F.; Ebrahimi, B.; Moini, A.; Ghiaseddin, A.; Bazrafkan, M.; Hassanzadeh, G.H.; Valojerdi, M.R. Chitosan Hydrogel Supports Integrity of Ovarian Follicles during In Vitro Culture: A Preliminary of A Novel Biomaterial for Three Dimensional Culture of Ovarian Follicles. Cell J. 2020, 21, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Hassanpour, A.; Talaei-Khozani, T.; Kargar-Abarghouei, E.; Razban, V.; Vojdani, Z. Decellularized human ovarian scaffold based on a sodium lauryl ester sulfate (SLES)-treated protocol, as a natural three-dimensional scaffold for construction of bioengineered ovaries. Stem Cell Res. Ther. 2018, 9, 252. [Google Scholar] [CrossRef]
- Cadoret, V.; Frapsauce, C.; Jarrier, P.; Maillard, V.; Bonnet, A.; Locatelli, Y.; Royère, D.; Monniaux, D.; Guérif, F.; Monget, P. Molecular evidence that follicle development is accelerated in vitro compared to in vivo. Reproduction 2017, 153, 493–508. [Google Scholar] [CrossRef] [PubMed]
- Wijayarathna, R.; de Kretser, D.M. Activins in reproductive biology and beyond. Hum. Reprod. Update 2016, 22, 342–357. [Google Scholar] [CrossRef]
- Bloise, E.; Ciarmela, P.; Dela Cruz, C.; Luisi, S.; Petraglia, F.; Reis, F.M. Activin A in Mammalian Physiology. Physiol. Rev. 2019, 99, 739–780. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, M.; Bromfield, J.J.; Albertini, D.F.; Telfer, E.E. Activin promotes follicular integrity and oogenesis in cultured pre-antral bovine follicles. Mol. Hum. Reprod. 2010, 16, 644–653. [Google Scholar] [CrossRef] [Green Version]
- Miró, F.; Smyth, C.D.; Hillier, S.G. Development-related effects of recombinant activin on steroid synthesis in rat granulosa cells. Endocrinology 1991, 129, 3388–3394. [Google Scholar] [CrossRef] [PubMed]
- Miró, F.; Hillier, S.G. Relative effects of activin and inhibin on steroid hormone synthesis in primate granulosa cells. J. Clin. Endocrinol. Metab. 1992, 75, 1556–1561. [Google Scholar] [CrossRef]
- Shukovski, L.; Findlay, J.K. Activin-A inhibits oxytocin and progesterone production by preovulatory bovine granulosa cells in vitro. Endocrinology 1990, 126, 2222–2224. [Google Scholar] [CrossRef]
- Kipp, J.L.; Kilen, S.M.; Bristol-Gould, S.; Woodruff, T.K.; Mayo, K.E. Neonatal exposure to estrogens suppresses activin expression and signaling in the mouse ovary. Endocrinology 2007, 148, 1968–1976. [Google Scholar] [CrossRef] [PubMed]
- Hua, V.K.; Fleming, S.D.; Illingworth, P. Effects of protein kinase A and C inhibitors on follicular inhibin and activin during ovulation. Reprod. Biomed. Online 2008, 17, 642–651. [Google Scholar] [CrossRef]
- Seifer, D.B.; Maclaughlin, D.T. Mullerian Inhibiting Substance is an ovarian growth factor of emerging clinical significance. Fertil. Steril. 2007, 88, 539–546. [Google Scholar] [CrossRef]
- Trounson, A.; Wood, C.; Kausche, A. In vitro maturation and the fertilization and developmental competence of oocytes recovered from untreated polycystic ovarian patients. Fertil. Steril. 1994, 62, 353–362. [Google Scholar] [CrossRef]
- De Roo, C.; Lierman, S.; Tilleman, K.; Peynshaert, K.; Braeckmans, K.; Caanen, M.; Lambalk, C.B.; Weyers, S.; T’Sjoen, G.; Cornelissen, R.; et al. Ovarian tissue cryopreservation in female-to-male transgender people: Insights into ovarian histology and physiology after prolonged androgen treatment. Reprod. Biomed. Online 2017, 34, 557–566. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Supplements | Range of Doses | Mean ± SD | Role | Specific notes | References |
|---|---|---|---|---|---|
| FSH (initial stage) | 0.01–1 IU/mL | 0.306 ± 0.360 IU/mL | Decreases follicular atresia [41] | Surprising use at a gonadotropin-independent stage | [7,24,27,31,32,33,42,43,44,48,49] |
| FSH (secondary stage) | 0.01–1.36 IU/mL | 0.556 ± 0.717 IU/mL | Follicular growth and maturation | In the studies showing the longest culture times and using oocytes in metaphase II, the mean ± SD FSH concentration was 0.017 ± 0.009 IU/mL [23,24,25,42]. | [23,28,30,34,39,46,47] |
| LH (secondary stage, antral stage) | 0.03–20 IU/mL | 5.05 ± 9.96 IU/mL | Follicular growth and maturation | Only one team added LH right from the beginning of the in vitro culture at a concentration of 0.3 IU/mL [27] | [24,42,45,49] |
| Insulin | 0.01–5000 µg/mL | 457.39 ± 1506.61 µg/mL | Action on follicular growth, steroidogenesis, and oocyte maturation [50] | Reduced follicular atresia and stimulated in vitro growth [41,51] Should be controlled; at high concentrations, insulin led to decrease in pre-antral follicle vitality, whereas, at low concentrations, it allowed the same expression of oocyte genes as in fresh tissue. In vitro, insulin reduced follicular atresia and stimulated growth [41,51] | [7,23,24,25,29,30,31,32,33,35,36,37,38,39,46,47] |
| Selenium | 4–6.7 ng/mL | 5.04 ± 1.04 ng/mL | Anti-oxidant that reduces the oxidative stress during in vitro culture | [7,23,24,25,29,30,31,32,33,35,36,37,38,46,47] | |
| Glutamine | 2–3 mM | 2.75 ± 0.5 mM | Essential amino-acid | [24,35,36,37,39,44] | |
| Transferrin | 2.5–5500 µg/mL | 1053.42 ± 2214.92 µg/mL | Binds to iron to limit its cellular toxicity | One group used concentrations between 2.5 and 6.25 µg/mL [7,23,24,25,29,30,31,32,35,37,38,39,46], and a second used 1000-fold concentrations, 2.5 to 5.5 mg/mL [33,36,47]. At a concentration in the µg range the results were more encouraging; they showed longer growing times and higher follicle growth rates [7,23,24,25,29,30,35,39]. | [7,23,24,25,29,30,31,32,33,35,36,37,38,39,46,47] |
| Ascorbic acid | 0.05–50 mg/mL | 14.32 ± 24.37 mg/mL | Anti-oxidant that increases follicular vitality (goats [40]; bovine [52]; equine [53]) | Concentrations ranging from 50 µg/mL [7,24,34,35,37,39] to 50 mg/mL [32,47]. | [7,24,32,34,35,37,39,47,48] |
| Glucose | NA | NA | Present in AlphaMEM medium (5 mM) and McCoy’s 5a medium (17 mM) | ||
| Sodium pyruvate | 0.47 mM | 0.47 ± 0.00 mM | [44,47] | ||
| 8-Bromo-cGMP | 2.5 mM | NA | [29] | ||
| Bovine fetuin | 0.005–1 mg/mL | 0.33 ± 0.57 mg/mL | [7,23,33] | ||
| Linoleic acid | 5.35 µg/mL [46] |
| Authors | Year | Tissue Size (mm) | Culture Period (Days) | Matrices | Use of a Culture System | System Description | Relative Change in the Number of Follicles between Beginning and End of Culture (%) | Peak Day of Secondary Follicles | Observations | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | W | T | Primordial Follicles | Primary Follicles | Secondary Follicles | Antral Follicles | ||||||||
| Hovatta et al | 1997 | NA | NA | 0.1–0.3 | 21 | Tissue | 2-D | Millicel insert + extracellular matrix | +1% | −17% | +14% | +1% | D 21 | - |
| Zhang et al. | 1995 | 1 | 1 | 1 | 25 | Tissue | 2-D | Costar membrane insert | NA | NA | +49% | 0% | NA | - |
| Wright et al. | 1999 | NA | NA | 1–3 | 15 | Tissue | 2-D | Matrigel | −36% | +36% (single category) | 0% | D 10 | - | |
| Hovatta et al. | 1999 | NA | NA | 0.1–0.3 | 28 | Tissue | 2-D | Millicel insert + extracellular matrix | −57% | +31% | +25% | 0% | D 7–9 | - |
| Hreinsson et al. | 2002 | 1 | 1 | 1 | 14 | Tissue | 2-D | Millicel insert + extracellular matrix | −45.9% | −9.4% | +55.3% | 0% | D 14 | - |
| Sadeu et al. | 2008 | 1 | 1 | 1 | 28 | Tissue | No | - | −33.2% | +26.3% | +6.8% | 0% | D 28 | Results in a single patient |
| Telfer et al. | 2008 | 0.5 | 0.5 | 0.5 | 10 | Tissue | No | - | −30% | +15% | +15% | 0% | D 6 | Follicle isolation for continued culture |
| Lerer-Serfaty et al. | 2013 | NA | NA | 1–2 | 12 | Tissue | 3-D | Alginate | −41% | +46% (single category) | 0% | D 7 | - | |
| Khosravi et al. | 2013 | 2 | 2 | 0.5 | 7 | Tissue | No | - | −35.4% | +4.8% | +26.2% | 0% | D 7 | - |
| Laronda et al. | 2014 | 1 | 1 | 0.5 | 45 | Tissue | 3-D | Alginate | NA | NA | NA | 1 from 15 ovarian tissues | NA | - |
| McLaughlin et al. | 2014 | 4 | 2 | 1 | 6 | Tissue | No | - | −46.2% | +60% (single category) | 0% | D 6 | - | |
| Asadi et al. | 2017 | 4 | 2 | 1 | 6 | Tissue | No | - | −48.1% | +7.8% | +35.7% | 0% | D 6 | - |
| Grosbois et al. | 2018 | 4 | 2 | 1 | 6 | Tissue | No | - | −52% | +29% | +35% | 0% | D 6 | - |
| McLaughlin et al. | 2018 | 1 | 1 | 0.5 | 8 | Tissue | No | - | −37.4% | +29.5% | +7.9% | 0% | D 8 | Follicle isolation for continued culture |
| Wang et al. | 2014 | - | - | - | 8 | Isolated follicles | 3-D | Alginate | −30.8% | −28.5% | +0.8% | +60% | D 8 | Follicle isolation for continued culture |
| Hosseini et al. | 2017 | - | - | - | 10 | Isolated follicles | 3-D | Alginate | NA | NA | NA | 0% | D 10 | Report on an increase in follicular diameter (52.4 to 176.4 µm), no count per histological category |
| Xu et al. | 2021 | Disc: volume 0.014 mm3–diameter 300 µm | 21 | Tissue | No | - | NA | NA | NA | NA | D 21 | No results on the evolution of the follicles but on AMH secretion | ||
| Authors | Year | Activator | Concentration | Duration of Activation (Days) | Total Culture Duration (Days) | Matrix | Culture System | Relative Change in the Number of Follicles between Beginning and End of Culture (%) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Primordial Follicles | Primary Follicles | Secondary Follicles | Antral Follicles | ||||||||
| Zhang et al. | 1995 | No | - | - | 25 | Tissue | Costar membrane insert | NA | NA | 49% | 0% |
| Hovatta et al. | 1997 | No | 21 | Tissue | Millicel insert + extracellular matrix | +1% | −17% | +14% | +1% | ||
| Wright et al. | 1999 | No | - | - | 15 | Tissue | Matrigel | −36% | 36% (single category) | 0% | |
| Hovatta et al. | 1999 | No | - | - | 28 | Tissue | Millicel insert + extracellular matrix | −57% | +31% | +25% | 0% |
| Hreinsson et al. | 2002 | GDF9 | 200 ng/mL | 14 | 14 | Tissue | Millicel insert + extracellular matrix | −45.9% | -9.4% | +55.3% | 0% |
| Sadeu and Smitz | 2008 | No | - | - | 28 | Tissue | No | −33.2% | +26.3% | +6.8% | 0% |
| Telfer et al. | 2008 | No | - | - | 10 | Tissue | No | −30% | +15% | +15% | 0% |
| Lerer-Serfaty et al. | 2013 | bpV/740YP | - | - | 12 | Tissue | Alginate | −41% | 46% (single category) | 0% | |
| Khosravi et al. | 2013 | No | - | - | 7 | Tissue | No | −35.4% | +4.8% | +26.2% | 0% |
| Laronda et al. | 2014 | No | - | - | 45 | Tissue | Alginate | NA | NA | NA | 1 out of 15 ovarian tissues |
| McLaughlin et al. | 2014 | bpV | 1 µM | 6 | 6 | Tissue | No | −46.2% | 60% (single category) | 0% | |
| Asadi et al. | 2017 | VEGFA165 | 100 ng/mL | 6 | 6 | Tissue | No | −48.1% | +7.8% | +35.7% | 0% |
| Grosbois et al. | 2018 | mTOR | - | 1–2 | 6 | Tissue | No | −52% | +29% | +35% | 0% |
| McLaughlin et al. | 2018 | No | - | - | 8 | Tissue | No | −37.4% | +29.5% | +7.9% | 0% |
| Wang et al. | 2014 | bFGF | 200 ng/mL | 8 | 8 | Isolated follicles | Alginate | −30.8% | −28.5% | +0.8% | +60% |
| Hosseini et al. | 2017 | +/−PRP | - | - | 10 | Isolated follicles | Alginate | NA | NA | NA | 0% |
| Xu et al. | 2021 | No | - | - | 21 | Tissue | No | NA | NA | NA | NA |
| Chiti et al. | 2022 | No | - | - | 7 | Isolated follicles | Alginate + extracellular matrix hydrogel | NA | NA | NA | 0% |
| Authors | Year | Type of Digestion | Mechanical Digestion | Enzymatic Digestion | Outcomes * | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Needle Diameter | Collagenase | DNase | Neutral Red Solution | Liberase | Time (min) | Temperature (°C) | ||||
| Roy et al. | 1993 | Enzymatic | - | 2.4 IU/mL | 180 IU/mL | - | - | 60 | 37 | Tertiary |
| Zhang et al. | 1995 | NM | - | - | - | - | - | - | MII | |
| Abir et al. | 1997 | Mechanical | 21 G | - | - | - | - | - | - | Secondary |
| Xu et al. | 2009 | Enzymatic + Mechanical | 25 G | 0.20% | 0.02% | - | - | 90 | 37 | Tertiary |
| Xiao et al. | 2015 | Mechanical | 25 G | - | - | - | - | - | - | MII |
| Xia et al. | 2015 | NM | - | - | - | - | - | - | - | Secondary |
| Yin et al. | 2016 | Enzymatic | - | 0.2 mg/mL | 0.2 mg/mL | 50 mg/mL | 0.04 mg/mL | NM | NM | Tertiary |
| McLaughlin et al. | 2018 | Mechanical | 25 G | - | - | - | - | - | - | MII |
| Xu et al. | 2021 | Mechanical | NM | MII | ||||||
| Authors | Year | Multistep | Step | Culture System | Biomaterial | Concentration | Duration (Days) | Matrix | Activator | Outcomes | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial Stage | Diameter (µm) | Final Stage | Survival Rate (%) | Diameter (µm) | Start of Antrum | ||||||||||
| Roy et al. | 1993 | No | 2 | 2-D | Agar | 0.60% | 5 | Isolated follicles | No | Secondary | NA | Tertiary | NA | NA | NA |
| Zhang et al. | 1995 | Yes | 2 | 2-D | Costar membrane insert (collagen) | NA | 30–40 | Isolated follicles | No | Secondary | 60 | Tertiary | NA | NA | NA |
| Zhang et al. | 1995 | Yes | 3 | No | - | - | 1.5 | COC | NA | COC | NA | MII | 25% | 80 | - |
| Abir et al. | 1997 | No | 2 | 2-D | Millicell insert | - | 28 | Isolated follicles | No | Secondary | NA | Secondary | NA | 351 ± 270 | NA |
| Abir et al. | 1999 | No | 2 | 3-D | Collagen | NA | 1 | No | Secondary | NA | Secondary | 0% | NA | ||
| Xu et al. | 2009 | No | 2 | 3-D | Matrigel | 33% | 30 | Isolated follicles | No | Secondary | 170.8 ± 51.1 | Tertiary | 75% | 715 ± 68 | D 12 |
| Xu et al. | 2009 | 2 | 3-D | Alginate | 0.50% | 30 | Isolated follicles | No | Secondary | 178.4 ± 69.2 | Tertiary | 75% | 715 ± 68 | D 12 | |
| Krotz et al. | 2010 | No | 3 | 3-D | 3-D granulosa and theca cells | NA | 7 | COC | No | COC | NA | MII | NA | NA | NA |
| Xiao et al. | 2015 | Yes | 2 | 3-D | Alginate | 0.50% | 10–15 | Isolated follicles | No | Secondary | 165.8 ± 32.3 | Tertiary | NA | 500 | NA |
| Xiao et al. | 2015 | Yes | 3 | No | - | - | 25–30 | Isolated follicles | NA | Tertiary | NA | MII * | NA | NA | - |
| Xia et al. | 2015 | No | 2 | 3-D | Alginate + mesenchymal stem cells | 1% | 8 | Isolated follicles | No | Secondary | 54.7 ± 2.7 | Secondary | NA | 82.9 | NA |
| Xia et al. | 2015 | No | 2 | 3-D | Alginate | 1% | 8 | Isolated follicles | No | Secondary | 53 ± 3.6 | Secondary | NA | 69.5 ± 5.5 | NA |
| Yin et al. | 2016 | No | 2 | 3-D | Alginate | 0.30–0.50% | 30 | Isolated follicles | No | Secondary | 184 ± 35 | Tertiary | 60% | 661 ± 120 | D 20 |
| McLaughlin et al. | 2018 | Yes | 2 | No | - | 8 | Isolated follicles | Activin A | Secondary | 100–150 | Tertiary | NA | NA | NA | |
| McLaughlin et al. | 2018 | Yes | 3 | 2-D | Track-etched nucleopore membranes | NA | 4 | COC | Activin A | COC | 100 | MII * | NA | NA | - |
| Xu et al. | 2021 | Yes | 2 | No | - | - | 42 | Isolated follicles | AMH modulation | Secondary | 125.0–198.4 | Tertiary | 50% | >600 | D 21 |
| Xu et al. | 2021 | Yes | 3 | No | - | - | 1.5 | COC | No | COC | NA | MII | NA | >110 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labrune, E.; Salle, B.; Lornage, J. An Update on In Vitro Folliculogenesis: A New Technique for Post-Cancer Fertility. Biomedicines 2022, 10, 2217. https://doi.org/10.3390/biomedicines10092217
Labrune E, Salle B, Lornage J. An Update on In Vitro Folliculogenesis: A New Technique for Post-Cancer Fertility. Biomedicines. 2022; 10(9):2217. https://doi.org/10.3390/biomedicines10092217
Chicago/Turabian StyleLabrune, Elsa, Bruno Salle, and Jacqueline Lornage. 2022. "An Update on In Vitro Folliculogenesis: A New Technique for Post-Cancer Fertility" Biomedicines 10, no. 9: 2217. https://doi.org/10.3390/biomedicines10092217