
Is Dutasteride a Therapeutic Alternative for Amyotrophic Lateral Sclerosis?

 , , , , and
, , , , and 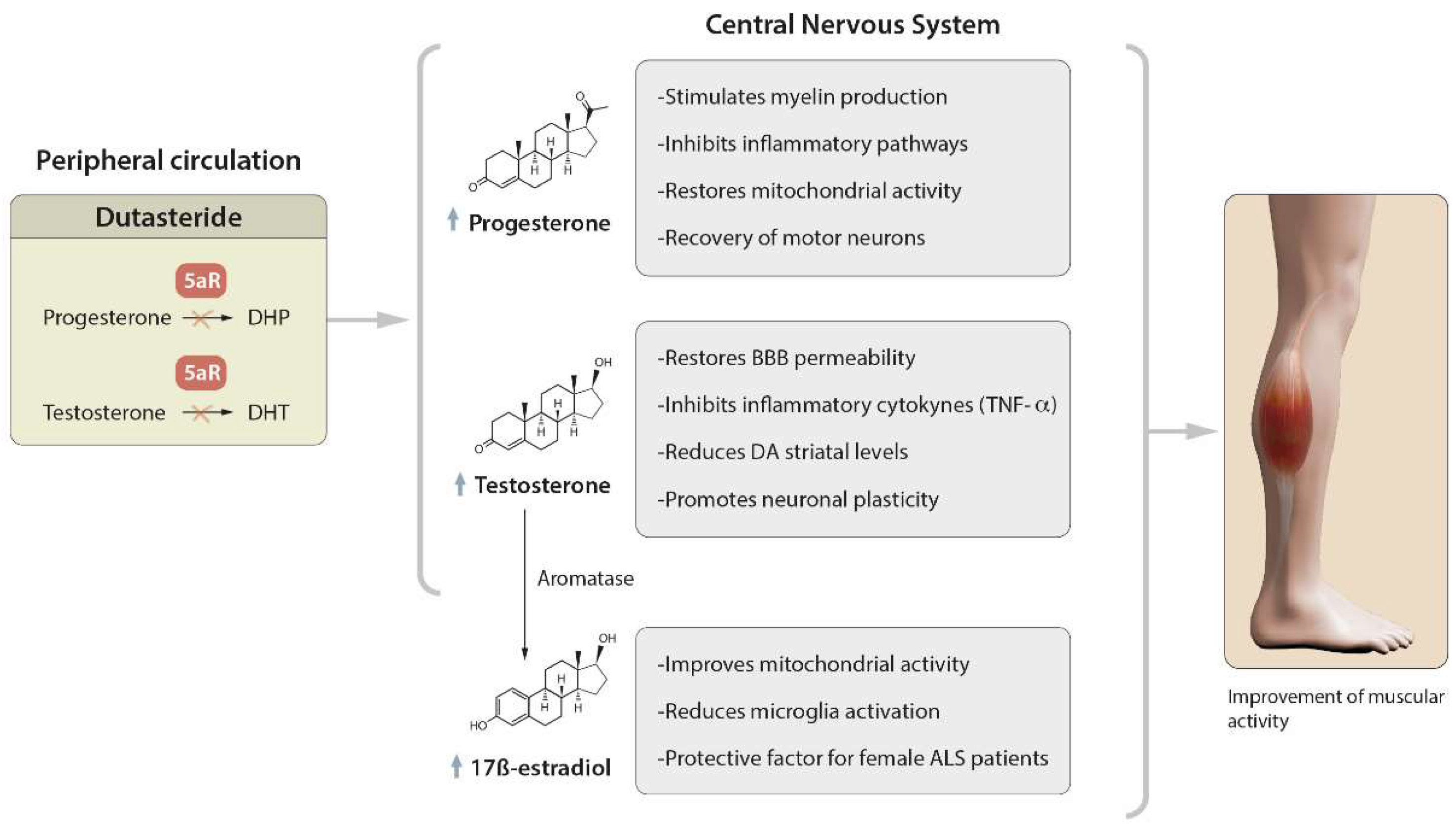{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Pathogenesis of Amyotrophic Lateral Sclerosis
2.1. The Role of Steroid Hormones
2.2. Mitochondrial Damage and Oxidative Stress
2.3. Mutations and Abnormal Protein Aggregation: SOD1, C9ORF72, TDP-43, FUS, VCP
2.4. Neuroinflammation: Activation of Classical NF-κB Pathway
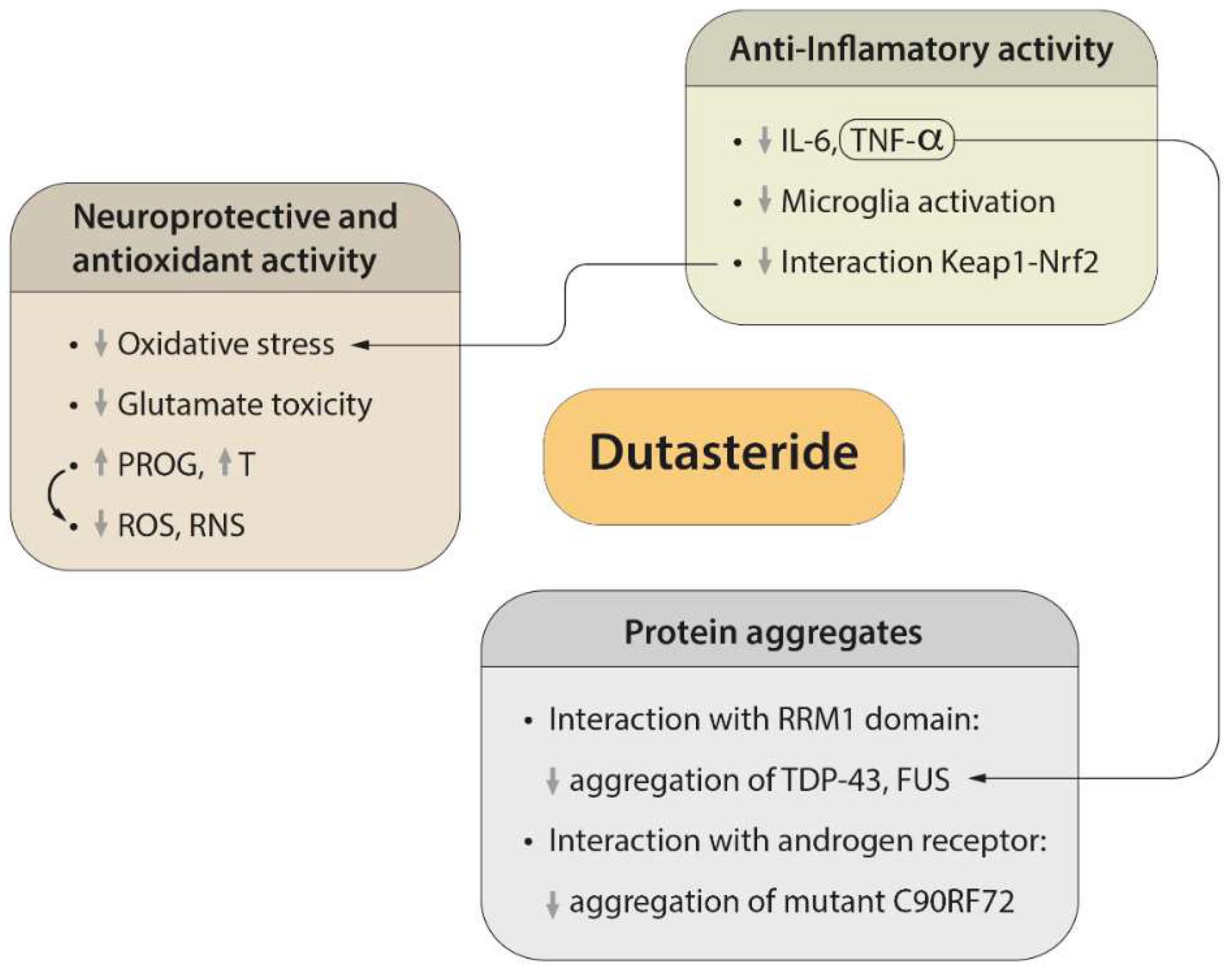
3. Therapeutic Effect of Dutasteride
3.1. Role of Dutasteride in the Activity of Steroid Hormones
3.2. Regulatory Mechanisms of Protein Aggregation
3.3. Neuroprotective and Antioxidant Effects of Dutasteride
3.4. Efficacy of Dutasteride against Neuroinflammation
3.5. Possible Side Effects of Dutasteride
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Logroscino, G.; Traynor, B.J.; Hardiman, O.; Chió, A.; Couratier, P.; Mitchell, J.D.; Swingler, R.J.; Beghi, E. Descriptive Epidemiology of Amyotrophic Lateral Sclerosis: New Evidence and Unsolved Issues. J. Neurol. Neurosurg. Psychiatry 2008, 79, 6–11. [Google Scholar] [CrossRef]
- Byers, J.S.; Huguenard, A.L.; Kuruppu, D.; Liu, N.K.; Xu, X.M.; Sengelaub, D.R. Neuroprotective Effects of Testosterone on Motoneuron and Muscle Morphology Following Spinal Cord Injury. J. Comp. Neurol. 2012, 520, 2683–2696. [Google Scholar] [CrossRef] [PubMed]
- Bensimon, G.; Lacomblez, L.; Meininger, V.; the ALS/Riluzole Study Group. A Controlled Trial of Riluzole in Amyotrophic Lateral Sclerosis. N. Engl. J. Med. 1994, 330, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, M.K. Riluzole and Edaravone: A Tale of Two Amyotrophic Lateral Sclerosis Drugs. Med. Res. Rev. 2019, 39, 733–748. [Google Scholar] [CrossRef] [PubMed]
- Galvez, J.; Llompart, J.; Land, D.; Pasinetti, G. Compositions for Treatment of Alzheimer’s Disease Using AB-Reducing and/or AB-Anti-Aggregation Compounds. CA Patent 2759187A1, 2010. [Google Scholar]
- Galvez, J.; Llompart, J.; Pal, K. N,N-Dicyclohexyl-(1S)-Isoborneol-10-Sulfonamide (MT103) and Related Compounds for the Treatment of Cancer. U.S. Patent 2004-836638, 2004. [Google Scholar]
- Galvez, J.; Llompart, J.; Pal, K. Treatment of Cancer with MT477 Derivatives. U.S. Patent 2004-60586519, 2006. [Google Scholar]
- Gastaminza, P.; Garaigorta, U.; Benlloch, J.M.; Galvez-Llompart, M.; Zanni, R.; Galvez, J. Compounds for the Treatment and Prevention of Viral Infections Caused by Coronaviruses. European Patent Application EP20382570.8, 2020. [Google Scholar]
- Galvez-Llompart, M.; Zanni, R.; Garcia-Domenech, R.; Galvez, J. How Molecular Topology Can Help in Amyotrophic Lateral Sclerosis (ALS) Drug Development: A Revolutionary Paradigm for a Merciless Disease. Pharmaceuticals 2022, 15, 94. [Google Scholar] [CrossRef]
- Frye, S. Discovery and Clinical Development of Dutasteride, a Potent Dual 5alpha-Reductase Inhibitor. Curr. Top. Med. Chem. 2006, 6, 405–421. [Google Scholar] [CrossRef]
- Fleshner, N.E.; Lucia, M.S.; Egerdie, B.; Aaron, L.; Eure, G.; Nandy, I.; Black, L.; Rittmaster, R.S. Dutasteride in Localised Prostate Cancer Management: The REDEEM Randomised, Double-Blind, Placebo-Controlled Trial. Lancet 2012, 379, 1103–1111. [Google Scholar] [CrossRef]
- Cadegiani, F.A.; McCoy, J.; Gustavo Wambier, C.; Goren, A. Early Antiandrogen Therapy With Dutasteride Reduces Viral Shedding, Inflammatory Responses, and Time-to-Remission in Males With COVID-19: A Randomized, Double-Blind, Placebo-Controlled Interventional Trial (EAT-DUTA AndroCoV Trial-Biochemical). Cureus 2021, 13, e13047. [Google Scholar] [CrossRef]
- Melcangi, R.C.; Garcia-Segura, L.M.; Mensah-Nyagan, A.G. Neuroactive Steroids: State of the Art and New Perspectives. Cell. Mol. Life Sci. 2008, 65, 777–797. [Google Scholar] [CrossRef]
- Giatti, S.; Garcia-Segura, L.M.; Melcangi, R.C. New Steps Forward in the Neuroactive Steroid Field. J. Steroid Biochem. Mol. Biol. 2015, 153, 127–134. [Google Scholar] [CrossRef]
- Blasco, H.; Guennoc, A.M.; Veyrat-Durebex, C.; Gordon, P.H.; Andres, C.R.; Camu, W.; Corcia, P. Amyotrophic Lateral Sclerosis: A Hormonal Condition? Amyotroph. Lateral Scler. 2012, 13, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Traxinger, K.; Kelly, C.; Johnson, B.A.; Lyles, R.H.; Glass, J.D. Prognosis and Epidemiology of Amyotrophic Lateral Sclerosis: Analysis of a Clinic Population, 1997–2011. Neurol. Clin. Pract. 2013, 3, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Kiernan, M.C.; Vucic, S.; Cheah, B.C.; Turner, M.R.; Eisen, A.; Hardiman, O.; Burrell, J.R.; Zoing, M.C. Amyotrophic Lateral Sclerosis. Lancet 2011, 377, 942–955. [Google Scholar] [CrossRef]
- Aggarwal, T.; Polanco, M.J.; Scaramuzzino, C.; Rocchi, A.; Milioto, C.; Emionite, L.; Ognio, E.; Sambataro, F.; Galbiati, M.; Poletti, A.; et al. Androgens Affect Muscle, Motor Neuron, and Survival in a Mouse Model of SOD1-Related Amyotrophic Lateral Sclerosis. Neurobiol. Aging 2014, 35, 1929–1938. [Google Scholar] [CrossRef]
- Manjaly, Z.R.; Scott, K.M.; Abhinav, K.; Wijesekera, L.; Ganesalingam, J.; Goldstein, L.H.; Janssen, A.; Dougherty, A.; Willey, E.; Stanton, B.R.; et al. The Sex Ratio in Amyotrophic Lateral Sclerosis: A Population Based Study. Amyotroph. Lateral Scler. 2010, 11, 439–442. [Google Scholar] [CrossRef]
- Poletti, A.; Coscarella, A.; Negri-Cesi, P.; Colciago, A.; Celotti, F.; Martini, L. 5a-Reductase Isozymes in the Central Nervous System. Steroids 1998, 63, 246–251. [Google Scholar] [CrossRef]
- Cermenati, G.; Giatti, S.; Audano, M.; Pesaresi, M.; Spezzano, R.; Caruso, D.; Mitro, N.; Melcangi, R.C. Diabetes Alters Myelin Lipid Profile in Rat Cerebral Cortex: Protective Effects of Dihydroprogesterone. J. Steroid Biochem. Mol. Biol. 2017, 168, 60–70. [Google Scholar] [CrossRef]
- Sumien, N.; Cunningham, J.T.; Davis, D.L.; Engelland, R.; Fadeyibi, O.; Farmer, G.E.; Mabry, S.; Mensah-Kane, P.; Trinh, O.T.P.; Vann, P.H.; et al. Neurodegenerative Disease: Roles for Sex, Hormones, and Oxidative Stress. Endocrinology 2021, 162, bqab185. [Google Scholar] [CrossRef]
- Vest, R.S.; Pike, C.J. Gender, Sex Steroid Hormones, and Alzheimer’s Disease. Horm. Behav. 2013, 63, 301–307. [Google Scholar] [CrossRef]
- Vegeto, E.; Villa, A.; Della Torre, S.; Crippa, V.; Rusmini, P.; Cristofani, R.; Galbiati, M.; Maggi, A.; Poletti, A. The Role of Sex and Sex Hormones in Neurodegenerative Diseases. Endocr. Rev. 2020, 41, 273–319. [Google Scholar] [CrossRef]
- De Vasconcelos, K.; Souza, A.; Oliveira, B.; Fernando, L.; Fuchs, P.; Simões, R.S.; João Batista, M.; Girão, C.; Soares Júnior, J.M.; Chada Baracat, E. Estrogens: Possible Protection against Amyotrophic Lateral Sclerosis? Rev. Assoc. Med. Bras. 2019, 65, 576–578. [Google Scholar] [CrossRef] [PubMed]
- McLeod, V.M.; Lau, C.L.; Chiam, M.D.F.F.; Rupasinghe, T.W.; Roessner, U.; Djouma, E.; Boon, W.C.; Turner, B.J.; Turner, C.J.B. Androgen Receptor Antagonism Accelerates Disease Onset in the SOD1 G93A Mouse Model of Amyotrophic Lateral Sclerosis. Br. J. Pharmacol. 2019, 176, 2111. [Google Scholar] [CrossRef]
- Scassellati, C.; Galoforo, A.C.; Esposito, C.; Ciani, M.; Ricevuti, G.; Bonvicini, C. Promising Intervention Approaches to Potentially Resolve Neuroinflammation And Steroid Hormones Alterations in Alzheimer’s Disease and Its Neuropsychiatric Symptoms. Aging Dis. 2021, 12, 1337–1357. [Google Scholar] [CrossRef] [PubMed]
- Rooney, J.P.K.; Visser, A.E.; D’Ovidio, F.; Vermeulen, R.; Beghi, E.; Chio, A.; Veldink, J.H.; Logroscino, G.; Van Den Berg, L.H.; Hardiman, O. A Case-Control Study of Hormonal Exposures as Etiologic Factors for ALS in Women: Euro-MOTOR. Neurology 2017, 89, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.; Gomes, C.; de Carvalho, M. Diagnosis, Pathogenesis and Therapeutic Targets in Amyotrophic Lateral Sclerosis. CNS Neurol. Disord. Drug Targets 2010, 9, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.; Kelley, D.B. Trophic Effects of Androgen: Receptor Expression and the Survival of Laryngeal Motor Neurons after Axotomy. J. Neurosci. 1996, 16, 6625. [Google Scholar] [CrossRef]
- Brooks, B.P.; Merry, D.E.; Paulson, H.L.; Lieberman, A.P.; Kolson, D.L.; Fischbeck, K.H. A Cell Culture Model for Androgen Effects in Motor Neurons. J. Neurochem. 1998, 70, 1054–1060. [Google Scholar] [CrossRef]
- Rothstein, J.D. Current Hypotheses for the Underlying Biology of Amyotrophic Lateral Sclerosis. Ann. Neurol. 2009, 65 (Suppl. S1), S3–S9. [Google Scholar] [CrossRef]
- Vivekananda, U.; Manjalay, Z.-R.R.; Ganesalingam, J.; Simms, J.; Shaw, C.E.; Leigh, P.N.; Turner, M.R.; Al-Chalabi, A. Low Index-to-Ring Finger Lenght Ratio in Sporadic ALS Supports Prenatally Defined Motor Neuronal Vulnerability. J. Neurol. Neurosurg. Psychiatry 2011, 82, 635–637. [Google Scholar] [CrossRef]
- Wicks, P. Hypothesis: Higher Prenatal Testosterone Predisposes ALS Patients to Improved Athletic Performance and Manual Professions. Amyotroph. Lateral Scler. 2012, 13, 251–253. [Google Scholar] [CrossRef] [Green Version]
- Behan, M.; Wenninger, J.M. Sex Steroidal Hormones and Respiratory Control. Respir. Physiol. Neurobiol. 2008, 164, 213. [Google Scholar] [CrossRef]
- Gargiulo-Monachelli, G.M.; Sivori, M.; Meyer, M.; Sica, R.E.P.P.; De Nicola, A.F.; Gonzalez-Deniselle, M.C.; De Nicola, A.F.; Gonzalez-Deniselle, M.C. Circulating Gonadal and Adrenal Steroids in Amyotrophic Lateral Sclerosis: Possible Markers of Susceptibility and Outcome. Horm. Metab. Res. 2014, 46, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Joseph, V.; Behan, M.; Kinkead, R. Sex, Hormones, and Stress: How They Impact Development and Function of the Carotid Bodies and Related Reflexes. Respir. Physiol. Neurobiol. 2013, 185, 75. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.E.; Rosenfeld, J.; Moore, D.H.; Bryan, W.W.; Barohn, R.J.; Wrench, M.; Myers, D.; Heberlin, L.; King, R.; Smith, J.; et al. A Preliminary Evaluation of a Prospective Study of Pulmonary Function Studies and Symptoms of Hypoventilation in ALS/MND Patients. J. Neurol. Sci. 2001, 191, 75–78. [Google Scholar] [CrossRef]
- Tatsumi, K.; Pickett, C.K.; Jacoby, C.R.; Weil, J.V.; Moore, L.G. Role of Endogenous Female Hormones in Hypoxic Chemosensitivity. J. Appl. Physiol. 1997, 83, 1706–1710. [Google Scholar] [CrossRef]
- Sawal, N.; Kaur, J.; Kaur, K.; Gombar, S. Dihydrotestosterone in Amyotrophic Lateral Sclerosis—The Missing Link? Brain Behav. 2020, 10, e01645. [Google Scholar] [CrossRef]
- Litim, N.; Bourque, M.; Al Sweidi, S.; Morissette, M.; Di Paolo, T. The 5α-Reductase Inhibitor Dutasteride but Not Finasteride Protects Dopamine Neurons in the MPTP Mouse Model of Parkinson’s Disease. Neuropharmacology 2015, 97, 86–94. [Google Scholar] [CrossRef]
- Militello, A.; Vitello, G.; Lunetta, C.; Toscano, A.; Maiorana, G.; Piccoli, T.; La Bella, V. The Serum Level of Free Testosterone Is Reduced in Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2002, 195, 67–70. [Google Scholar] [CrossRef]
- Loganathan, S.; Wilson, B.A.; Carey, S.B.; Manzo, E.; Joardar, A.; Ugur, B.; Zarnescu, D.C. TDP-43 Proteinopathy Causes Broad Metabolic Alterations Including TCA Cycle Intermediates and Dopamine Levels in Drosophila Models of ALS. Metabolites 2022, 12, 101. [Google Scholar] [CrossRef]
- Fu, X.; Zhu, W.; Guo, Z.; Shu, G.; Cui, F.; Yang, F.; Zhang, Y.; Ren, Y.; Zhang, X.; Zhang, X.; et al. 18 F-Fallypride PET-CT of Dopamine D2/D3 Receptors in Patients with Sporadic Amyotrophic Lateral Sclerosis. J. Neurol. Sci. 2017, 377, 79–84. [Google Scholar] [CrossRef]
- Dupuis, L.; Aguilar, J.L.; Oudart, H.; De, T.M.; Barbeito, L.; Loeffler, J.P.; Gonzalez De Aguilar, J.L.; Oudart, H.; De Tapia, M.; Barbeito, L.; et al. Mitochondria in Amyotrophic Lateral Sclerosis: A Trigger and a Target. Neurodegener. Dis. 2004, 1, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Vadakkadath Meethal, S.; Atwood, C.S. Lactate Dyscrasia: A Novel Explanation for Amyotrophic Lateral Sclerosis. Neurobiol. Aging 2012, 33, 569–581. [Google Scholar] [CrossRef]
- Jia, R.; Chen, Q.; Zhou, Q.; Zhang, R.; Jin, J.; Hu, F.; Liu, X.; Qin, X.; Kang, L.; Zhao, S.; et al. Characteristics of Serum Metabolites in Sporadic Amyotrophic Lateral Sclerosis Patients Based on Gas Chromatography-Mass Spectrometry. Sci. Rep. 2021, 11, 20786. [Google Scholar] [CrossRef]
- Echaniz-Laguna, A.; Zoll, J.; Ponsot, E.; N’Guessan, B.; Tranchant, C.; Loeffler, J.P.; Lampert, E.; Echaniz-Laguna, A.; Zoll, J.; Ponsot, E.; et al. Muscular Mitochondrial Function in Amyotrophic Lateral Sclerosis Is Progressively Altered as the Disease Develops: A Temporal Study in Man. Exp. Neurol. 2006, 198, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Dhaliwal, G.K.; Grewal, R.P. Mitochondrial DNA Deletion Mutation Levels Are Elevated in ALS Brains. Neuroreport 2000, 11, 2507–2509. [Google Scholar] [CrossRef] [PubMed]
- Grossini, E.; Garhwal, D.; Venkatesan, S.; Ferrante, D.; Mele, A.; Saraceno, M.; Scognamiglio, A.; Mandrioli, J.; Amedei, A.; De Marchi, F.; et al. The Potential Role of Peripheral Oxidative Stress on the Neurovascular Unit in Amyotrophic Lateral Sclerosis Pathogenesis: A Preliminary Report from Human and In Vitro Evaluations. Biomedicines 2022, 10, 691. [Google Scholar] [CrossRef]
- Blasco, H.; Garcon, G.; Patin, F.; Veyrat-Durebex, C.; Boyer, J.; Devos, D.; Vourc’H, P.; Andres, C.R.; Corcia, P. Panel of Oxidative Stress and Inflammatory Biomarkers in ALS: A Pilot Study. Can. J. Neurol. Sci. 2017, 44, 90–95. [Google Scholar] [CrossRef]
- Lall, D.; Baloh, R.H. Microglia and C9orf72 in Neuroinflammation and ALS and Frontotemporal Dementia. J. Clin. Investig. 2017, 127, 3250–3258. [Google Scholar] [CrossRef]
- Shatunov, A.; Al-Chalabi, A. The Genetic Architecture of ALS. Neurobiol. Dis. 2021, 147, 105156. [Google Scholar] [CrossRef]
- Benson, B.C.; Shaw, P.J.; Azzouz, M.; Highley, J.R.; Hautbergue, G.M. Proteinopathies as Hallmarks of Impaired Gene Expression, Proteostasis and Mitochondrial Function in Amyotrophic Lateral Sclerosis. Front. Neurosci. 2021, 15, 1738. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, D.; Malik, A.N.; Kulac, I.; Basak, A.N.; Lazoglu, I.; Ulusu, N.N. Impact of the Amyotrophic Lateral Sclerosis Disease on the Biomechanical Properties and Oxidative Stress Metabolism of the Lung Tissue Correlated With the Human Mutant SOD1 G93A Protein Accumulation. Front. Bioeng. Biotechnol. 2022, 10, 810243. [Google Scholar] [CrossRef] [PubMed]
- McAlary, L.; Shephard, V.K.; Wright, G.S.A.; Yerbury, J.J. A Copper Chaperone-Mimetic Polytherapy for SOD1-Associated Amyotrophic Lateral Sclerosis. J. Biol. Chem. 2022, 298, 101612. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.N.; Wu, Y.; Odeh, H.M.; Gendron, T.F.; Jansen-West, K.; del Rosso, G.; Yue, M.; Jiang, P.; Gomes, E.; Tong, J.; et al. C9orf72 Poly(GR) Aggregation Induces TDP-43 Proteinopathy. Sci. Transl. Med. 2020, 12, eabb3774. [Google Scholar] [CrossRef]
- Majounie, E.; Renton, A.E.; Mok, K.; Dopper, E.G.P.; Waite, A.; Rollinson, S.; Chiò, A.; Restagno, G.; Nicolaou, N.; Simon-Sanchez, J.; et al. Frequency of the C9orf72 Hexanucleotide Repeat Expansion in Patients with Amyotrophic Lateral Sclerosis and Frontotemporal Dementia: A Cross-Sectional Study. Lancet Neurol. 2012, 11, 323–330. [Google Scholar] [CrossRef]
- Davidson, Y.; Kelley, T.; Mackenzie, I.R.A.; Pickering-Brown, S.; Du Plessis, D.; Neary, D.; Snowden, J.S.; Mann, D.M.A. Ubiquitinated Pathological Lesions in Frontotemporal Lobar Degeneration Contain the TAR DNA-Binding Protein, TDP-43. Acta Neuropathol. 2007, 113, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.K.; Chiang, M.H.; Toh, E.K.W.; Chang, C.F.; Huang, T.H. Molecular Mechanism of Oxidation-Induced TDP-43 RRM1 Aggregation and Loss of Function. FEBS Lett. 2013, 587, 575–582. [Google Scholar] [CrossRef] [PubMed]
- McAlary, L.; Yerbury, J.J.; Cashman, N.R. The Prion-like Nature of Amyotrophic Lateral Sclerosis. Prog. Mol. Biol. Transl. Sci. 2020, 175, 261–296. [Google Scholar] [CrossRef]
- Cicardi, M.E.; Marrone, L.; Azzouz, M.; Trotti, D. Proteostatic Imbalance and Protein Spreading in Amyotrophic Lateral Sclerosis. EMBO J. 2021, 40, e106389. [Google Scholar] [CrossRef]
- Basu, S.; Rajendra, K.C.; Alagar, S.; Bahadur, R.P. Impaired Nuclear Transport Induced by Juvenile ALS Causing P525L Mutation in NLS Domain of FUS: A Molecular Mechanistic Study. Biochim. Biophys. Acta Proteins Proteom. 2022, 1870, 140766. [Google Scholar] [CrossRef]
- Jensen, B.K.; McAvoy, K.J.; Heinsinger, N.M.; Lepore, A.C.; Ilieva, H.; Haeusler, A.R.; Trotti, D.; Pasinelli, P. Targeting TNFα Produced by Astrocytes Expressing Amyotrophic Lateral Sclerosis-Linked Mutant Fused in Sarcoma Prevents Neurodegeneration and Motor Dysfunction in Mice. Glia 2022, 70, 1426–1449. [Google Scholar] [CrossRef]
- Ajmone-Cat, M.A.; Onori, A.; Toselli, C.; Stronati, E.; Morlando, M.; Bozzoni, I.; Monni, E.; Kokaia, Z.; Lupo, G.; Minghetti, L.; et al. Increased FUS Levels in Astrocytes Leads to Astrocyte and Microglia Activation and Neuronal Death. Sci. Rep. 2019, 9, 4572. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.S.; Fuentealba, R.A.; Miller, S.E.; Jackson, E.; Piwnica-Worms, D.; Baloh, R.H.; Weihl, C.C. Valosin-Containing Protein (VCP) Is Required for Autophagy and Is Disrupted in VCP Disease. J. Cell Biol. 2009, 187, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, F. Role of Neuroinflammation in Amyotrophic Lateral Sclerosis: Cellular Mechanisms and Therapeutic Implications. Front. Immunol. 2017, 8, 1005. [Google Scholar] [CrossRef]
- Filiano, A.J.; Gadani, S.P.; Kipnis, J. Interactions of Innate and Adaptive Immunity in Brain Development and Function. Brain Res. 2015, 1617, 18–27. [Google Scholar] [CrossRef]
- Ransohoff, R.M. How Neuroinflammation Contributes to Neurodegeneration. Science 2016, 353, 777–783. [Google Scholar] [CrossRef]
- Frakes, A.E.; Ferraiuolo, L.; Haidet-Phillips, A.M.; Schmelzer, L.; Braun, L.; Miranda, C.J.; Ladner, K.J.; Bevan, A.K.; Foust, K.D.; Godbout, J.P.; et al. Microglia Induce Motor Neuron Death via the Classical NF-ΚB Pathway in Amyotrophic Lateral Sclerosis. Neuron 2014, 81, 1009–1023. [Google Scholar] [CrossRef] [PubMed]
- Keleku-Lukwete, N.; Suzuki, M.; Yamamoto, M. An Overview of the Advantages of KEAP1-NRF2 System Activation During Inflammatory Disease Treatment. Antioxid. Redox Signal. 2018, 29, 1746–1755. [Google Scholar] [CrossRef]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 Suppresses Macrophage Inflammatory Response by Blocking Proinflammatory Cytokine Transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef]
- Thimmulappa, R.K.; Scollick, C.; Traore, K.; Yates, M.; Trush, M.A.; Liby, K.T.; Sporn, M.B.; Yamamoto, M.; Kensler, T.W.; Biswal, S. Nrf2-Dependent Protection from LPS Induced Inflammatory Response and Mortality by CDDO-Imidazolide. Biochem. Biophys. Res. Commun. 2006, 351, 883–889. [Google Scholar] [CrossRef]
- Lu, M.C.; Zhao, J.; Liu, Y.T.; Liu, T.; Tao, M.M.; You, Q.D.; Jiang, Z.Y. CPUY192018, a Potent Inhibitor of the Keap1-Nrf2 Protein-Protein Interaction, Alleviates Renal Inflammation in Mice by Restricting Oxidative Stress and NF-ΚB Activation. Redox Biol. 2019, 26, 101266. [Google Scholar] [CrossRef]
- Prell, T.; Lautenschläger, J.; Weidemann, L.; Ruhmer, J.; Witte, O.W.; Grosskreutz, J. Endoplasmic Reticulum Stress Is Accompanied by Activation of NF-ΚB in Amyotrophic Lateral Sclerosis. J. Neuroimmunol. 2014, 270, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.C.; Rosario, E.R. The Potential Use of Hormone-Based Therapeutics for the Treatment of Alzheimer’s Disease. Curr. Alzheimer Res. 2012, 9, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.V.; Hermann, D.J.; Cunningham, G.R.; Wilson, T.H.; Morrill, B.B.; Hobbs, S. Marked Suppression of Dihydrotestosterone in Men with Benign Prostatic Hyperplasia by Dutasteride, a Dual 5α-Reductase Inhibitor. J. Clin. Endocrinol. Metab. 2004, 89, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Penning, T.M. Steroid 5alpha-Reductases and 3alpha-Hydroxysteroid Dehydrogenases: Key Enzymes in Androgen Metabolism. Best Pract. Res. Clin. Endocrinol. Metab. 2001, 15, 79–94. [Google Scholar] [CrossRef]
- Monachelli, G.G.; Meyer, M.; Rodríguez, G.E.; Garay, L.I.; Sica, R.E.P.; De Nicola, A.F.; Deniselle, M.C.G. Endogenous Progesterone Is Associated to Amyotrophic Lateral Sclerosis Prognostic Factors. Acta Neurol. Scand. 2011, 123, 60–67. [Google Scholar] [CrossRef]
- Schumacher, M.; Denier, C.; Oudinet, J.P.; Adams, D.; Guennoun, R. Progesterone Neuroprotection: The Background of Clinical Trial Failure. J. Steroid Biochem. Mol. Biol. 2016, 160, 53–56. [Google Scholar] [CrossRef]
- Wright, D.W.; Kellermann, A.L.; Hertzberg, V.S.; Clark, P.L.; Frankel, M.; Goldstein, F.C.; Salomone, J.P.; Dent, L.L.; Harris, O.A.; Ander, D.S.; et al. ProTECT: A Randomized Clinical Trial of Progesterone for Acute Traumatic Brain Injury. Ann. Emerg. Med. 2007, 49, 391–402. [Google Scholar] [CrossRef]
- De Jong, S.; Huisman, M.; Sutedja, N.; Van Der Kooi, A.; De Visser, M.; Schelhaas, J.; Van Der Schouw, Y.; Veldink, J.; Van Den Berg, L.; De Jong, S.; et al. Endogenous Female Reproductive Hormones and the Risk of Amyotrophic Lateral Sclerosis. J. Neurol. 2013, 260, 507–512. [Google Scholar] [CrossRef]
- Hancevic, M.; Bilic, H.; Sitas, B.; Pavlisa, G.; Borovecki, F.; Munitic, I.; Bilic, E. Attenuation of ALS Progression during Pregnancy—Lessons to Be Learned or Just a Coincidence? Neurol. Sci. 2019, 40, 1275–1278. [Google Scholar] [CrossRef]
- Mhaouty-Kodja, S. Role of the Androgen Receptor in the Central Nervous System. Mol. Cell. Endocrinol. 2018, 465, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-Mediated Neuroinflammation in Neurodegenerative Diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Mouihate, A. TLR4-Mediated Brain Inflammation Halts Neurogenesis: Impact of Hormonal Replacement Therapy. Front. Cell. Neurosci. 2014, 8, 146. [Google Scholar] [CrossRef] [PubMed]
- Novak, C.M.; Ozen, M.; McLane, M.; Alqutub, S.; Lee, J.Y.; Lei, J.; Burd, I. Progesterone Improves Perinatal Neuromotor Outcomes in a Mouse Model of Intrauterine Inflammation via Immunomodulation of the Placenta. Am. J. Reprod. Immunol. 2018, 79, e12842. [Google Scholar] [CrossRef] [PubMed]
- Atallah, A.; Mhaouty-Kodja, S.; Grange-Messent, V. Chronic Depletion of Gonadal Testosterone Leads to Blood-Brain Barrier Dysfunction and Inflammation in Male Mice. J. Cereb. Blood Flow Metab. 2017, 37, 3161–3175. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Roet, K.C.D.; Zhang, L.; Brault, A.; Berg, A.P.; Jefferson, A.B.; Klug-McLeod, J.; Leach, K.L.; Vincent, F.; Yang, H.; et al. Human Amyotrophic Lateral Sclerosis Excitability Phenotype Screen: Target Discovery and Validation. Cell Rep. 2021, 35, 109224. [Google Scholar] [CrossRef]
- Litim, N.; Morissette, M.; Caruso, D.; Melcangi, R.C.; Di Paolo, T. Effect of the 5α-Reductase Enzyme Inhibitor Dutasteride in the Brain of Intact and Parkinsonian Mice. J. Steroid Biochem. Mol. Biol. 2017, 174, 242–256. [Google Scholar] [CrossRef]
- Bianchi, V.E.; Rizzi, L.; Bresciani, E.; Omeljaniuk, R.J.; Torsello, A. Androgen Therapy in Neurodegenerative Diseases. J. Endocr. Soc. 2020, 4, bvaa120. [Google Scholar] [CrossRef]
- Fargo, K.N.; Foecking, E.M.; Jones, K.J.; Sengelaub, D.R.; Fargo, K.N.; Galbiati, M.; Foecking, E.M.; Poletti, A.; Jones, K.J. Neuroprotective Actions of Androgens on Motoneurons. Front. Neuroendocr. 2009, 30, 130–141. [Google Scholar] [CrossRef]
- Wilborn, C.; Taylor, L.; Poole, C.; Foster, C.; Willoughby, D.; Kreider, R. Effects of a Purported Aromatase and 5α-Reductase Inhibitor on Hormone Profiles in College-Age Men. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 457–465. [Google Scholar] [CrossRef]
- Kim, Y.J.; Soto, M.; Branigan, G.L.; Rodgers, K.; Brinton, R.D. Association between Menopausal Hormone Therapy and Risk of Neurodegenerative Diseases: Implications for Precision Hormone Therapy. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2021, 7, e12174. [Google Scholar] [CrossRef]
- Lukavsky, P.J.; Daujotyte, D.; Tollervey, J.R.; Ule, J.; Stuani, C.; Buratti, E.; Baralle, F.E.; Damberger, F.F.; Allain, F.H.T. Molecular Basis of UG-Rich RNA Recognition by the Human Splicing Factor TDP-43. Nat. Struct. Mol. Biol. 2013, 20, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.H.; Chiang, C.H.; Wang, Y.T.; Doudeva, L.G.; Yuan, H.S. The Crystal Structure of TDP-43 RRM1-DNA Complex Reveals the Specific Recognition for UG- and TG-Rich Nucleic Acids. Nucleic Acids Res. 2014, 42, 4712–4722. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, C.; Shan, J.; Wang, Y.; Chen, G. Insights into the Aggregation Mechanism of RNA Recognition Motif Domains in TDP-43: A Theoretical Exploration. R. Soc. Open Sci. 2021, 8, 210160. [Google Scholar] [CrossRef] [PubMed]
- McLeod, V.M.; Chiam, M.D.F.; Perera, N.D.; Lau, C.L.; Boon, W.C.; Turner, B.J. Mapping Motor Neuron Vulnerability in the Neuraxis of Male SOD1G93A Mice Reveals Widespread Loss of Androgen Receptor Occurring Early in Spinal Motor Neurons. Front. Endocrinol. 2022, 13, 122. [Google Scholar] [CrossRef] [PubMed]
- McLeod, V.M.; Chiam, M.D.F.; Lau, C.L.; Rupasinghe, T.W.; Boon, W.C.; Turner, B.J. Dysregulation of Steroid Hormone Receptors in Motor Neurons and Glia Associates with Disease Progression in ALS Mice. Endocrinology 2020, 161, bqaa113. [Google Scholar] [CrossRef]
- Fujita, K.; Nakamura, Y.; Oka, T.; Ito, H.; Tamura, T.; Tagawa, K.; Sasabe, T.; Katsuta, A.; Motoki, K.; Shiwaku, H.; et al. A Functional Deficiency of TERA/VCP/P97 Contributes to Impaired DNA Repair in Multiple Polyglutamine Diseases. Nat. Commun. 2013, 4, 1816. [Google Scholar] [CrossRef]
- Zhou, F.; Dong, H.; Liu, Y.; Yan, L.; Sun, C.; Hao, P.; Liu, Y.; Zhai, J.; Liu, Y. Raloxifene, a Promising Estrogen Replacement, Limits TDP-25 Cell Death by Enhancing Autophagy and Suppressing Apoptosis. Brain Res. Bull. 2018, 140, 281–290. [Google Scholar] [CrossRef]
- Boddy, S.; Islam, M.; Moll, T.; Kurz, J.; Burrows, D.; McGown, A.; Bhargava, A.; Julian, T.H.; Harvey, C.; Marshall, J.N.; et al. Unbiased Metabolome Screen Leads to Personalized Medicine Strategy for Amyotrophic Lateral Sclerosis. Brain Commun. 2022, 4, fcac069. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: A Concept in Redox Biology and Medicine. Redox Biol. 2015, 4, 180. [Google Scholar] [CrossRef] [Green Version]
- D’Ambrosi, N.; Cozzolino, M.; Carrì, M.T. Neuroinflammation in Amyotrophic Lateral Sclerosis: Role of Redox (Dys)Regulation. Antioxid. Redox Signal. 2018, 29, 15–36. [Google Scholar] [CrossRef] [PubMed]
- Merz, S.F.; Bengtson, C.P.; Tepohl, C.; Hagenston, A.M.; Bading, H.; Bas-Orth, C. A Microscopy-Based Small Molecule Screen in Primary Neurons Reveals Neuroprotective Properties of the FDA-Approved Anti-Viral Drug Elvitegravir. Mol. Brain 2020, 13, 124. [Google Scholar] [CrossRef] [PubMed]
- Soskic, V.; Schrattenholz, A. WO2006058781A2-Finasteride, Dutasteride and Related Compounds for Preventing/Treating Neurologically-Associated Disorders-Google Patents. Available online: https://patents.google.com/patent/WO2006058781A2/en (accessed on 17 August 2022).
- Meydan, S.; Kus, I.; Tas, U. Effects of Testosterone on Orchiectomy-Induced Oxidative Damage in the Rat Hippocampus. J. Chem. Neuroanat. 2010, 40, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Son, S.W.; Lee, J.S.; Kim, H.G.; Kim, D.W.; Ahn, Y.C.; Son, C.G. Testosterone Depletion Increases the Susceptibility of Brain Tissue to Oxidative Damage in a Restraint Stress Mouse Model. J. Neurochem. 2016, 136, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Toro-Urrego, N.; Garcia-Segura, L.M.; Echeverria, V.; Barreto, G.E. Testosterone Protects Mitochondrial Function and Regulates Neuroglobin Expression in Astrocytic Cells Exposed to Glucose Deprivation. Front. Aging Neurosci. 2016, 8, 152. [Google Scholar] [CrossRef]
- Ota, H.; Akishita, M.; Akiyoshi, T. Testosterone Deficiency Accelerates Neuronal and Vascular Aging of SAMP8 Mice: Protective Role of ENOS and SIRT1. PLoS ONE 2012, 7, e29598. [Google Scholar]
- Matsumoto, A. Hormonally Induced Neuronal Plasticity in the Adult Motoneurons. Brain Res. Bull. 1997, 44, 539–547. [Google Scholar] [CrossRef]
- Leranth, C.; Petnehazy, O.; MacLusky, N.J. Gonadal Hormones Affect Spine Synaptic Density in the CA1 Hippocampal Subfield of Male Rats. J. Neurosci. 2003, 23, 1588–1592. [Google Scholar] [CrossRef]
- Beyer, C.; Hutchison, J.B. Androgens Stimulate the Morphological Maturation of Embryonic Hypothalamic Aromatase-Immunoreactive Neurons in the Mouse. Brain Res. Dev. Brain Res. 1997, 98, 74–81. [Google Scholar] [CrossRef]
- Marron, T.U.; Guerini, V.; Rusmini, P. Androgen-Induced Neurite Outgrowth Is Mediated by Neuritin in Motor Neurones. J. Neurochem. 2005, 92, 10–20. [Google Scholar] [CrossRef]
- Hong, Y.; Liu, Y.; Yu, D.; Wang, M.; Hou, Y. The Neuroprotection of Progesterone against Aβ-Induced NLRP3-Caspase-1 Inflammasome Activation via Enhancing Autophagy in Astrocytes. Int. Immunopharmacol. 2019, 74, 105669. [Google Scholar] [CrossRef] [PubMed]
- De Nicola, A.F.; Meyer, M.; Garay, L.; Kruse, M.S.; Schumacher, M.; Guennoun, R.; Gonzalez Deniselle, M.C. Progesterone and Allopregnanolone Neuroprotective Effects in the Wobbler Mouse Model of Amyotrophic Lateral Sclerosis. Cell. Mol. Neurobiol. 2022, 42, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rhodes, L.E.; Kokkinis, A.D.; White, M.J.; Watts, C.A.; Auh, S.; Jeffries, N.O.; Shrader, J.A.; Lehky, T.J.; Li, L.; Ryder, J.E.; et al. Efficacy and Safety of Dutasteride in Patients with Spinal and Bulbar Muscular Atrophy: A Randomised Placebo-Controlled Trial. Lancet. Neurol. 2011, 10, 140–147. [Google Scholar] [CrossRef]
- Mizoguchi, S.; Mori, K.; Shin, T.; Wang, Z.; DeFranco, D.B.; Yoshimura, N.; Mimata, H. Effects of Dutasteride in a Rat Model of Chemically Induced Prostatic Inflammation—Potential Role of Estrogen Receptor β. Prostate 2020, 80, 1413. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Han, L.; Gao, S.; Xiao, Z.; Zhou, Q.; Cheng, X.; Zhang, Y.; Zhou, W. Lincs Dataset-Based Repositioning of Dutasteride as an Anti-Neuroinflammation Agent. Brain Sci. 2021, 11, 1411. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, C.Y.; Lin, H.Q.; Hu, J.S.; Ip, T.M.; Chi-Cheong Wan, D. Development of an Enzyme-Linked Immunosorbent Assay for Keap1-Nrf2 Interaction Inhibitors Identification. Redox Biol. 2020, 34, 101573. [Google Scholar] [CrossRef]
- Hirshburg, J.M.; Kelsey, P.A.; Therrien, C.A.; Gavino, A.C.; Reichenberg, J.S. Adverse Effects and Safety of 5-Alpha Reductase Inhibitors (Finasteride, Dutasteride): A Systematic Review-PubMed. J. Clin. Aesthet Dermatol 2016, 9, 56–62. [Google Scholar]
- Roehrborn, C.G.; Boyle, P.; Nickel, J.C.; Hoefner, K.; Andriole, G. Efficacy and Safety of a Dual Inhibitor of 5-Alpha-Reductase Types 1 and 2 (Dutasteride) in Men with Benign Prostatic Hyperplasia. Urology 2002, 60, 434–441. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proaño, B.; Casani-Cubel, J.; Benlloch, M.; Rodriguez-Mateos, A.; Navarro-Illana, E.; Lajara-Romance, J.M.; de la Rubia Ortí, J.E. Is Dutasteride a Therapeutic Alternative for Amyotrophic Lateral Sclerosis? Biomedicines 2022, 10, 2084. https://doi.org/10.3390/biomedicines10092084
Proaño B, Casani-Cubel J, Benlloch M, Rodriguez-Mateos A, Navarro-Illana E, Lajara-Romance JM, de la Rubia Ortí JE. Is Dutasteride a Therapeutic Alternative for Amyotrophic Lateral Sclerosis? Biomedicines. 2022; 10(9):2084. https://doi.org/10.3390/biomedicines10092084
Chicago/Turabian StyleProaño, Belén, Julia Casani-Cubel, María Benlloch, Ana Rodriguez-Mateos, Esther Navarro-Illana, Jose María Lajara-Romance, and Jose Enrique de la Rubia Ortí. 2022. "Is Dutasteride a Therapeutic Alternative for Amyotrophic Lateral Sclerosis?" Biomedicines 10, no. 9: 2084. https://doi.org/10.3390/biomedicines10092084