
Identification of the Natural Steroid Sapogenin Diosgenin as a Direct Dual-Specific RORα/γ Inverse Agonist

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines, Plasmids and Chemicals
2.2. Cell Culture
2.3. Luciferase Assays
2.4. Resazurin Conversion Assay
2.5. Determination of Target Gene Expression by RT-qPCR
2.6. In Silico Docking
2.7. Statistical Analysis
3. Results
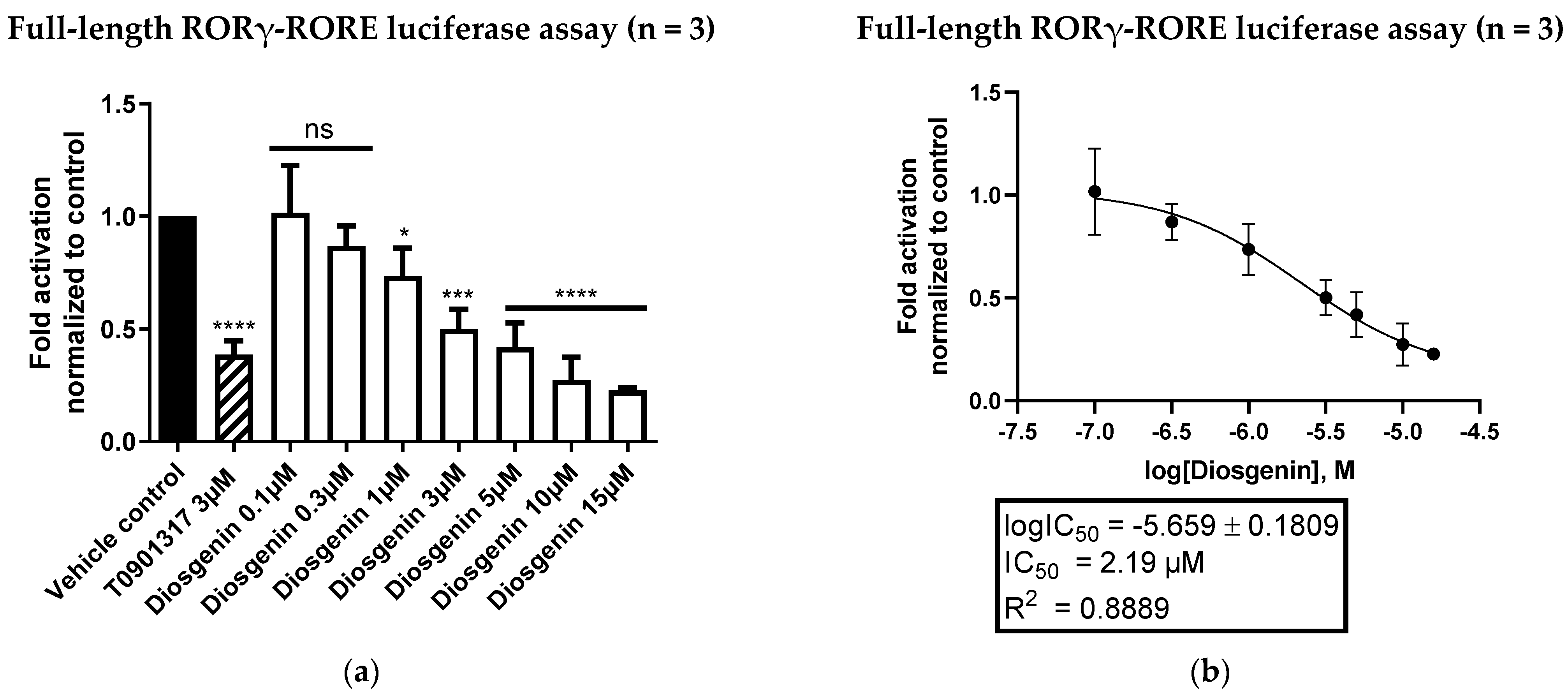
3.1. Diosgenin Is a Potent and Direct Inverse Agonist of the Nuclear Receptor RORγ
3.2. Diosgenin Shows No Signs of Cytotoxicity in a Resazurin Conversion Assay
3.3. The Steroid Saponins Dioscin and Protodioscin Do Not Act as ROR Inverse Agonists in Non-Toxic Concentrations
3.4. Diosgenin Is a Dual Specific Inhibitor of RORα and RORγ, but Shows No Activity on RORβ or Other Important Nuclear Receptors
3.5. Diosgenin Downregulates ROR Target Gene Expression in Functional Cellular Models
3.6. In Silico Modelling of the Binding Mode of Diosgenin to the Ligand-Binding Domain of RORγ
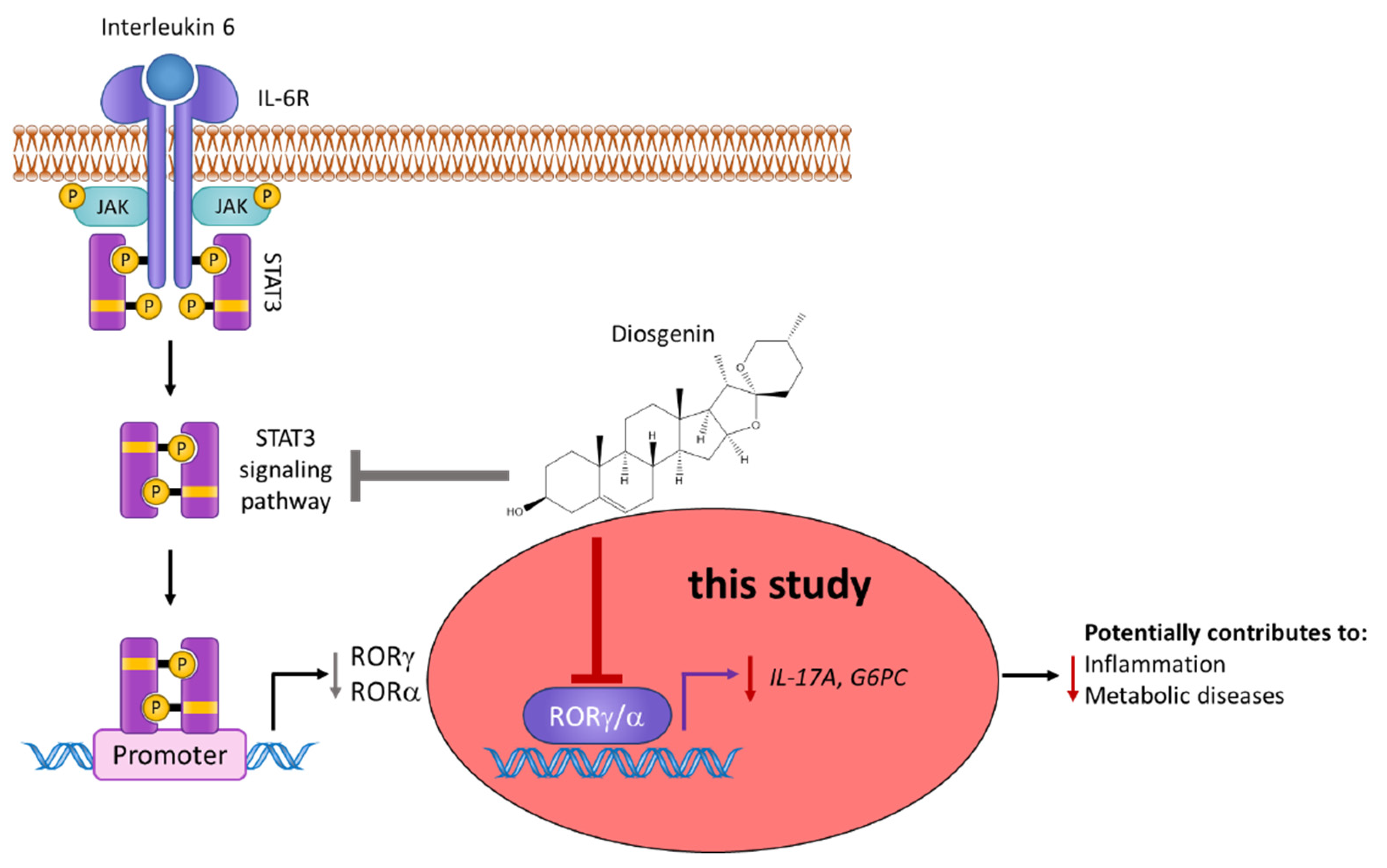
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Solt, L.A.; Griffin, P.R.; Burris, T.P. Ligand regulation of retinoic acid receptor-related orphan receptors: Implications for development of novel therapeutics. Curr. Opin. Lipidol. 2010, 21, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Stehlin, C.; Wurtz, J.-M.; Steinmetz, A.; Greiner, E.; Schüle, R.; Moras, D.; Renaud, J.-P. X-ray structure of the orphan nuclear receptor RORβ ligand-binding domain in the active conformation. EMBO J. 2001, 20, 5822–5831. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E.; Lepelley, A.; Lafaille, J.J.; Cua, D.J.; Littman, D.R. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006, 126, 1121–1133. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.I.; Zhou, L.; Littman, D.R. Transcriptional regulation of Th17 cell differentiation. Semin. Immunol. 2007, 19, 409–417. [Google Scholar] [CrossRef]
- Takeda, Y.; Kang, H.S.; Freudenberg, J.; DeGraff, L.M.; Jothi, R.; Jetten, A.M. Retinoic acid-related orphan receptor gamma (RORgamma): A novel participant in the diurnal regulation of hepatic gluconeogenesis and insulin sensitivity. PLOS Genet. 2014, 10, e1004331. [Google Scholar] [CrossRef]
- Guendisch, U.; Weiss, J.; Ecoeur, F.; Riker, J.C.; Kaupmann, K.; Kallen, J.; Hintermann, S.; Orain, D.; Dawson, J.; Billich, A.; et al. Pharmacological inhibition of RORgammat suppresses the Th17 pathway and alleviates arthritis in vivo. PLoS ONE 2017, 12, e0188391. [Google Scholar] [CrossRef]
- Kumar, N.; Lyda, B.; Chang, M.R.; Lauer, J.L.; Solt, L.A.; Burris, T.P.; Kamenecka, T.M.; Griffin, P.R. Identification of SR2211: A potent synthetic RORgamma-selective modulator. ACS Chem. Biol. 2012, 7, 672–677. [Google Scholar] [CrossRef]
- Solt, L.A.; Kumar, N.; Nuhant, P.; Wang, Y.; Lauer, J.L.; Liu, J.; Istrate, M.A.; Kamenecka, T.M.; Roush, W.R.; Vidović, D.; et al. Suppression of TH17 differentiation and autoimmunity by a synthetic ROR ligand. Nature 2011, 472, 491–494. [Google Scholar] [CrossRef]
- Jetten, A.M.; Kang, H.S.; Takeda, Y. Retinoic acid-related orphan receptors alpha and gamma: Key regulators of lipid/glucose metabolism, inflammation, and insulin sensitivity. Front. Endocrinol. 2013, 4, 1. [Google Scholar] [CrossRef]
- Yang, X.O.; Pappu, B.P.; Nurieva, R.; Akimzhanov, A.; Kang, H.S.; Chung, Y.; Ma, L.; Shah, B.; Panopoulos, A.D.; Schluns, K.S.; et al. T helper 17 lineage differentiation is programmed by orphan nuclear receptors ROR alpha and ROR gamma. Immunity 2008, 28, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Lau, P.; Fitzsimmons, R.L.; Raichur, S.; Wang, S.C.; Lechtken, A.; Muscat, G.E. The orphan nuclear receptor, RORalpha, regulates gene expression that controls lipid metabolism: Staggerer (SG/SG) mice are resistant to diet-induced obesity. J. Biol. Chem. 2008, 283, 18411–18421. [Google Scholar] [CrossRef] [PubMed]
- Kadiri, S.; Monnier, C.; Ganbold, M.; Ledent, T.; Capeau, J.; Antoine, B. The nuclear retinoid-related orphan receptor-alpha regulates adipose tissue glyceroneogenesis in addition to hepatic gluconeogenesis. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E105–E114. [Google Scholar] [CrossRef]
- Lau, P.; Fitzsimmons, R.L.; Pearen, M.A.; Watt, M.J.; Muscat, G.E. Homozygous staggerer (sg/sg) mice display improved insulin sensitivity and enhanced glucose uptake in skeletal muscle. Diabetologia 2011, 54, 1169–1180. [Google Scholar] [CrossRef]
- Chopra, A.R.; Louet, J.F.; Saha, P.; An, J.; Demayo, F.; Xu, J.; York, B.; Karpen, S.; Finegold, M.; Moore, D.; et al. Absence of the SRC-2 coactivator results in a glycogenopathy resembling Von Gierke’s disease. Science 2008, 322, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- Dong, C. TH17 cells in development: An updated view of their molecular identity and genetic programming. Nat. Rev. Immunol. 2008, 8, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.R.; Littman, D.R. Small molecule inhibitors of RORgammat: Targeting Th17 cells and other applications. Eur. J. Immunol. 2012, 42, 2232–2237. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Downes, M.; Yu, R.T.; Bookout, A.L.; He, W.; Straume, M.; Mangelsdorf, D.J.; Evans, R.M. Nuclear receptor expression links the circadian clock to metabolism. Cell 2006, 126, 801–810. [Google Scholar] [CrossRef]
- Kang, H.S.; Angers, M.; Beak, J.Y.; Wu, X.; Gimble, J.M.; Wada, T.; Xie, W.; Collins, J.B.; Grissom, S.F.; Jetten, A.M. Gene expression profiling reveals a regulatory role for ROR alpha and ROR gamma in phase I and phase II metabolism. Physiol. Genom. 2007, 31, 281–294. [Google Scholar] [CrossRef]
- Tai Lahans, L. Integrating Conventional and Chinese Medicine in Cancer Care: A Clinical Guide; Elsevier Health Sciences: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Yi, T.; Fan, L.L.; Chen, H.L.; Zhu, G.Y.; Suen, H.M.; Tang, Y.N.; Zhu, L.; Chu, C.; Zhao, Z.Z.; Chen, H.B. Comparative analysis of diosgenin in Dioscorea species and related medicinal plants by UPLC-DAD-MS. BMC Biochem. 2014, 15, 19. [Google Scholar] [CrossRef]
- Tsukamoto, T.; Ueno, Y.; Ohta, Z. Diosgenin II. Glucoside of Dioscorea tokoro Makino. 3. Constitution of diosgenin. J. Pharm. Soc. Jpn. 1937, 57, 985–991. [Google Scholar] [CrossRef] [Green Version]
- Marker, R.E.; Krueger, J. Sterols. CXII. Sapogenins. XLI. The Preparation of Trillin and its Conversion to Progesterone. J. Am. Chem. Soc. 1940, 62, 3349–3350. [Google Scholar] [CrossRef]
- Lemin, A.J.; Djerassi, C. The Conversion of Diosgenin to Cortisone via 11-Ketosteroids of the 5β-Series. J. Am. Chem. Soc. 1954, 76, 5672–5674. [Google Scholar] [CrossRef]
- Jesus, M.; Martins, A.P.; Gallardo, E.; Silvestre, S. Diosgenin: Recent Highlights on Pharmacology and Analytical Methodology. J. Anal. Methods Chem. 2016, 2016, 4156293. [Google Scholar] [CrossRef]
- Kim, J.K.; Park, S.U. An update on the biological and pharmacological activities of diosgenin. EXCLI J. 2018, 17, 24–28. [Google Scholar] [CrossRef]
- Liu, W.; Zhu, M.; Yu, Z.; Yin, D.; Lu, F.; Pu, Y.; Zhao, C.; He, C.; Cao, L. Therapeutic effects of diosgenin in experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2017, 313, 152–160. [Google Scholar] [CrossRef]
- Cao, Y.J.; Xu, Y.; Liu, B.; Zheng, X.; Wu, J.; Zhang, Y.; Li, X.S.; Qi, Y.; Sun, Y.M.; Wen, W.B.; et al. Dioscin, a Steroidal Saponin Isolated from Dioscorea nipponica, Attenuates Collagen-Induced Arthritis by Inhibiting Th17 Cell Response. Am. J. Chin. Med. 2019, 47, 423–437. [Google Scholar] [CrossRef]
- Xing, E.; Guo, Y.; Feng, G.; Song, H.; An, G.; Zhao, X.; Wang, M. Effects of dioscin on T helper 17 and regulatory T-cell subsets in chicken collagen type II-induced arthritis mice. J. Chin. Med. Assoc. 2019, 82, 202–208. [Google Scholar] [CrossRef]
- Song, H.; Gao, Y.; Wang, Y.; Guo, Y.; Xing, E.; Zhao, X.; Li, W.; Zhang, J.; Yu, C. Effect of diosgenin on T-helper 17 cells in mice with collagen-induced arthritis. Pharmacogn. Mag. 2020, 16, 486. [Google Scholar] [CrossRef]
- Wu, S.; Zhao, M.; Sun, Y.; Xie, M.; Le, K.; Xu, M.; Huang, C. The potential of Diosgenin in treating psoriasis: Studies from HaCaT keratinocytes and imiquimod-induced murine model. Life Sci. 2020, 241, 117115. [Google Scholar] [CrossRef]
- Ladurner, A.; Schwarz, P.F.; Dirsch, V.M. Natural products as modulators of retinoic acid receptor-related orphan receptors (RORs). Nat. Prod. Rep. 2021, 38, 757–781. [Google Scholar] [CrossRef]
- Karas, K.; Salkowska, A.; Karwaciak, I.; Walczak-Drzewiecka, A.; Dastych, J.; Bachorz, R.A.; Ratajewski, M. The Dichotomous Nature of AZ5104 (an EGFR Inhibitor) Towards RORgamma and RORgammaT. Int. J. Mol. Sci. 2019, 20, 5780. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [PubMed]
- GOLD, version v2020.2.0; Cambridge Crystallographic Data Centre (CCDC): Cambridge, UK, 2020.
- Maestro, version 2021-3; Schrödinger Inc.: New York, NY, USA, 2021.
- Roos, K.; Wu, C.; Damm, W.; Reboul, M.; Stevenson, J.M.; Lu, C.; Dahlgren, M.K.; Mondal, S.; Chen, W.; Wang, L.; et al. OPLS3e: Extending Force Field Coverage for Drug-Like Small Molecules. J. Chem. Theory Comput. 2019, 15, 1863–1874. [Google Scholar] [CrossRef]
- Kumar, N.; Solt, L.A.; Conkright, J.J.; Wang, Y.; Istrate, M.A.; Busby, S.A.; Garcia-Ordonez, R.D.; Burris, T.P.; Griffin, P.R. The benzenesulfoamide T0901317 [N-(2,2,2-trifluoroethyl)-N-[4-[2,2,2-trifluoro-1-hydroxy-1-(trifluoromethyl)ethy l]phenyl]-benzenesulfonamide] is a novel retinoic acid receptor-related orphan receptor-alpha/gamma inverse agonist. Mol. Pharmacol. 2010, 77, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Hiebl, V.; Ladurner, A.; Latkolik, S.; Dirsch, V.M. Natural products as modulators of the nuclear receptors and metabolic sensors LXR, FXR and RXR. Biotechnol. Adv. 2018, 36, 1657–1698. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Wang, X.; Zhong, B.; Nurieva, R.I.; Ding, S.; Dong, C. Ursolic acid suppresses interleukin-17 (IL-17) production by selectively antagonizing the function of RORgamma t protein. J. Biol. Chem. 2011, 286, 22707–22710. [Google Scholar] [CrossRef]
- Zhou, Q.; Qin, S.; Zhang, J.; Zhon, L.; Pen, Z.; Xing, T. 1,25(OH)2D3 induces regulatory T cell differentiation by influencing the VDR/PLC-gamma1/TGF-beta1/pathway. Mol. Immunol. 2017, 91, 156–164. [Google Scholar] [CrossRef]
- Li, F.; Fernandez, P.P.; Rajendran, P.; Hui, K.M.; Sethi, G. Diosgenin, a steroidal saponin, inhibits STAT3 signaling pathway leading to suppression of proliferation and chemosensitization of human hepatocellular carcinoma cells. Cancer Lett. 2010, 292, 197–207. [Google Scholar] [CrossRef]
- Wang, R.; Campbell, S.; Amir, M.; Mosure, S.A.; Bassette, M.A.; Eliason, A.; Sundrud, M.S.; Kamenecka, T.M.; Solt, L.A. Genetic and pharmacological inhibition of the nuclear receptor RORalpha regulates TH17 driven inflammatory disorders. Nat. Commun. 2021, 12, 76. [Google Scholar] [CrossRef]
- Manda, V.K.; Avula, B.; Ali, Z.; Wong, Y.H.; Smillie, T.J.; Khan, I.A.; Khan, S.I. Characterization of in vitro ADME properties of diosgenin and dioscin from Dioscorea villosa. Planta Med. 2013, 79, 1421–1428. [Google Scholar] [CrossRef] [PubMed]
- McAnuff, M.; Omoruyi, F.; Morrison, E.; Asemota, H. Changes in some liver enzymes in streptozotocin-induced diabetic rats fed sapogenin extract from bitter yam (Dioscorea polygonoides) or commercial diosgenin. West. Indian Med. J. 2005, 54, 97–101. [Google Scholar] [CrossRef]
- Li, R.; Liu, Y.; Shi, J.; Yu, Y.; Lu, H.; Yu, L.; Liu, Y.; Zhang, F. Diosgenin regulates cholesterol metabolism in hypercholesterolemic rats by inhibiting NPC1L1 and enhancing ABCG5 and ABCG8. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 1124–1133. [Google Scholar] [CrossRef] [PubMed]
- Binesh, A.; Devaraj, S.N.; Halagowder, D. Atherogenic diet induced lipid accumulation induced NFkappaB level in heart, liver and brain of Wistar rat and diosgenin as an anti-inflammatory agent. Life Sci. 2018, 196, 28–37. [Google Scholar] [CrossRef]
- Malisetty, V.; Patlolla, J.; Raju, J.; Marcus, L.; Choi, C.; Rao, C. Chemoprevention of colon cancer by diosgenin, a steroidal saponin constituent of fenugreek. Proc. Amer. Assoc. Cancer Res. 2005, 46, 2473. [Google Scholar]
- Raju, J.; Rao, C.V. Diosgenin, a steroid saponin constituent of yams and fenugreek: Emerging evidence for applications in medicine. Bioact. Compd. Phytomed. 2012, 125, 143. [Google Scholar]
- Wu, F.C.; Jiang, J.G. Effects of diosgenin and its derivatives on atherosclerosis. Food Funct. 2019, 10, 7022–7036. [Google Scholar] [CrossRef]
- Cayen, M.N.; Ferdinandi, E.S.; Greselin, E.; Dvornik, D. Studies on the disposition of diosgenin in rats, dogs, monkeys and man. Atherosclerosis 1979, 33, 71–87. [Google Scholar] [CrossRef]
- Okawara, M.; Tokudome, Y.; Todo, H.; Sugibayashi, K.; Hashimoto, F. Effect of beta-cyclodextrin derivatives on the diosgenin absorption in Caco-2 cell monolayer and rats. Biol. Pharm. Bull. 2014, 37, 54–59. [Google Scholar] [CrossRef]
- Liu, C.Z.; Chang, J.H.; Zhang, L.; Xue, H.F.; Liu, X.G.; Liu, P.; Fu, Q. Preparation and Evaluation of Diosgenin Nanocrystals to Improve Oral Bioavailability. AAPS PharmSciTech 2017, 18, 2067–2076. [Google Scholar] [CrossRef]
- Okawara, M.; Hashimoto, F.; Todo, H.; Sugibayashi, K.; Tokudome, Y. Effect of liquid crystals with cyclodextrin on the bioavailability of a poorly water-soluble compound, diosgenin, after its oral administration to rats. Int. J. Pharm. 2014, 472, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Parama, D.; Boruah, M.; Yachna, K.; Rana, V.; Banik, K.; Harsha, C.; Thakur, K.K.; Dutta, U.; Arya, A.; Mao, X.; et al. Diosgenin, a steroidal saponin, and its analogs: Effective therapies against different chronic diseases. Life Sci. 2020, 260, 118182. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward Primer | Reverse Primer |
|---|---|---|
| IL-17A | 5′-ACCGATCCACCTCACCTTGG-3′ | 5′-AGTCCACGTTCCCATCAGCG-3′ |
| G6PC [32] | 5′-TCCATACTGGTGGGTTTTGG-3′ | 5′-GAGGAAAATGAGCAGCAAGG-3′ |
| GAPDH | Hs_GAPDH_1_SG QuantiTect Primer Assay, GeneGlobe ID: QT00079247, Detected transcript: NM_001256799 | |
| beta actin | Hs_ACTB_1_SG QuantiTect Primer Assay, GeneGlobe ID: QT00095431, Detected transcript: NM_001101 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarz, P.F.; Perhal, A.F.; Schöberl, L.N.; Kraus, M.M.; Kirchmair, J.; Dirsch, V.M. Identification of the Natural Steroid Sapogenin Diosgenin as a Direct Dual-Specific RORα/γ Inverse Agonist. Biomedicines 2022, 10, 2076. https://doi.org/10.3390/biomedicines10092076
Schwarz PF, Perhal AF, Schöberl LN, Kraus MM, Kirchmair J, Dirsch VM. Identification of the Natural Steroid Sapogenin Diosgenin as a Direct Dual-Specific RORα/γ Inverse Agonist. Biomedicines. 2022; 10(9):2076. https://doi.org/10.3390/biomedicines10092076
Chicago/Turabian StyleSchwarz, Patrik F., Alexander F. Perhal, Lucia N. Schöberl, Martin M. Kraus, Johannes Kirchmair, and Verena M. Dirsch. 2022. "Identification of the Natural Steroid Sapogenin Diosgenin as a Direct Dual-Specific RORα/γ Inverse Agonist" Biomedicines 10, no. 9: 2076. https://doi.org/10.3390/biomedicines10092076