
Targeting Gut Microbiota as a Novel Strategy for Prevention and Treatment of Hypertension, Atrial Fibrillation and Heart Failure: Current Knowledge and Future Perspectives


Abstract
:1. Introduction
2. Arterial Hypertension
2.1. Gut Microbiota and Its Metabolites Contribute to Blood Pressure Regulation
2.1.1. Possible Links between Gut Microbiota Composition and Arterial Hypertension
2.1.2. Gut Microbiota Contributes to Salt-Sensitivity of Blood Pressure
2.2. Gut Microbiota Modulation Approaches in Arterial Hypertension Management: From Dietary Interventions to Washed Microbiota Transplantation
2.2.1. Dietary Interventions
2.2.2. Prebiotics and Probiotics Supplementation
2.2.3. Postbiotics (e.g., SCFAs)
2.2.4. Washed Microbiota Transplantation
3. Atrial Fibrillation
3.1. The Role of «Pro-Arrhythmic» Gut Microbiota and Its Metabolites in Atrial Fibrillation Development
3.1.1. Gut Microbiota Dysbiosis Contributes to Atrial Fibrillation Susceptibility through Inflammation
3.1.2. The Mechanistic Links between Gut Microbiota-Derived Metabolites and AF
3.2. Possible Clinical Interventions to Modify Gut Microbiota Composition and Metabolites in AF Management
3.2.1. Dietary Interventions
3.2.2. Targeting the Metabolic Pathway of TMAO
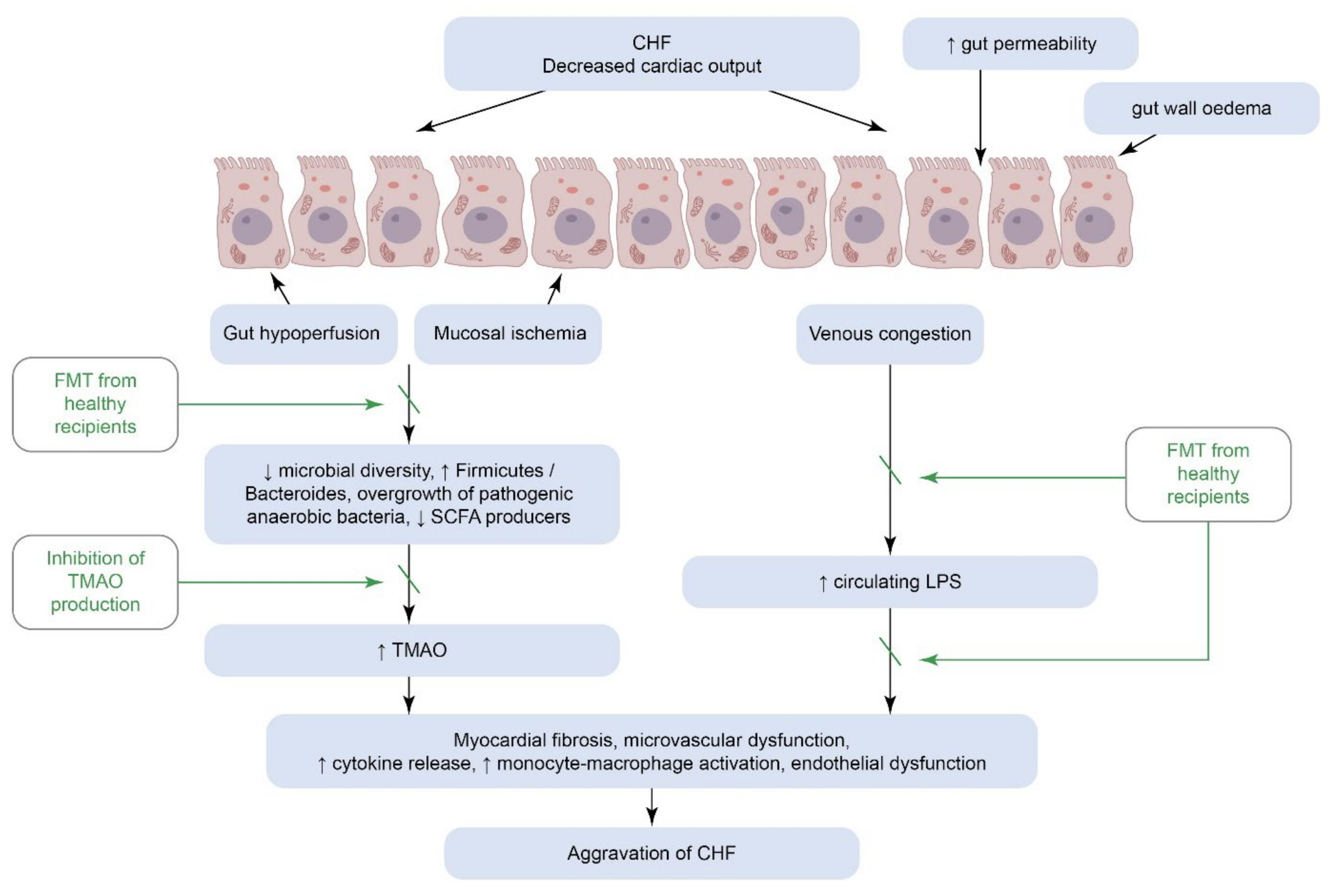
4. Congestive Heart Failure
4.1. Gut Microbiota as a Cornerstone in the Development of Congestive Heart Failure
4.1.1. “Gut Hypothesis” of Congestive Heart Failure
4.1.2. Gut Microbiota Implication in the Initial Development of Immune-Mediated Cardiomyopathies
4.2. Targeting the Gut Microbiome to Prevent and Treat Heart Failure
4.2.1. Dietary Interventions
4.2.2. Probiotics
4.2.3. Lowering TMAO Could Break the Vicious Circle
4.3. The GutHeart Trial Results
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global burden of cardiovascular diseases and risk factors, 1990–2019: Update from the GBD 2019 Study. J. Am. Coll. Card. 2020, 76, 298–3021. [Google Scholar] [CrossRef] [PubMed]
- Khot, U.N.; Khot, M.B.; Bajzer, C.T.; Sapp, S.K.; Ohman, E.M.; Brener, S.J.; Ellis, S.G.; Lincoff, A.M.; Topol, E.J. Prevalence of conventional risk factors in patients with coronary heart disease. JAMA 2003, 290, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.F.F.A.; Sampaio-Maia, B.; Araujo, R.; Nascimento, D.S.; Ferreira-Gomes, J.; Pestana, M.; Azevedo, M.J.; Alencastre, I.S. Gut microbiome and organ fibrosis. Nutrients 2022, 14, 352. [Google Scholar] [CrossRef] [PubMed]
- Mishima, E.; Abe, T. Role of the microbiota in hypertension and antihypertensive drug metabolism. Hypertens. Res. 2022, 45, 246–253. [Google Scholar] [CrossRef]
- Velasquez, M.T.; Ramezani, A.; Manal, A.; Raj, D.S. Trimethylamine N-Oxide: The Good, the Bad and the Unknown. Toxins 2016, 8, 326. [Google Scholar] [CrossRef]
- van den Munckhof, I.C.L.; Kurilshikov, A.; Ter Horst, R.; Riksen, N.P.; Joosten, L.A.B.; Zhernakova, A.; Fu, J.; Keating, S.T.; Netea, M.G.; de Graaf, J.; et al. Role of gut microbiota in chronic low-grade inflammation as potential driver for atherosclerotic cardiovascular disease: A systematic review of human studies. Obes. Rev. 2018, 19, 1719–1734. [Google Scholar] [CrossRef]
- Farnworth, E.R.; Mainville, I.; Desjardins, M.P.; Gardner, N.; Fliss, I.; Champagne, C. Growth of probiotic bacteria and bifidobacteria in a soy yogurt formulation. Int. J. Food Microbiol. 2007, 116, 174–181. [Google Scholar] [CrossRef]
- FAO/WHO. Evaluation of health and nutritional properties of powder milk and live lactic acid bacteria. In Report from FAO/WHO Expert Consultation, Córdoba, Argentina; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001; pp. 1–4. [Google Scholar]
- Bafeta, A.; Yavchitz, A.; Riveros, C.; Batista, R.; Ravaud, P. Methods and reporting studies assessing fecal microbiota transplantation: A systematic review. Ann. Intern. Med. 2017, 167, 34–39. [Google Scholar] [CrossRef]
- Yang, G.; Wei, J.; Liu, P.; Zhang, Q.; Tian, Y.; Hou, G.; Meng, L.; Xin, Y.; Jiang, X. Role of the gut microbiota in type 2 diabetes and related diseases. Metabolism 2021, 117, 154712. [Google Scholar] [CrossRef]
- Piccioni, A.; de Cunzo, T.; Valletta, F.; Covino, M.; Rinninella, E.; Raoul, P.; Zanza, C.; Mele, M.C.; Franceschi, F. Gut Microbiota and Environment in Coronary Artery Disease. Int. J. Environ. Res. Public Health 2021, 18, 4242. [Google Scholar] [CrossRef]
- Zhang, L.; Chu, J.; Hao, W.; Zhang, J.; Li, H.; Yang, C.; Yang, J.; Chen, X.; Wang, H. Gut Microbiota and Type 2 Diabetes Mellitus: Association, Mechanism, and Translational Applications. Mediat. Inflamm. 2021, 2021, 5110276. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhuang, J.; Tang, P.; Li, J.; Xiong, X.; Deng, H. The Role of the Gut Microbiota in Coronary Heart Disease. Curr. Atheroscler. Rep. 2020, 22, 77. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef]
- Palmu, J.; Salosensaari, A.; Havulinna, A.S.; Cheng, S.; Inouye, M.; Jain, M.; Salido, R.A.; Sanders, K.; Brennan, C.; Humphrey, G.C.; et al. Association between the gut microbiota and blood pressure in a population cohort of 6953 individuals. J. Am. Heart Assoc. 2020, 9, e016641. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Lulla, A.; Sioda, M.; Winglee, K.; Wu, M.C.; Jacobs, D.R., Jr.; Shikany, J.M.; Lloyd-Jones, D.M.; Launer, L.J.; Fodor, A.A.; et al. Gut microbiota composition and blood pressure. Hypertension 2019, 73, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Verhaar, B.J.H.; Collard, D.; Prodan, A.; Levels, J.H.M.; Zwinderman, A.H.; Bäckhed, F.; Vogt, L.; Peters, M.J.L.; Muller, M.; Nieuwdorp, M.; et al. Associations between gut microbiota, faecal short-chain fatty acids, and blood pressure across ethnic groups: The HELIUS study. Eur. Heart J. 2020, 41, 4259–4267. [Google Scholar] [CrossRef]
- Karbach, S.H.; Schönfelder, T.; Brandão, I.; Wilms, E.; Hörmann, N.; Jäckel, S.; Schüler, R.; Finger, S.; Knorr, M.; Lagrange, J.; et al. Gut Microbiota Promote Angiotensin II-Induced Arterial Hypertension and Vascular Dysfunction. J. Am. Heart Assoc. 2016, 5, e003698. [Google Scholar] [CrossRef]
- Jiang, S.; Shui, Y.; Cui, Y.; Tang, C.; Wang, X.; Qiu, X.; Hu, W.; Fei, L.; Li, Y.; Zhang, S.; et al. Gut microbiota dependent trimethylamine N-oxide aggravates angiotensin II-induced hypertension. Redox Biol. 2021, 46, 102115. [Google Scholar] [CrossRef]
- Toral, M.; Robles-Vera, I.; de la Visitación, N.; Romero, M.; Sánchez, M.; Gómez-Guzmán, M.; Rodriguez-Nogales, A.; Yang, T.; Jiménez, R.; Algieri, F.; et al. Role of the immune system in vascular function and blood pressure control induced by faecal microbiota transplantation in rats. Acta Physiol. 2019, 227, e13285. [Google Scholar] [CrossRef]
- Elijovich, F.; Laffer, C.L. Why is salt-sensitivity of blood pressure, a known cardiovascular risk factor, not treated? Int. J. Cardiol. Hypertens. 2021, 9, 100096. [Google Scholar] [CrossRef]
- Marques, F.Z.; Mackay, C.R.; Kaye, D.M. Beyond gut feelings: How the gut microbiota regulates blood pressure. Nat. Rev. Cardiol. 2018, 15, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Linz, B.; Hohl, M.; Reil, J.C.; Böhm, M.; Linz, D. Inhibition of NHE3-mediated sodium absorption in the gut reduced cardiac end-organ damage without deteriorating renal function in obese spontaneously hypertensive rats. J. Cardiovasc. Pharmacol. 2016, 67, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Bier, A.; Braun, T.; Khasbab, R.; Di Segni, A.; Grossman, E.; Haberman, Y.; Leibowitz, A. A high salt diet modulates the gut microbiota and short chain fatty acids production in a salt-sensitive hypertension rat model. Nutrients 2018, 10, 1154. [Google Scholar] [CrossRef] [PubMed]
- Seck, E.H.; Senghor, B.; Merhej, V.; Bachar, D.; Cadoret, F.; Robert, C.; Azhar, E.I.; Yasir, M.; Bibi, F.; Jiman-Fatani, A.A.; et al. Salt in stools is associated with obesity, gut halophilic microbiota and Akkermansiamuciniphila depletion in humans. Int. J. Obes. 2019, 43, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.F.; Aden, L.A.; Barbaro, N.R.; Van Beusecum, J.P.; Xiao, L.; Simmons, A.J.; Warden, C.; Pasic, L.; Himmel, L.E.; Washington, M.K.; et al. High dietary salt-induced dendritic cell activation underlies microbial dysbiosis-associated hypertension. JCI Insight 2019, 5, e126241. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, F.J.; Dong, Y.; Huang, Y.; Wang, C.; Harshfield, G.A.; Zhu, H. Modest sodium reduction increases circulating short-chain fatty acids in untreated hypertensives: A randomized, double-blind, placebo-controlled trial. Hypertension 2020, 76, 73–79. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Chaimani, A.; Schwedhelm, C.; Toledo, E.; Pünsch, M.; Hoffmann, G.; Boeing, H. Comparative effects of different dietary approaches on blood pressure in hypertensive and pre-hypertensive patients: A systematic review and network meta-analysis. Crit. Rev. Food Sci. Nutr. 2019, 59, 2674–2687. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, B.; Abo-Hamzy, T.; Nelson, J.W.; Ambati, C.S.R.; Petrosino, J.F.; Bryan, R.M., Jr.; Durgan, D.J. Restructuring the gut microbiota by intermittent fasting lowers blood pressure. Circ. Res. 2021, 128, 1240–1254. [Google Scholar] [CrossRef]
- Wilhelmi de Toledo, F.; Grundler, F.; Bergouignan, A.; Drinda, S.; Michalsen, A. Safety, health improvement and well-being during a 4 to 21-day fasting period in an observational study including 1422 subjects. PLoS ONE 2019, 14, e0209353. [Google Scholar] [CrossRef]
- Maifeld, A.; Bartolomaeus, H.; Löber, U.; Avery, E.G.; Steckhan, N.; Markó, L.; Wilck, N.; Hamad, I.; Šušnjar, U.; Mähler, A.; et al. Fasting alters the gut microbiome reducing blood pressure and body weight in metabolic syndrome patients. Nat. Commun. 2021, 12, 1970. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Merra, G.; Noce, A.; Marrone, G.; Cintoni, M.; Tarsitano, M.G.; Capacci, A.; De Lorenzo, A. Influence of Mediterranean Diet on Human Gut Microbiota. Nutrients 2020, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on Gut Microbiota Associated to Mediterranean Diet Adherence and Specific Dietary Intakes on General Adult Population. Front. Microbiol. 2018, 9, 890. [Google Scholar] [CrossRef] [PubMed]
- Ivey, K.L.; Hodgson, J.M.; Kerr, D.A.; Thompson, P.L.; Stojceski, B.; Prince, R.L. The effect of yoghurt and its probiotics on blood pressure and serum lipid profile; a randomised controlled trial. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Hariri, M.; Salehi, R.; Feizi, A.; Mirlohi, M.; Kamali, S.; Ghiasvand, R. The effect of probiotic soy milk and soy milk on anthropometric measures and blood pressure in patients with type II diabetes mellitus: A randomized double-blind clinical trial. ARYA Atheroscler. 2015, 11 (Suppl. S1), 74–80. [Google Scholar] [PubMed]
- Marques, F.Z.; Nelson, E.; Chu, P.Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef]
- Khan, K.; Jovanovski, E.; Ho, H.V.T.; Marques, A.C.R.; Zurbau, A.; Mejia, S.B.; Sievenpiper, J.L.; Vuksan, V. The effect of viscous soluble fiber on blood pressure: A systematic review and meta-analysis of randomized controlled trials. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 3–13. [Google Scholar] [CrossRef]
- Faghihimani, Z.; Namazi, N.; Ghaffari, S.; Kelishadi, M.R.; Sharifi, S.; Nattagh-Eshtivani, E.; Akbarzadeh, M.; Moravejolahkami, A.R.; Khorvash, F.; Roshanravan, N.; et al. Effects of Inulin Type-Carbohydrates on blood pressure: A systematic review and meta-analysis. Int. J. Food Prop. 2021, 24, 129–139. [Google Scholar] [CrossRef]
- Qi, D.; Nie, X.L.; Zhang, J.J. The effect of probiotics supplementation on blood pressure: A systemic review and meta-analysis. Lipids Health Dis. 2020, 19, 79. [Google Scholar] [CrossRef]
- Cornelissen, V.A.; Fagard, R.H. Effect of resistance training on resting blood pressure: A meta-analysis of randomized controlled trials. J. Hypertens. 2005, 23, 251–259. [Google Scholar] [CrossRef]
- Aoyagi, Y.; Park, S.; Matsubara, S.; Honda, Y.; Amamoto, R.; Kushiro, A.; Miyazaki, K.; Shephard, R.J. Habitual intake of fermented milk products containing Lactobacillus casei strain Shirota and a reduced risk of hypertension in older people. Benef. Microbes 2017, 8, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Khalesi, S.; Sun, J.; Buys, N.; Jayasinghe, R. Effect of probiotics on blood pressure: A systematic review and meta-analysis of randomized, controlled trials. Hypertension 2014, 64, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Durgan, D.J.; Ganesh, B.P.; Cope, J.L.; Ajami, N.J.; Phillips, S.C.; Petrosino, J.F.; Hollister, E.B.; Bryan, R.M., Jr. Role of the Gut Microbiome in Obstructive Sleep Apnea-Induced Hypertension. Hypertension 2016, 67, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, N.; Hori, D.; Flavahan, S.; Steppan, J.; Flavahan, N.A.; Berkowitz, D.E.; Pluznick, J.L. Microbial short chain fatty acid metabolites lower blood pressure via endothelial G protein-coupled receptor 41. Physiol. Genomics 2016, 48, 826–834. [Google Scholar] [CrossRef]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-Chain Fatty Acid Propionate Protects From Hypertensive Cardiovascular Damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef]
- Kim, S.; Goel, R.; Kumar, A.; Qi, Y.; Lobaton, G.; Hosaka, K.; Mohammed, M.; Handberg, E.M.; Richards, E.M.; Pepine, C.J.; et al. Imbalance of gut microbiome and intestinal epithelial barrier dysfunction in patients with high blood pressure. Clin. Sci. 2018, 132, 701–718. [Google Scholar] [CrossRef]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.X.; Rey, F.; Wang, T.; et al. Olfactory receptor responding to gut microbiota–derived signals plays a role in renin secretion and blood pressure regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef]
- Kaye, D.M.; Shihata, W.A.; Jama, H.A.; Tsyganov, K.; Ziemann, M.; Kiriazis, H.; Horlock, D.; Vijay, A.; Giam, B.; Vinh, A.; et al. Deficiency of Prebiotic Fiber and Insufficient Signaling Through Gut Metabolite-Sensing Receptors Leads to Cardiovascular Disease. Circulation 2020, 141, 1393–1403. [Google Scholar] [CrossRef]
- Shi, Q. Nanjing consensus on methodology of washed microbiota transplantation. Chin. Med. J. 2020, 133, 2330–2332. [Google Scholar] [CrossRef]
- Zhong, H.J.; Zeng, H.L.; Cai, Y.L.; Zhuang, Y.P.; Liou, Y.L.; Wu, Q.; He, X.X. Washed Microbiota Transplantation Lowers Blood Pressure in Patients with Hypertension. Front. Cell. Infect. Microbiol. 2021, 11, 679624. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Li, B.; Luo, Y.; Gong, Y.; Jin, X.; Zhang, J.; Zhou, Y.; Zhuo, X.; Wang, Z.; et al. Gut microbiota dysbiosis promotes age-related atrial fibrillation by lipopolysaccharide and glucose-induced activation of NLRP3-inflammasome. Cardiovasc. Res. 2022, 118, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Chelu, M.G.; Dobrev, D.; Li, N. Cardiomyocyte Inflammasome Signaling in Cardiomyopathies and Atrial Fibrillation: Mechanisms and Potential Therapeutic Implications. Front. Physiol. 2018, 9, 1115. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Meng, G.; Huang, B.; Zhou, X.; Stavrakis, S.; Wang, M.; Li, X.; Zhou, L.; Wang, Y.; Wang, M.; et al. A potential relationship between gut microbes and atrial fibrillation: Trimethylamine N-oxide, a gut microbe-derived metabolite, facilitates the progression of atrial fibrillation. Int. J. Cardiol. 2018, 255, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Zhang, S.; Zhu, J.; Jiang, H.; Jia, D.; Ou, T.; Qi, Z.; Zou, Y.; Qian, J.; Sun, A.; et al. Gut microbe-derived metabolite trimethylamine N-oxide accelerates fibroblast-myofibroblast differentiation and induces cardiac fibrosis. J. Mol. Cell. Cardiol. 2019, 134, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Gawałko, M.; Agbaedeng, T.A.; Saljic, A.; Müller, D.N.; Wilck, N.; Schnabel, R.; Penders, J.; Rienstra, M.; van Gelder, I.; Jespersen, T.; et al. Gut microbiota, dysbiosis and atrial fibrillation. Arrhythmogenic mechanisms and potential clinical implications. Cardiovasc. Res. 2021, cvab292. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Zhang, L.; Zhang, Y.; Wang, F.; Zhao, Z.; Zhou, X. Gut Microbial Metabolite Trimethylamine N-Oxide Is Related to Thrombus Formation in Atrial Fibrillation Patients. Am. J. Med. Sci. 2019, 358, 422–428. [Google Scholar] [CrossRef]
- Melgaard, L.; Gorst-Rasmussen, A.; Lane, D.A.; Rasmussen, L.H.; Larsen, T.B.; Lip, G.Y. Assessment of the CHA2DS2-VASc Score in Predicting Ischemic Stroke, Thromboembolism, and Death in Patients With Heart Failure With and Without Atrial Fibrillation. JAMA 2015, 314, 1030–1038. [Google Scholar] [CrossRef]
- Chen, W.T.; Chen, Y.C.; Hsieh, M.H.; Huang, S.Y.; Kao, Y.H.; Chen, Y.A.; Lin, Y.K.; Chen, S.A.; Chen, Y.J. The uremic toxin indoxyl sulfate increases pulmonary vein and atrial arrhythmogenesis. J. Cardiovasc. Electrophysiol. 2015, 26, 203–210. [Google Scholar] [CrossRef]
- Yamagami, F.; Tajiri, K.; Yumino, D.; Ieda, M. Uremic Toxins and Atrial Fibrillation: Mechanisms and Therapeutic Implications. Toxins 2019, 11, 597. [Google Scholar] [CrossRef]
- Aoki, K.; Teshima, Y.; Kondo, H.; Saito, S.; Fukui, A.; Fukunaga, N.; Nawata, T.; Shimada, T.; Takahashi, N.; Shibata, H. Role of Indoxyl Sulfate as a Predisposing Factor for Atrial Fibrillation in Renal Dysfunction. J. Am. Heart Assoc. 2015, 4, e002023. [Google Scholar] [CrossRef]
- Asgharzadeh, F.; Bargi, R.; Hosseini, M.; Farzadnia, M.; Khazaei, M. Cardiac and renal fibrosis and oxidative stress balance in lipopolysaccharide-induced inflammation in male rats. ARYA Atheroscler. 2018, 14, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Pastori, D.; Carnevale, R.; Nocella, C.; Novo, M.; Santulli, M.; Cammisotto, V.; Menichelli, D.; Pignatelli, P.; Violi, F. Gut-Derived Serum Lipopolysaccharide is Associated with Enhanced Risk of Major Adverse Cardiovascular Events in Atrial Fibrillation: Effect of Adherence to Mediterranean Diet. J. Am. Heart Assoc. 2017, 6, e005784. [Google Scholar] [CrossRef]
- Pastori, D.; Carnevale, R.; Menichelli, D.; Nocella, C.; Bartimoccia, S.; Novo, M.; Leo, I.; Violi, F.; Pignatelli, P. Is There an Interplay Between Adherence to Mediterranean Diet, Antioxidant Status, and Vascular Disease in Atrial Fibrillation Patients? Antioxid. Redox Signal 2016, 25, 751–755. [Google Scholar] [CrossRef]
- Mattioli, A.V.; Pennella, S.; Pedrazzi, P.; Farinetti, A. Gender Differences in Adherence to Mediterranean Diet and Risk of Atrial Fibrillation. JHC 2015, 1, 4–13. [Google Scholar] [CrossRef]
- Wang, G.; Kong, B.; Shuai, W.; Fu, H.; Jiang, X.; Huang, H. 3,3-Dimethyl-1-butanol attenuates cardiac remodeling in pressure-overload-induced heart failure mice. J. Nutr. Biochem. 2020, 78, 108341. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zheng, X.; Feng, M.; Li, D.; Zhang, H. Gut Microbiota-Dependent Metabolite Trimethylamine N-Oxide Contributes to Cardiac Dysfunction in Western Diet-Induced Obese Mice. Front. Physiol. 2017, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.W.; Li, D.Y.; Hazen, S.L. Dietary metabolism, the gut microbiome, and heart failure. Nat. Rev. Cardiol. 2019, 16, 137–154. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-F.; Zhang, W.-Y.; Wen, Q.; Chen, W.J.; Wang, Z.M.; Chen, J.; Zhu, F.; Liu, K.; Cheng, L.X.; Yang, J.; et al. Fecal Microbiota Transplantation Alleviates Myocardial Damage in Myocarditis by Restoring the Microbiota Composition. Pharmacol. Res. 2019, 139, 412–421. [Google Scholar] [CrossRef]
- Gil-Cruz, C.; Perez-Shibayama, C.; De Martin, A.; Ronchi, F.; van der Borght, K.; Niederer, R.; Onder, L.; Lütge, M.; Novkovic, M.; Nindl, V. Microbiota-derived peptide mimics drive lethal inflammatory cardiomyopathy. Science 2019, 366, 881–886. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Q.; Qi, H.; Wang, C.; Wang, C.; Zhang, J.; Dong, L. Doxorubicin-induced systemic inflammation is driven by upregulation of toll-like receptor TLR4 and endotoxin leakage. Cancer Res. 2016, 76, 6631–6642. [Google Scholar] [CrossRef]
- An, L.; Wuri, J.; Zheng, Z.; Li, W.; Yan, T. Microbiota modulate Doxorubicin induced cardiotoxicity. Eur. J. Pharm. Sci. 2021, 166, 105977. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.J.; Ordóñez-Mena, J.M.; Roalfe, A.K.; Lay-Flurrie, S.; Jones, N.R.; Marshall, T.; Hobbs, F.D.R. Trends in survival after a diagnosis of heart failure in the United Kingdom 2000-2017: Population based cohort study. BMJ 2019, 364, l223. [Google Scholar] [CrossRef] [PubMed]
- Chrysohoou, C.; Metallinos, G.; Aggelopoulos, P.; Kastorini, C.; Athanasopoulou, S.; Pitsavos, C.; Panagiotakos, D.B.; Stefanadis, C. Long-term adherence to the traditional Mediterranean diet is associated with improved biventricular systolic function, in chronic heart failure patients. Eur. J. Heart Fail. 2009, 120, 739. [Google Scholar]
- Tektonidis, T.G.; Åkesson, A.; Gigante, B.; Wolk, A.; Larsson, S.C. A Mediterranean diet and risk of myocardial infarction, heart failure and stroke: A population-based cohort study. Atherosclerosis 2015, 243, 93–98. [Google Scholar] [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef]
- Liyanage, T.; Ninomiya, T.; Wang, A.; Neal, B.; Jun, M.; Wong, M.G.; Jardine, M.; Hillis, G.S.; Perkovic, V. Effects of the Mediterranean Diet on Cardiovascular Outcomes-A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0159252. [Google Scholar] [CrossRef]
- Levitan, E.B.; Lewis, C.E.; Tinker, L.F.; Eaton, C.B.; Ahmed, A.; Manson, J.E.; Snetselaar, L.G.; Martin, L.W.; Trevisan, M.; Howard, B.V.; et al. Mediterranean and DASH diet scores and mortality in women with heart failure: The Women’s Health Initiative. Circ. Heart Fail. 2013, 6, 1116–1123. [Google Scholar] [CrossRef]
- Gan, X.T.; Ettinger, G.; Huang, C.X.; Burton, J.P.; Haist, J.V.; Rajapurohitam, V.; Sidaway, J.E.; Martin, G.; Gloor, G.B.; Swann, J.R.; et al. Probiotic administration attenuates myocardial hypertrophy and heart failure after myocardial infarction in the rat. Circ. Heart Fail. 2014, 7, 491–499. [Google Scholar] [CrossRef]
- Costanza, A.C.; Moscavitch, S.D.; FariaNeto, H.C.; Mesquita, E.T. Probiotic therapy with Saccharomyces boulardii for heart failure patients: A randomized, double-blind, placebo-controlled pilot trial. Int. J. Cardiol. 2015, 179, 348–350. [Google Scholar] [CrossRef]
- Organ, C.L.; Li, Z.; Sharp, T.E., III; Polhemus, D.J.; Gupta, N.; Goodchild, T.T.; Tang, W.H.W.; Hazen, S.L.; Lefer, D.J. Nonlethal Inhibition of Gut Microbial Trimethylamine N-oxide Production Improves Cardiac Function and Remodeling in a Murine Model of Heart Failure. J. Am. Heart Assoc. 2020, 9, e016223. [Google Scholar] [CrossRef]
- Awoyemi, A.; Mayerhofer, C.; Felix, A.S.; Hov, J.R.; Moscavitch, S.D.; Lappegård, K.T.; Hovland, A.; Halvorsen, S.; Halvorsen, B.; Gregersen, I.; et al. Rifaximin or Saccharomyces boulardii in heart failure with reduced ejection fraction: Results from the randomized GutHeart trial. EBioMedicine 2021, 70, 103511. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.; Aquilani, R.; Testa, C.; Baiardi, P.; Angioletti, S.; Boschi, F.; Verri, M.; Dioguardi, F. Pathogenic gut flora in patients with chronic heart failure. JACC Heart Fail. 2016, 4, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Yamashita, T.; Watanabe, H.; Kami, K.; Yoshida, N.; Tabata, T.; Emoto, T.; Sasaki, N.; Mizoguchi, T.; Irino, Y.; et al. Gut Microbiome and Plasma Microbiome-Related Metabolites in Patients with Decompensated and Compensated Heart Failure. Circ. J. 2018, 83, 182–192. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Patients/Model | Intervention | Main Findings | Ref |
|---|---|---|---|
| Rat model | Every other day fasting (EODF); cholic acid supplementation | EODF ↓ systolic BP, significantly altered the community structure of GMB and ↑ BA metabolism genes | [29] |
| Murine model | High-fiber diet, acetate supplementation | ↓ Firmicutes/Bacteroidetes ratio, ↑ the abundance of Bacteroides acidifaciens; ↓ systolic and diastolic BPs, cardiac fibrosis, and left ventricular hypertrophy; acetate also markedly ↓ renal fibrosis. | [37] |
| Murine model | Propionate intake in drinking water | Attenuation of cardiac hypertrophy, fibrosis, vascular dysfunction, ↓ susceptibility to ventricular arrhythmias; ↓ aortic atherosclerotic lesion area. | [46] |
| Murine model | Butyrate administration | ↓ mean arterial pressure; ↓ spontaneous cardiac baroreceptor reflex gain and ↑ cardiac sympathetic tone in Ang II-treated mice; Improvement of gut barrier function, acceleration of mucus turnover and amelioration of gut inflammation. | [47] |
| Murine model | Diet lacking prebiotic fiber with or without addition SCFAs | In the absence of dietary fiber, SCFAs supplementation protected against the development of HTN, myocardial hypertrophy and fibrosis. | [49] |
| 145 untreated hypertensive patients | Sodium restriction to 2000 mg/day | ↑ of circulating SCFAs; ↓ BP and improved arterial compliance | [27] |
| 1422 subjects | Intermittent fasting | The mean values for the whole cohort ↓ dramatically (from 131.6 ± 0.7 to 120.7 ± 0.4 for systolic BP and from 83.7 ± 0.4 to 77.9 ± 0.3 for diastolic BP (fasting intervention: p < 0.001 for both); the effect size was directly related to fasting duration (p < 0.001) without gender difference | [30] |
| Patients with metabolic syndrome | 5-day fasting + DASH-diet/DASH diet alone | Fasting + DASH diet substantially ↓ 24 h ambulatory systolic BP and mean arterial pressure 3 months post-intervention as opposed to DASH diet alone. Participants who were prescribed fasting + DASH diet have been shown to ↓ their intake of antihypertensive medication by 3 months post-intervention in comparison to counterparts who followed a DASH diet only. | [31] |
| 5 studies involving 233 participants | Inulin-type carbohydrate supplementation | ↓ systolic BP in women (WMD: −12.19 mmHg; 95% CI: −17.25, −7.13, p = 0.012). | [39] |
| 23 RCTs involving 2037 participants | Probiotic consumption | Probiotics ↓ systolic BP only in HTN [WMD = −3.31 mmHg, 95% CI: −5.71, −0.92; p = 0.007] and type 2 diabetes mellitis (WMD = −4.85 mmHg, 95% CI: −9.28, −0.42; p = 0.032); Probiotics ↓ diastolic BP only in HTN (WMD = −2.02 mmHg, 95% CI: −3.68, −0.36; p = 0.017). BP-lowering effect could be observed for a short-term time 8–10 weeks; | [40] |
| 352 community-living initially normotensive subjects | Fermented milk products containing Lactobacillus casei strain Shirota | BP was more likely to remain in normal range over 5 years in participants who took ≥3 fermented milk products rather than <3 times/week (relative risk 0.398 [95% confidence interval 0.167–0.948], p = 0.037). | [42] |
| 260 patients (187 normotensive patients and 73 patients with HTN) | WMT | ↓ systolic BP: −5.09 ± 15.51, (p = 0.009); ↓ diastolic BP: −7.74 ± 10.42, (p < 0.001). WMT via the lower gastrointestinal tract and those not taking antihypertensive drugs had a greater decrease in systolic BP, and hypertensive patients not taking antihypertensive drugs also had a greater decrease in diastolic BP. WMT increased the Shannon Diversity Index in 6 of 8 hypertensive patients. | [51] |
| Patients/Model | Intervention | Main Findings | Ref |
|---|---|---|---|
| Rat model | FMT | FMT from aged AF-rats to young rats ↑ AF susceptibility, atrial fibrosis, ↑ inducibility and longer AF-duration; FMT from aged to young rats resulted in gut barrier dysfunction; Transferring GMB from young to aged rats ↓ the activation of atrial NLRP3-inflammasome and ↓ atrial fibrosis; also, the aged rats that were re-colonized with young microbiota failed in inducing AF by burst electrical pacing. | [52] |
| Rat model | AST-120 | ↓ renal dysfunction-induced oxidative stress, inflammation, and atrial fibrosis; ↓AF inducibility. | [61] |
| Pressure-overload-induced HF mice | Normal diet and given water supplemented with or without 1.0% DMB for 6 weeks | ↓ TMAO levels in overload-induced HF mice, ↓ adverse cardiac structural and electrical remodeling. | [67] |
| 690 AF-patients | Mediterranean diet | ↑ antioxidant activity of GPx3 in AF, ↓ vascular events rate. | [65] |
| Patients/Model | Intervention | Main Findings | Ref |
|---|---|---|---|
| EAM mouse | FMT | ↓ myocardial damage, ↓ inflammatory infiltration with cardiomyocyte disarray and necrosis in the cardiac tissues. ↑ increasing the Bacteroides population, ↓ Firmicutes/Bacteroidetes ratio and reshaping the GMB composition; | [71] |
| DOX-treated mice | FMT | Improvement of cardiac function, ↓ of DOX- induced GMB dysbiosis,↑ colorectum length, ↓ goblet cell loss, number of intestinal ulcers and lymphocyte cluster infiltration, loss of tight junction protein, and ↓ plasma LPS level. | [73] |
| Murine model | Normal diet or a Western diet (WD), without or with 1.0% DMB in drinking water | ↓ cardiac dysfunction and fibrosis in mice fed a WD by inhibition WD-induced increase in serum TMAO level. | [68] |
| Rat model | Administration of the probiotic Lactobacillus rhamnosus GR-1 or placebo in the drinking water | Significant attenuation of left ventricular hypertrophy, improvement of hemodynamic parameters such as preservation of left ventricular ejection fraction and fractional shortening. | [80] |
| 218 HF-patients | Mediterranean diet | Improvement of biventricular systolic function. | [75] |
| 32,921 women | Mediterranean diet | A high adherence to the MedD ↓ risk of myocardial infarction (RR: 0.74, 95% CI: 0.61–0.90, p = 0.003), HF (RR: 0.79, 95% CI: 0.68–0.93, p = 0.004) and ischemic stroke (RR: 0.78, 95% CI: 0.65–0.93, p = 0.007) | [76] |
| 3215 women experienced a HF hospitalization | Mediterranean and Dietary Approaches to StopHypertension (DASH) diet | Higher adherence to the Mediterranean diet was associated with a ↓ hazard rate of death among women with HF; Women in the top quartile of the DASH diet score had a 16% ↓ hazard rate of death than those in the bottom quartile (p for linear trend = 0.01). | [79] |
| 150 patients with stable HFrEF | Rifaximin or Saccharomyces boulardii | No significant effect on LVEF, microbiota diversity, plasma C-reactive protein and TMAO levels. | [84] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drapkina, O.M.; Yafarova, A.A.; Kaburova, A.N.; Kiselev, A.R. Targeting Gut Microbiota as a Novel Strategy for Prevention and Treatment of Hypertension, Atrial Fibrillation and Heart Failure: Current Knowledge and Future Perspectives. Biomedicines 2022, 10, 2019. https://doi.org/10.3390/biomedicines10082019
Drapkina OM, Yafarova AA, Kaburova AN, Kiselev AR. Targeting Gut Microbiota as a Novel Strategy for Prevention and Treatment of Hypertension, Atrial Fibrillation and Heart Failure: Current Knowledge and Future Perspectives. Biomedicines. 2022; 10(8):2019. https://doi.org/10.3390/biomedicines10082019
Chicago/Turabian StyleDrapkina, Oxana M., Adel A. Yafarova, Anastasia N. Kaburova, and Anton R. Kiselev. 2022. "Targeting Gut Microbiota as a Novel Strategy for Prevention and Treatment of Hypertension, Atrial Fibrillation and Heart Failure: Current Knowledge and Future Perspectives" Biomedicines 10, no. 8: 2019. https://doi.org/10.3390/biomedicines10082019