
CARD14 Signalling Ensures Cell Survival and Cancer Associated Gene Expression in Prostate Cancer Cells

 , , and
, , and 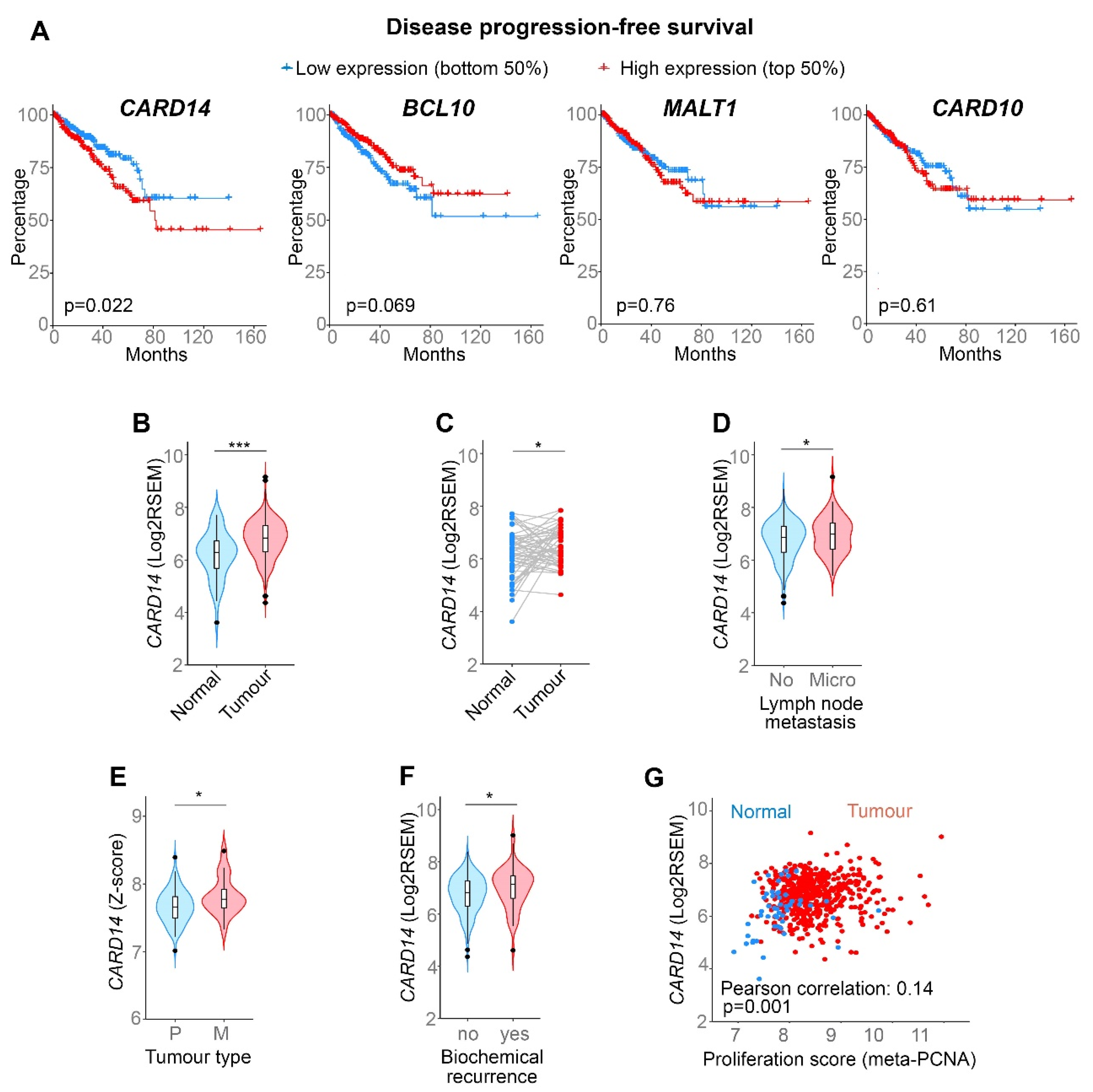{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Human PCa Data Sets
2.3. Proliferation Score
2.4. Gene Ontology (GO) Enrichment Analysis
2.5. Gene Set Enrichment Analysis (GSEA)
2.6. DNA Damage Response (DDR) Pathway Activity
2.7. Cells, siRNA-Mediated Knockdown and Cell Death Assays
2.8. PC3 Genome Editing
2.9. Multiplex Antibody Array Analysis of Protein Expression
2.10. IL-6 ELISA
3. Results
3.1. CARD14 Is Associated with Aggressive PCa in Human Patients
3.2. CARD14 Expression Is Essential for PCa Cell Survival
3.3. CARD14 Expression Correlates with DNA Repair Gene Enrichment
3.4. PCa Cells Show Constitutive MALT1 Proteolytic Activity, but PCa Cell Survival Only Requires MALT1 Scaffold Activity
3.5. Proteolytic Activity of MALT1 Does Not Contribute to Constitutive NF-κB/JNK/Akt Signalling in PCa Cells, but Promotes Cancer-Associated Gene Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haas, G.P.; Delongchamps, N.; Brawley, O.W.; Wang, C.Y.; de la Roza, G. The worldwide epidemiology of prostate cancer: Perspectives from autopsy studies. Can. J. Urol. 2008, 15, 3866–3871. [Google Scholar] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Di Sebastiano, K.M.; Pinthus, J.H.; Duivenvoorden, W.C.M.; Mourtzakis, M. Glucose impairments and insulin resistance in prostate cancer: The role of obesity, nutrition and exercise. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2018, 19, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Ianni, M.; Porcellini, E.; Carbone, I.; Potenzoni, M.; Pieri, A.M.; Pastizzaro, C.D.; Benecchi, L.; Licastro, F. Genetic factors regulating inflammation and DNA methylation associated with prostate cancer. Prostate Cancer Prostatic Dis. 2013, 16, 56–61. [Google Scholar] [CrossRef]
- Staal, J.; Beyaert, R. Inflammation and NF-kappaB Signaling in Prostate Cancer: Mechanisms and Clinical Implications. Cells 2018, 7, 122. [Google Scholar] [CrossRef]
- Afonina, I.S.; Van Nuffel, E.; Baudelet, G.; Driege, Y.; Kreike, M.; Staal, J.; Beyaert, R. The paracaspase MALT1 mediates CARD14-induced signaling in keratinocytes. EMBO Rep. 2016, 17, 914–927. [Google Scholar] [CrossRef]
- Howes, A.; O’Sullivan, P.A.; Breyer, F.; Ghose, A.; Cao, L.; Krappmann, D.; Bowcock, A.M.; Ley, S.C. Psoriasis mutations disrupt CARD14 autoinhibition promoting BCL10-MALT1-dependent NF-kappaB activation. Biochem. J. 2016, 473, 1759–1768. [Google Scholar] [CrossRef]
- Jordan, C.T.; Cao, L.; Roberson, E.D.; Pierson, K.C.; Yang, C.F.; Joyce, C.E.; Ryan, C.; Duan, S.; Helms, C.A.; Liu, Y.; et al. PSORS2 is due to mutations in CARD14. Am. J. Hum. Genet. 2012, 90, 784–795. [Google Scholar] [CrossRef]
- Scudiero, I.; Vito, P.; Stilo, R. The three CARMA sisters: So different, so similar: A portrait of the three CARMA proteins and their involvement in human disorders. J. Cell Physiol. 2014, 229, 990–997. [Google Scholar] [CrossRef]
- Jordan, C.T.; Cao, L.; Roberson, E.D.; Duan, S.; Helms, C.A.; Nair, R.P.; Duffin, K.C.; Stuart, P.E.; Goldgar, D.; Hayashi, G.; et al. Rare and common variants in CARD14, encoding an epidermal regulator of NF-kappaB, in psoriasis. Am. J. Hum. Genet. 2012, 90, 796–808. [Google Scholar] [CrossRef]
- Coornaert, B.; Baens, M.; Heyninck, K.; Bekaert, T.; Haegman, M.; Staal, J.; Sun, L.; Chen, Z.J.; Marynen, P.; Beyaert, R. T cell antigen receptor stimulation induces MALT1 paracaspase-mediated cleavage of the NF-kappaB inhibitor A20. Nat. Immunol. 2008, 9, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Staal, J.; Driege, Y.; Bekaert, T.; Demeyer, A.; Muyllaert, D.; Van Damme, P.; Gevaert, K.; Beyaert, R. T-cell receptor-induced JNK activation requires proteolytic inactivation of CYLD by MALT1. EMBO J. 2011, 30, 1742–1752. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.L.; Jin, G.; Li, C.F.; Jeong, Y.S.; Moten, A.; Xu, D.; Feng, Z.; Chen, W.; Cai, Z.; Darnay, B.; et al. Cycles of ubiquitination and deubiquitination critically regulate growth factor-mediated activation of Akt signaling. Sci. Signal. 2013, 6, ra3. [Google Scholar] [CrossRef]
- Seshadri, M.R.; Melnick, A.M. Targeting MALT1 for the treatment of diffuse large B-cell lymphoma. Leuk. Lymphoma 2022, 63, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Mahanivong, C.; Chen, H.M.; Yee, S.W.; Pan, Z.K.; Dong, Z.; Huang, S. Protein kinase C alpha-CARMA3 signaling axis links Ras to NF-kappa B for lysophosphatidic acid-induced urokinase plasminogen activator expression in ovarian cancer cells. Oncogene 2008, 27, 1273–1280. [Google Scholar] [CrossRef]
- Pan, D.; Jiang, C.; Ma, Z.; Blonska, M.; You, M.J.; Lin, X. MALT1 is required for EGFR-induced NF-kappaB activation and contributes to EGFR-driven lung cancer progression. Oncogene 2016, 35, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Zhu, Y.; Zhou, Z.; Wang, T.; You, H.; Jiang, C.; Lin, X. The CBM Complex Underwrites NF-kappaB Activation to Promote HER2-Associated Tumor Malignancy. Mol. Cancer Res. 2016, 14, 93–102. [Google Scholar] [CrossRef]
- Rehman, A.O.; Wang, C.Y. CXCL12/SDF-1 alpha activates NF-kappaB and promotes oral cancer invasion through the Carma3/Bcl10/Malt1 complex. Int. J. Oral Sci. 2009, 1, 105–118. [Google Scholar] [CrossRef]
- Tan, H.; Xie, Y.; Zhang, X.; Wu, S.; Zhao, H.; Wu, J.; Wang, W.; Lin, C. Integrative Analysis of MALT1 as a Potential Therapeutic Target for Prostate Cancer and its Immunological Role in Pan-Cancer. Front. Mol. Biosci. 2021, 8, 714906. [Google Scholar] [CrossRef]
- Tsui, K.H.; Chang, K.S.; Sung, H.C.; Hsu, S.Y.; Lin, Y.H.; Hou, C.P.; Yang, P.S.; Chen, C.L.; Feng, T.H.; Juang, H.H. Mucosa-Associated Lymphoid Tissue 1 Is an Oncogene Inducing Cell Proliferation, Invasion, and Tumor Growth via the Upregulation of NF-kappaB Activity in Human Prostate Carcinoma Cells. Biomedicines 2021, 9, 250. [Google Scholar] [CrossRef]
- Juilland, M.; Thome, M. Role of the CARMA1/BCL10/MALT1 complex in lymphoid malignancies. Curr. Opin. Hematol. 2016, 23, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Qu, L.; Dong, Q.; Huang, B.; Li, H.; Tang, Z.; Xu, Y.; Luo, W.; Liu, L.; Qiu, X.; et al. Overexpression of CARMA3 in non-small-cell lung cancer is linked for tumor progression. PLoS ONE 2012, 7, e36903. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; He, K.; Cao, Z.; Bi, L.; Yu, D.; Wang, Q.; Wang, J. CARD10 promotes the progression of renal cell carcinoma by regulating the NFkappaB signaling pathway. Mol. Med. Rep. 2020, 21, 329–337. [Google Scholar] [PubMed]
- Zhao, T.; Miao, Z.; Wang, Z.; Xu, Y.; Wu, J.; Liu, X.; You, Y.; Li, J. CARMA3 overexpression accelerates cell proliferation and inhibits paclitaxel-induced apoptosis through NF-kappaB regulation in breast cancer cells. Tumour Biol. 2013, 34, 3041–3047. [Google Scholar] [CrossRef]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjostedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef]
- Grasso, C.S.; Wu, Y.M.; Robinson, D.R.; Cao, X.; Dhanasekaran, S.M.; Khan, A.P.; Quist, M.J.; Jing, X.; Lonigro, R.J.; Brenner, J.C.; et al. The mutational landscape of lethal castration-resistant prostate cancer. Nature 2012, 487, 239–243. [Google Scholar] [CrossRef]
- Bardet, M.; Unterreiner, A.; Malinverni, C.; Lafossas, F.; Vedrine, C.; Boesch, D.; Kolb, Y.; Kaiser, D.; Gluck, A.; Schneider, M.A.; et al. The T-cell fingerprint of MALT1 paracaspase revealed by selective inhibition. Immunol. Cell Biol. 2018, 96, 81–99. [Google Scholar] [CrossRef]
- Firehose Broad GDAC. Available online: https://gdac.broadinstitute.org/ (accessed on 4 March 2019).
- The Cancer Genome Atlas. Available online: https://portal.gdc.cancer.gov/ (accessed on 23 March 2019).
- cBioPortal. Available online: https://www.cbioportal.org/ (accessed on 19 March 2019).
- Venet, D.; Dumont, J.E.; Detours, V. Most random gene expression signatures are significantly associated with breast cancer outcome. PLoS Comput. Biol. 2011, 7, e1002240. [Google Scholar] [CrossRef]
- PANTHER. Available online: http://www.pantherdb.org/ (accessed on 17 May 2022).
- GSEA. Available online: http://www.gsea-msigdb.org/gsea/index.jsp (accessed on 27 April 2021).
- Human DNA Repair Genes. Available online: https://www.mdanderson.org/documents/Labs/Wood-Laboratory/human-dna-repair-genes.html (accessed on 18 May 2022).
- de Vasconcelos, N.M.; Van Opdenbosch, N.; Van Gorp, H.; Martín-Pérez, R.; Zecchin, A.; Vandenabeele, P.; Lamkanfi, M. An Apoptotic Caspase Network Safeguards Cell Death Induction in Pyroptotic Macrophages. Cell Rep. 2020, 32, 107959. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef]
- Nickoloff, J.A.; Taylor, L.; Sharma, N.; Kato, T.A. Exploiting DNA repair pathways for tumor sensitization, mitigation of resistance, and normal tissue protection in radiotherapy. Cancer Drug Resist. 2021, 4, 244–263. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, T.; Suzuki, S.; Takeda, M.; Peh, J.T.; Aiba, M.; Natsuga, K.; Fujita, Y.; Takeichi, T.; Sakamoto, T.; Akiyama, M.; et al. Altered replication stress response due to CARD14 mutations promotes recombination-induced revertant mosaicism. Am. J. Hum. Genet. 2021, 108, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Kramara, J.; Osia, B.; Malkova, A. Break-Induced Replication: The Where, The Why, and The How. Trends Genet. 2018, 34, 518–531. [Google Scholar] [CrossRef] [PubMed]
- Afonina, I.S.; Elton, L.; Carpentier, I.; Beyaert, R. MALT1—A universal soldier: Multiple strategies to ensure NF-kappaB activation and target gene expression. FEBS J. 2015, 282, 3286–3297. [Google Scholar] [CrossRef]
- Xu, R.; Hu, J. The role of JNK in prostate cancer progression and therapeutic strategies. Biomed. Pharmacother. 2020, 121, 109679. [Google Scholar] [CrossRef]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-mTOR Pathway and Prostate Cancer: At the Crossroads of AR, MAPK, and WNT Signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef]
- Staal, J.; Driege, Y.; Haegman, M.; Kreike, M.; Iliaki, S.; Vanneste, D.; Lork, M.; Afonina, I.S.; Braun, H.; Beyaert, R. Defining the combinatorial space of PKC:CARD-CC signal transduction nodes. FEBS J. 2021, 288, 1630–1647. [Google Scholar] [CrossRef]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods. 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Tanaka, M.; Kobiyama, K.; Honda, T.; Uchio-Yamada, K.; Natsume-Kitatani, Y.; Mizuguchi, K.; Kabashima, K.; Ishii, K.J. Essential Role of CARD14 in Murine Experimental Psoriasis. J. Immunol. 2018, 200, 71–81. [Google Scholar] [CrossRef]
- Sun, X.; Gao, H.; Yang, Y.; He, M.; Wu, Y.; Song, Y.; Tong, Y.; Rao, Y. PROTACs: Great opportunities for academia and industry. Signal. Transduct. Target. Ther. 2019, 4, 64. [Google Scholar] [CrossRef] [PubMed]
- Demeyer, A.; Staal, J.; Beyaert, R. Targeting MALT1 Proteolytic Activity in Immunity, Inflammation and Disease: Good or Bad? Trends Mol. Med. 2016, 22, 135–150. [Google Scholar] [CrossRef] [PubMed]
- Demeyer, A.; Skordos, I.; Driege, Y.; Kreike, M.; Hochepied, T.; Baens, M.; Staal, J.; Beyaert, R. MALT1 Proteolytic Activity Suppresses Autoimmunity in a T Cell Intrinsic Manner. Front. Immunol. 2019, 10, 1898. [Google Scholar] [CrossRef] [PubMed]
- Demeyer, A.; Driege, Y.; Skordos, I.; Coudenys, J.; Lemeire, K.; Elewaut, D.; Staal, J.; Beyaert, R. Long-term MALT1 inhibition in adult mice without severe systemic autoimmunity. iScience 2020, 23, 101557. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.; Junker, U.; Tritto, E.; Sutter, E.; Rubic-Schneider, T.; Morgan, H.; Niwa, S.; Li, J.; Schlapbach, A.; Walker, D.; et al. Pharmacological Inhibition of MALT1 Protease Leads to a Progressive IPEX-Like Pathology. Front. Immunol. 2020, 11, 745. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Chalishazar, A.; Helou, Y.; DiSpirito, J.; DeChristopher, B.; Chatterjee, D.; Merselis, L.; Vincent, B.; Monroe, J.G.; Rabah, D.; et al. Pharmacological Inhibition of MALT1 Ameliorates Autoimmune Pathogenesis and Can Be Uncoupled From Effects on Regulatory T-Cells. Front. Immunol. 2022, 13, 875320. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanneste, D.; Staal, J.; Haegman, M.; Driege, Y.; Carels, M.; Van Nuffel, E.; De Bleser, P.; Saeys, Y.; Beyaert, R.; Afonina, I.S. CARD14 Signalling Ensures Cell Survival and Cancer Associated Gene Expression in Prostate Cancer Cells. Biomedicines 2022, 10, 2008. https://doi.org/10.3390/biomedicines10082008
Vanneste D, Staal J, Haegman M, Driege Y, Carels M, Van Nuffel E, De Bleser P, Saeys Y, Beyaert R, Afonina IS. CARD14 Signalling Ensures Cell Survival and Cancer Associated Gene Expression in Prostate Cancer Cells. Biomedicines. 2022; 10(8):2008. https://doi.org/10.3390/biomedicines10082008
Chicago/Turabian StyleVanneste, Domien, Jens Staal, Mira Haegman, Yasmine Driege, Marieke Carels, Elien Van Nuffel, Pieter De Bleser, Yvan Saeys, Rudi Beyaert, and Inna S. Afonina. 2022. "CARD14 Signalling Ensures Cell Survival and Cancer Associated Gene Expression in Prostate Cancer Cells" Biomedicines 10, no. 8: 2008. https://doi.org/10.3390/biomedicines10082008